
黄瓜枯萎病拮抗菌的拮抗促生因子检测及生物效应研究
2018-07-04 07:51:28董星晨罗超越邓德雷张春红王友玲邱慧珍
干旱地区农业研究 2018年3期
马 兴,董星晨,张 健,罗超越,邓德雷,张春红,王友玲,邱慧珍
(甘肃农业大学资源与环境学院/甘肃省干旱生境作物学重点实验室,甘肃 兰州 730070)
黄瓜是具有较高经济价值的重要蔬菜作物[1],中国的黄瓜种植面积和总产量一直位于世界首位。甘肃省是我国北方黄瓜的主要种植区[2],2014年种植面积达到2.33万hm2[3]。近年来,黄瓜高密度、高复指数的种植模式使其土传病害频繁发生,尤其是由尖孢镰刀菌黄瓜专化型(FOC,Fusariumoxysporumf.sp.cucumerinum)侵染引起的黄瓜枯萎病,是一种极具毁灭性的土传真菌病害[4-5],在我国北方黄瓜主要种植区该病造成黄瓜产量下降10%~30%,严重时可达80%~90%,给种植者带来极大的经济损失[6-7]。
目前黄瓜枯萎病的防治方法主要采用化学防治[8]、物理防治[9]、农业防治[10]及生物防治[11]等方法,其中以药剂为主导的化学防治是控制黄瓜枯萎病的重要手段,但是长期的化学防治会使病原菌产生抗性,导致防治效果衰退,而且化学药剂在使用过程中易在土壤中残留,会造成土壤微生物区系的破坏和多样性下降等一系列土壤健康问题,其严重阻碍了黄瓜产业的健康和可持续发展[12-14]。
相对于化学防治,生物防治是利用高效功能微生物对黄瓜枯萎病进行防治,是一种对环境无污染,对人畜无毒,对土壤中有益微生物无影响,对黄瓜无副作用,且尖孢镰刀菌不易产生抗性,使用成本较低的防治方法[15-17]。目前已有许多生防菌剂用于植物病虫害的防治,取得了可观的成效,其中研究最多的高效功能微生物类群有:荧光假单胞菌(Pseudomonasfluorescens)、芽孢杆菌属(Bacillusspp.)、木霉属(Trichodermaspp.)[18]。据研究报道枯草芽孢杆菌(Bacillussubtilis)制成的生物有机肥对棉花黄萎病的防治效果达到39%以上[19];俞鲁等[20]筛选的芽孢杆菌,其生物有机肥对香蕉枯萎病的防治效果达到74%;有学者用解淀粉芽孢杆菌Th-1防治西瓜枯萎病,其盆栽试验结果发现:添加菌株Th-1的处理与对照相比防治效果增加78.5%[21];Perze-Vicente等的研究发现:功能微生物——哈茨木霉与抗病品种的结合,使香蕉枯萎病的防治效果达到95%[22];有研究针对辣椒青枯病(Ralstoniasolanacarum)筛选拮抗菌,得到一株枯草芽孢杆菌(Bacillussubtilis),其对辣椒青枯病的防治效果很好[23]。对于生物防治,高效功能微生物是其关键因子[1]。
本研究采集了黄瓜蔬菜大棚枯萎病发病地块的健康植株根际土和根系,通过分离、筛选,得到2株对尖孢镰刀菌黄瓜专化型有稳定抑制作用的菌株QHZ-Yb1和QHZ-Yb2(以下简称Yb1和Yb2)并对2株菌进行了鉴定,探索了其潜在的拮抗促生因子,检验了其对黄瓜种子萌发的影响及对植株防病和促生的效果,旨在为黄瓜枯萎病的生物防治及植株健壮生长提供优良菌株。
1 材料与方法
1.1 试验材料
黄瓜枯萎病病原菌:尖孢镰刀菌黄瓜专化型(FOC,Fusariumoxysporumf.sp.cucumerinum)由本实验室提供。
培养基LB、PDA、尖孢镰刀菌选择性培养基的配制,采用凌宁等的方法[24]。
菌株DNA提取试剂盒购于上海生工生物工程股份有限公司。
1.2 拮抗菌的筛选
2014年9月在甘肃省白银市白银区,采集黄瓜蔬菜大棚枯萎病发病地块的健康植株根际土和根系。称取根际土和根系共5 g,其中根系用无菌剪刀剪成1 mm的小段,进行梯度稀释,取10-4、10-5、10-6的稀释液0.1 mL涂布,28 ℃恒温培养3 d后挑选形态各异的菌落,纯化后采用平板对峙法筛选拮抗菌,平板对峙法参照文献[12]。
1.3 拮抗菌的鉴定
拮抗菌传统的形态、生理生化鉴定参照《微生物学实验技术》的方法[25-26]。
菌株16S rDNA序列测定及分析:采用Ezup柱式基因组细菌DNA抽提试剂盒(Sangon Biotech, Shanghai, China),按照说明书的步骤提取细菌DNA。使用PCR对样品16S rDNA进行扩增,引物为:27F、1492R[27]。PCR反应程序:95 ℃ 5 min;94℃ 45 s,55 ℃ 45 s,72 ℃ 1 min,30次循环;72 ℃ 7 min,4 ℃条件下,将PCR产物进行纯化后交南京金斯瑞生物科技有限公司(Genscript,Nanjing,China)进行测序,测序结果在NCBI基因数据库中进行BLAST比对,同时使用软件MEGA7.0构建系统发育树。
1.4 拮抗菌拮抗促生因子检测
1.4.1 拮抗菌脂肽类粗提取物的抑菌效果测定 打取直径5 mm的黄瓜枯萎病病原菌菌饼接于PDA固体平板中央,将牛津杯放置于距中央2.5 cm处,每皿放置4个,每个牛津杯中加100 μL提取物,3次重复,25 ℃恒温培养7 d后,计算其抑菌率,拮抗菌脂肽类物质粗提取参照文献[28]。
1.4.2 拮抗菌分泌IAA的测定 将活化后的菌株分别接种至LB液体培养基中(每升LB液体培养基中含色氨酸5 mmol),28±2℃,170 r/min摇床培养2 d,取适量菌液,10 000 r·min-1离心10 min,取上清液1 mL于10 mL无菌离心管中,加入Salkawski’s显色剂显色,以不接菌的培养基为对照,30 min后对比观察颜色变化。定量测定:将活化后的菌株分别接种至LB液体培养基中(每升LB液体培养基中含色氨酸5 mmol),28±2 ℃,170 r/min摇床培养2 d,取适量菌液,10 000 r·min-1离心10 min,取上清液1 mL于10 mL无菌离心管中,加入PC显色剂[29]2 mL,30 min后,测定OD530值,以空白培养基为对照,用纯的IAA作标准曲线,计算菌株产IAA的量。
1.4.3 拮抗菌溶P能力测定 定性测定:将菌株点接于Pikovaskaia[30]固体平板上,每皿点接3次,3次重复,28±2 ℃恒温培养7 d,观察菌株是否形成溶磷圈。定量测定:制备108cell/mL的菌悬液,OD600≈ 2,分别接种各菌液1mL置100mL Pikovaskaia液体培养基中,28±2 ℃,170r/min摇床培养14d,取菌液于50mL的离心管,低温4 ℃,10000r/min,离心20min,以不接菌的Pikovaskaia液体培养基作对照,用钼锑抗比色法测定上清液中有效P含量[30]。
1.4.4 拮抗菌产嗜铁素的测定 在LB液体培养基中,接种活化后的菌株,28±2 ℃,170 r/min摇床培养12 h,离心,获取上清液,按1∶1比例取菌株上清液和CAS显色液[31]混合,显色2 h,以不接菌的培养基作对照,3次重复,若培养基颜色改变,则有嗜铁素产生。
1.4.5 拮抗菌产酪蛋白酶的测定 将活化后的菌株点接于酪蛋白固体培养基[32],每皿点接3次,3次重复,28±2 ℃恒温培养,观察菌株周围有无透明圈产生。
1.5 菌液对黄瓜种子萌发的影响
将黄瓜种子置于60 ℃热水中,缓慢搅拌10 min,再用1%次氯酸钠溶液处理1 min,最后用无菌水反复冲洗。分别接种菌株Yb1和Yb2于LB液体培养基中,28±2 ℃,170 r/min摇床恒温培养20 h,OD600≈ 2,分别吸取1 mL均匀置于垫有无菌湿润滤纸的培养皿内,每皿放置10粒大小均等的黄瓜种子,3次重复,再将培养皿置于28±2 ℃恒温培养箱36 h(期间定期加1~2 mL无菌水保持培养皿内湿度),对照加等量LB液体培养基,观察出芽长度。
1.6 拮抗菌生物效应研究
盆栽试验用土:用健康的大田土,土壤基本理化性状:有机质7.47 g·kg-1,碱解氮16.30 mg·kg-1,速效磷25.8 mg·kg-1,速效钾350.84 mg·kg-1,pH为7.84。盆栽试验共设4个处理:T1,对照(CK);T2,+Yb1菌悬液;T3,+Yb2菌悬液;T4,+(Yb1+Yb2)菌悬液。12盆/处理,3株/盆,5 kg土/盆,常规的水肥管理,所有处理均在黄瓜移栽前7 d接种3.29×104CFU/g尖孢镰刀菌黄瓜专化型孢子,移栽时各处理均匀拌入菌悬液,加菌量达到0.8×106CFU/g。
黄瓜种子用60 ℃温水和5%过氧化氢消毒后催芽育苗,在育苗盘育苗30 d后移栽于盆钵,移栽后观察记录植株生长和发病状况,在移栽60d时,测定不同处理黄瓜植株的生物量[17]并计算各处理枯萎病的发病率及防治效果,测定根际可培养微生物的数量[20]。发病率(%)=发病植株/调查总株数×100%,防治效果=(对照发病率-处理发病率)/对照发病率×100%[14]。
采用SPSS 20进行数据统计分析。
2 结果与分析
2.1 菌株的筛选结果
通过7d对峙试验得到2株对黄瓜枯萎病病原菌具有显著抑制作用的菌株Yb1和Yb2,抑菌带宽度达3.37 mm和3.42 mm,抑菌圈直径>2.7 cm,抑菌率分别达到61.9%和62.3%(图1)。
2.2 菌株的鉴定结果
2.2.1 菌株的形态特征结果 菌株Yb1和Yb2在LB平板培养基上,28±2 ℃,培养24 h,均可呈现明显菌落。Yb1菌落呈圆形、乳白色、不透明、表面褶皱、边缘为波状;Yb2菌落呈圆形、乳白色、不透明、表面有褶皱、边缘不整齐;3 d后两菌落均成褐色。菌株Yb1和Yb2革兰氏染色均呈阳性,均为中生芽孢。
2.2.2 菌株生理生化特性的测定结果 对菌株Yb1和Yb2进行生理生化特性的测定,结果见表1。通过自动检索数据库比对测定结果,结果表明Yb1和Yb2均属于芽孢杆菌属。
2.2.3 菌株16S rDNA序列分析结果 菌株Yb1和Yb2的16SrDNA序列长度分别为1412 bp和1417 bp。在NCBI基因数据库中进行BLAST比对,绘制系统发育树(图3)。结果表明:菌株Yb1与萎缩芽孢杆菌BacillusatrophaeusstrainJCM9070NR—024689.1的亲缘性最高(图3A),菌株Yb2与萎缩芽孢杆菌BacillusatrophaeusstrainNBRC15539NR112723.1的亲缘性最高(图3B),结合菌株的形态特征及生理生化测定结果,初步确定Yb1和Yb2均为萎缩芽孢杆菌(B.atrophaeus)。
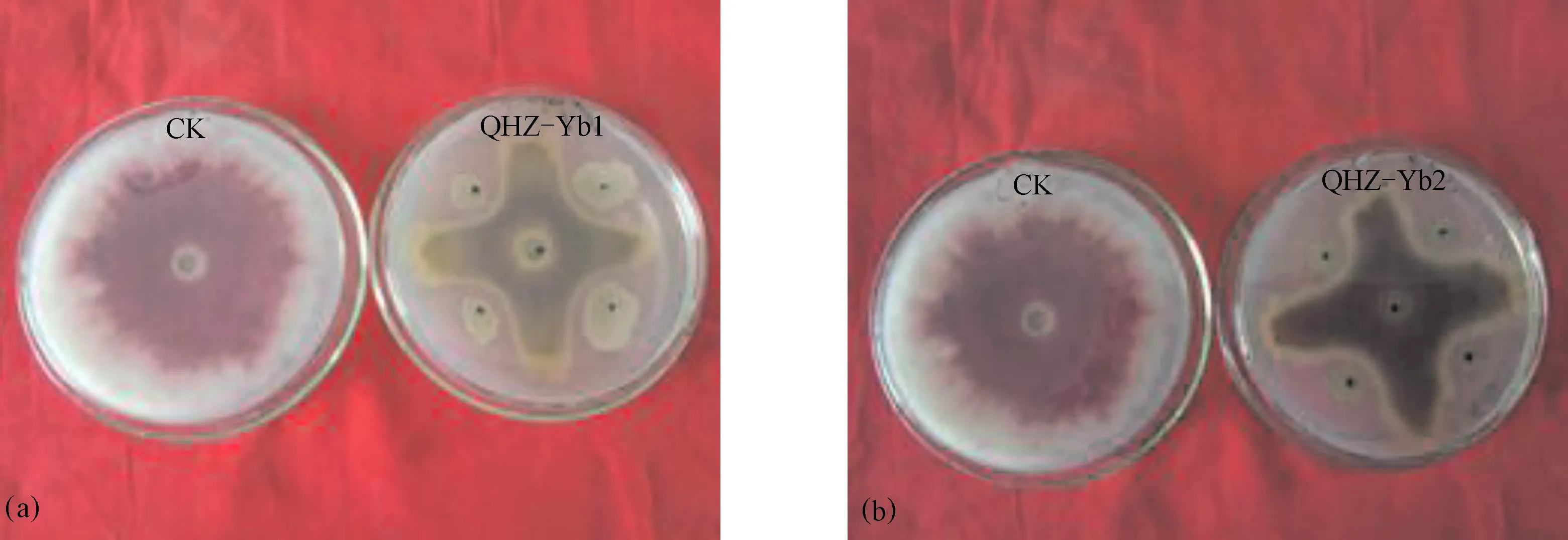
图1 菌株Yb1(a)和Yb2(b)对黄瓜枯萎病病原菌生长的影响(7 d)Fig.1 Effects of Yb1 (a) and Yb2 (b) on the growth of Fusarium oxysporum f.sp. cucumerinum(7 d)
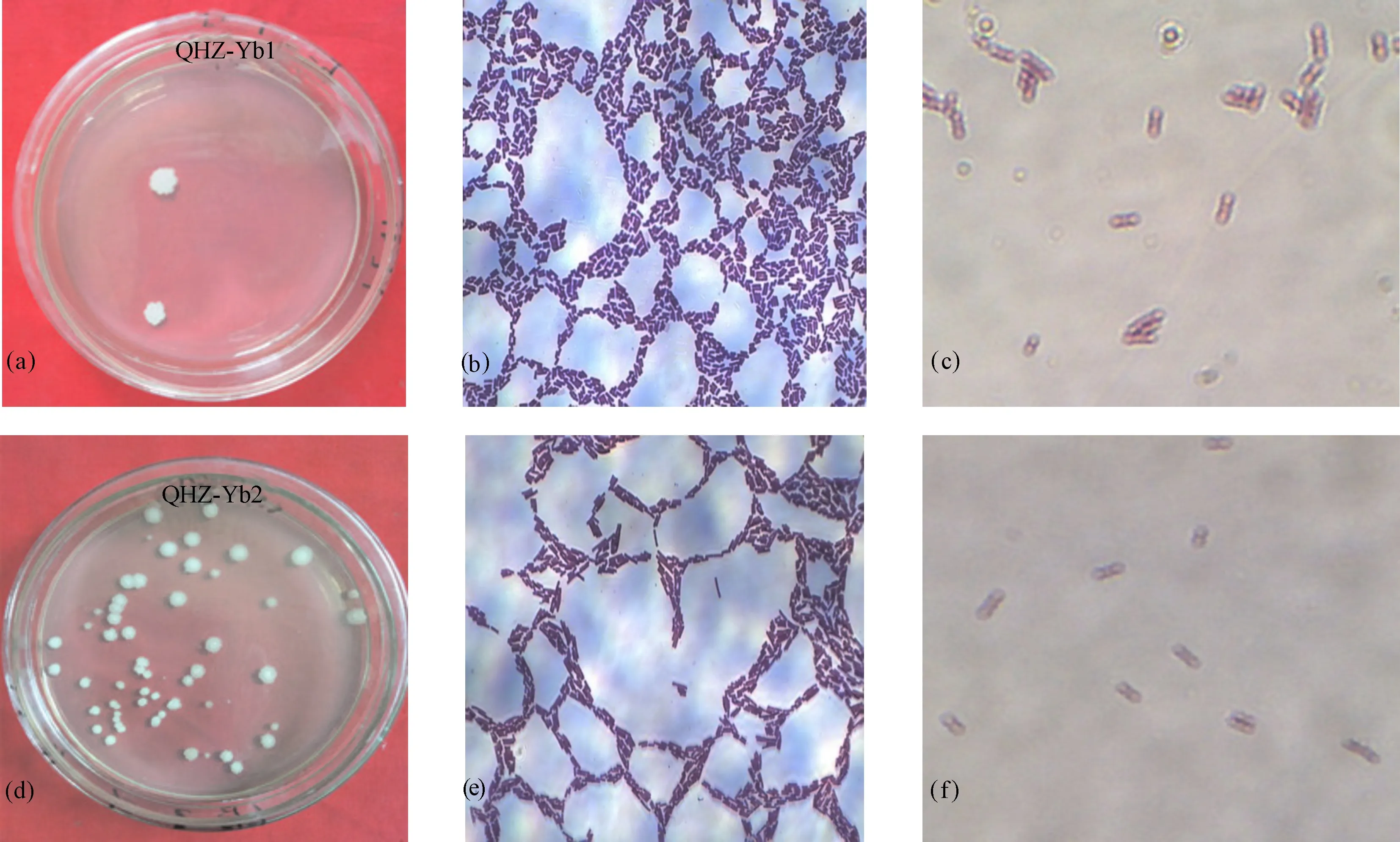
(a)Yb1菌落形态;(b)Yb1革兰氏染色;(c)Yb1芽孢染色;(d)Yb2菌落形态;(e)Yb2革兰氏染色;(f)Yb2芽孢染色(a)Colonial morphology of strainYb1; (b)Gram stain of strain Yb1; (c)Spore staining of strainYb1; (d)Colonial morphology of strain Yb2; (e)Gram stain of strain Yb2; (f)Spore staining of strain Yb2图2 菌株Yb1和Yb2形态学鉴定Fig.2 Morphological identifications of Yb1 and Yb2

测定指标Test index试验结果 ResultYb1Yb2测定指标Test index试验结果 ResultYb1Yb2葡萄糖 Glucose++产H2S Sulfuretted hydrogen++果糖 Fructose++产吲哚乙酸 Indole++蔗糖 Sucrose++尿素水解 Urea hydrolysis+-淀粉 Starch++柠檬酸 Citric acid++甘油 Glycerol++苯丙氨酸 Phenylalanine+-V-P试验 V-P test++色氨酸Tryptophane++M-R试验 M-R test--酪氨酸 Tyrosine++淀粉水解 Amylolysis++接触酶 Catalase++明胶液化 Gelatine liquefication++硝酸盐还原 Nitrate reduction++产氨 Production of ammonia++石蕊牛奶试验 Litmus milk test++
注:+:阳性反应;-:阴性反应。 Note:+:positive reaction;-:negative reaction.

图3 Yb1(A)和Yb2(B)系统发育树Fig.3 The phylogenetic tree of Yb1 and Yb2
2.3 菌株拮抗促生因子检测结果
2.3.1 菌株脂肽类粗提取物的抑菌效果 菌株Yb1和Yb2脂肽类粗提物的抑菌效果如图4,抑菌率分别为57.4%和58.0%。
2.3.2 菌株分泌IAA的测定结果 如图5结果显示,加入Salkawski’s显色剂,显色30 min后,菌株Yb1和Yb2的上清液均较CK颜色变深,说明菌株Yb1和Yb2均分泌IAA。定量测定结果表明,菌株Yb1和Yb2分泌IAA的量分别为1.56 mg·L-1,3.69 mg·L-1。
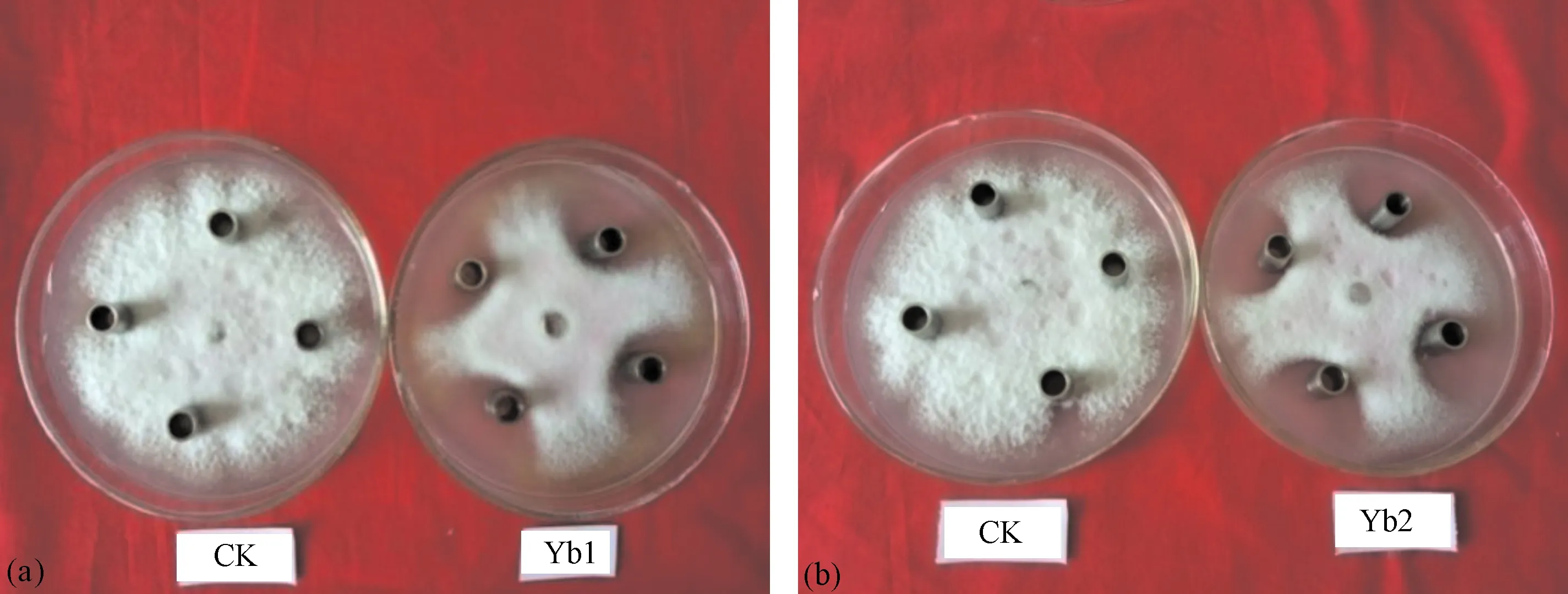
图4 Yb1(a)和Yb2(b)脂肽类粗提物对黄瓜枯萎病病原菌生长的影响Fig.4 Effects of lipopeptide from Yb1(a) and Yb2(b)on the growth of pathogen of cucumber wilt

图5 菌株Yb1(a)和Yb2(b)产IAAFig.5 IAA produced from Yb1(a) and Yb2(b)
2.3.3 菌株溶P能力测定结果 图6结果显示,在菌株Yb1和Yb2的菌落周围有明显的溶磷圈,且后者显著大于前者,定量测定结果表明,菌株Yb1和Yb2的溶磷量分别为0.77 mg·L-1和1.19 mg·L-1。
2.3.4 菌株产嗜铁素的测定结果 如图7结果显示,对比CK,菌株Yb1和Yb2的上清液颜色改变,说明菌株Yb1和Yb2均产嗜铁素。
2.3.5 菌株产蛋白酶的测定结果 点接在酪蛋白培养基(图8)上的Yb1和Yb2菌株周围均有明显的透明圈产生,说明两菌株均有产酪蛋白酶的能力且Yb2菌株产酪蛋白酶的能力强于Yb1。
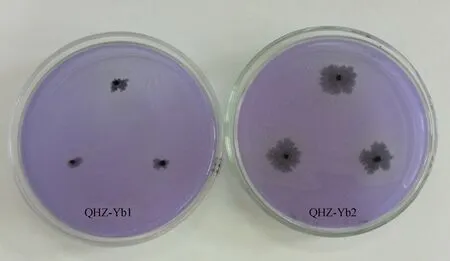
图6 菌株Yb1和Yb2溶PFig.6 Phosphorus solubilization from Yb1 and Yb2

图7 菌株Yb1(a)和Yb2(b)产嗜铁素Fig.7 Siderophore produced from Yb1(a) and Yb2(b)
2.4 菌液对黄瓜种子萌发的影响结果
如图9所示,黄瓜种子催芽36 h后,接种Yb1和Yb2菌液的黄瓜种子出芽长度明显长于CK,说明菌株Yb1和Yb2均能促进黄瓜种子的萌发及芽的生长。
2.5 拮抗菌生物效应研究结果
如表2所示,对比CK,Yb1、Yb2、Yb1+Yb2菌悬液的处理不仅对黄瓜枯萎病有不同程度的生防效果,而且均使植株生物量增加。对比CK,Yb1和Yb2菌悬液无论是单独使用还是加和在一起使用,都使黄瓜枯萎病的发病率降低5.0%以上,防治效果达到6.0%以上,其中Yb1+Yb2菌悬液的处理使黄瓜枯萎病的发病率降低了11.11个百分点。黄瓜移栽后60 d,接种Yb1、Yb2和Yb1+Yb2菌悬液处理的株高分别比CK增加了52.1%、34.0%和73.7%,植株干重增加了17.9%、8.9%和56.7%,植株地上部和根系干重分别增加了15.97%和6.14%、53.07%和61.11%、77.78%和138.89%。可以看出,Yb1和Yb2无论对黄瓜植株地上部还是根系生长的促进作用,均具有明显的加和效果。
各处理对黄瓜根际土中可培养微生物数量的计数结果如表3所示。可以看出,与CK相比,Yb1、Yb2和Yb1+Yb2菌悬液处理的黄瓜植株根际土中的细菌、放线菌数量明显增加,真菌和尖孢镰刀菌数显著减少,说明拮抗菌Yb1和Yb2能够抑制病原菌的数量,使根际微生物区系向健康转变,有利于降低植株发病率而且加和效果突出。
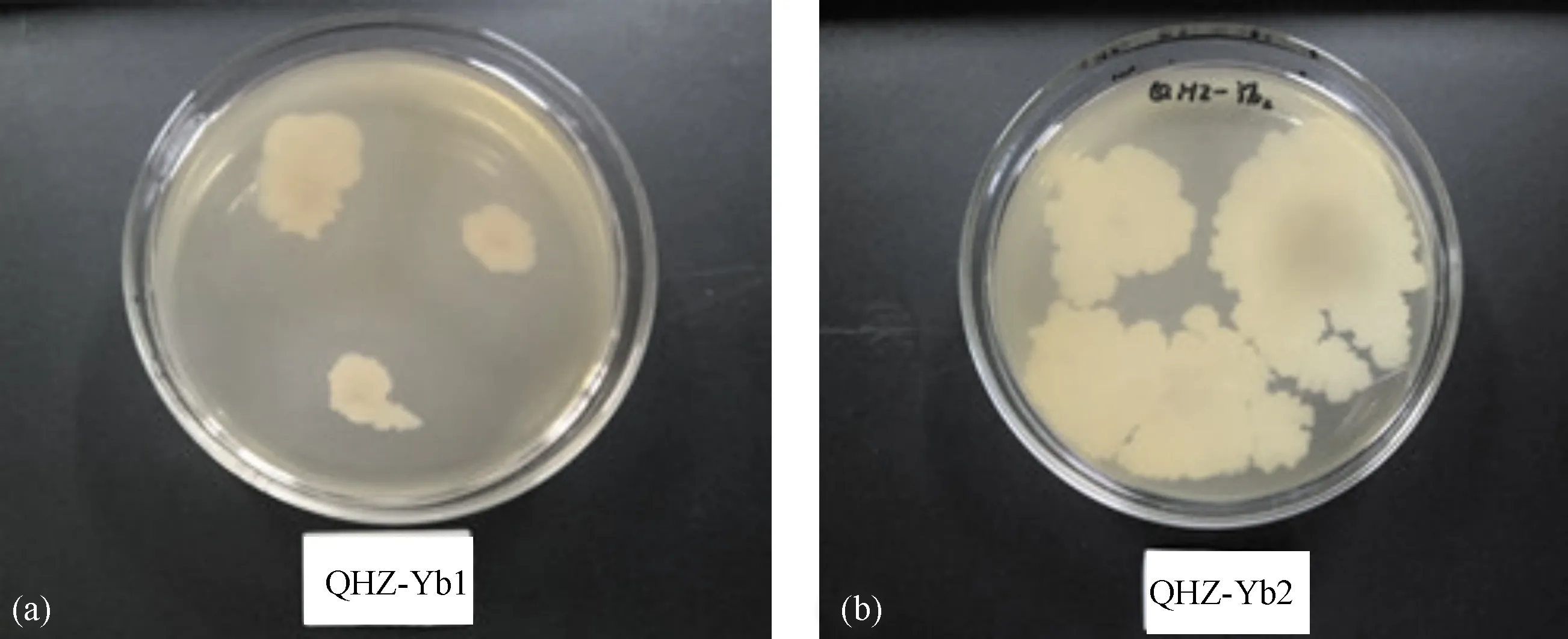
图8 菌株Yb1(a)和Yb2(b)产蛋白酶Fig.8 Casease produced from Yb1(a) and Yb2(b)
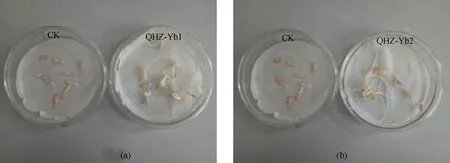
图9 菌株Yb1(a)和Yb2(b)对黄瓜种子萌发的影响Fig.9 Effects of the strains of Yb1(a) and Yb2 (b) on cucumber seed germination
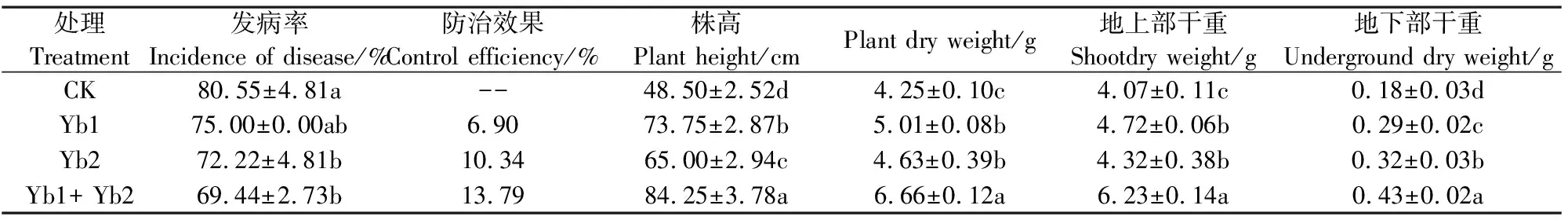
处理Treatment发病率Incidence of disease/%防治效果Control efficiency/%株高Plant height/cmPlant dry weight/g地上部干重Shootdry weight/g地下部干重Underground dry weight/gCK80.55±4.81a--48.50±2.52d4.25±0.10c4.07±0.11c0.18±0.03dYb175.00±0.00ab6.9073.75±2.87b5.01±0.08b4.72±0.06b0.29±0.02cYb272.22±4.81b10.3465.00±2.94c4.63±0.39b4.32±0.38b0.32±0.03bYb1+ Yb269.44±2.73b13.7984.25±3.78a6.66±0.12a6.23±0.14a0.43±0.02a
注:同列数值后不同字母表示差异达5%显著水平。下同。
Note:Values followed by different small letters in the same column mean significant at 5% level. The same as below.

表3 不同处理对黄瓜根际土中可培养微生物数量的影响
3 结论与讨论
芽孢杆菌是一类重要的生防菌,不仅对植物病害有防治作用而且能够促进植物的生长发育[33]。有研究发现枯草芽孢杆菌B29,其菌剂(7×106CFU·mL-1)对黄瓜枯萎病的田间防效达84.9%,增产12.57%[34]。据研究报道生防菌CT205不仅对黄瓜生长有促进作用而且对黄瓜枯萎病的防治效果达到51.85%[35]。有研究者对几株芽孢杆菌的促生能力进行了研究,结果发现其对温室条件下的玉米和番茄有显著的促生作用[36]。本研究通过平板对峙试验筛选到2株拮抗效果好的菌株:Yb1和Yb2,通过形态学结合生理生化及菌株16S rDNA序列分析,初步确定这两株菌均为萎缩芽孢杆菌(Bacillusatrophaeus)。
生防芽孢杆菌通过产生抗菌物质及其它生防机制共同或单独作用发挥良好的生防效果[37]。生防芽孢杆菌发挥抗菌作用抑制病原菌主要源于其分泌的脂肽类抗生素,包括伊枯草素(Iturin)、泛革素(Fengycin)和表面活性素(Surfactin)三大类脂肽[38]。陈莉的研究发现萎缩芽孢杆菌(Bacillusatrophaeus)B44产生的iturin A是关键的抑菌活性物质,菌株B44在田间条件下,对棉苗立枯病、棉花黄萎病、加工番茄根腐病均有良好的生防治效果[39]。生防菌Bs-916分泌的脂肽类化合物BacillomycinL,用500 μg·mL-1防治水稻纹枯病和苗瘟,防效分别为70.8%和56.3%[40]。生防芽孢杆菌也可以通过分泌细胞壁降解酶抑制病原菌的生长[41]以及通过产嗜铁素,在与病原菌竞争铁元素时占据绝对优势,使病原菌因缺铁而生长异常,进而起到生防作用[38]。研究发现菌株Yb1和Yb2的脂肽类粗提取物(用量:100 μL)对黄瓜枯萎病病原菌的抑菌率分别达到57.4%和58.0%,且均产蛋白酶和嗜铁素,菌株Yb2产蛋白酶的能力较Yb1强,说明菌株Yb1和Yb2具有良好的生防潜力,Yb2的防治效果或许强于Yb1;盆栽试验检验菌株Yb1和Yb2的生防能力发现:Yb1、Yb2及Yb1+Yb2菌悬液处理对黄瓜枯萎病的防治效果分别为6.9%、10.34%、13.79%,菌株Yb2的生防效果较Yb1好,这与菌株平板对峙试验和菌株脂肽类粗提取物的抑菌效果及酪蛋白酶的分泌量结果一致;Yb1+Yb2菌悬液处理的生防效果最好,说明两菌株具有加和效应。
有些生防芽孢杆菌通过促进植物生长间接的发挥其生防作用[42],其促生机制包括:产生长素(IAA)促进植物生长、产嗜铁素将多余的铁元素供给植物、溶磷使土壤中无效态磷转化成有效态磷[43]等。菌株Yb1和Yb2既能分泌生长素(IAA),也可溶磷并产嗜铁素,通过种子发芽试验及盆栽试验检验其防病和促生能力发现:菌株Yb1和Yb2对黄瓜种子的萌发有促进作用。盆栽试验结果表明:对比CK,Yb1、Yb2及Yb1+Yb2菌悬液处理均使黄瓜植株生物量增加,株高分别增加了52.1%、34.0%、73.7%,植株干重增加了17.9%,8.9%和56.7%,植株地上部和根系干重分别增加了15.97%、6.14%、53.07%,61.11%、77.78%、138.89%。以上说明,菌株Yb1和Yb2对黄瓜植株地上部及根系生长具有促进作用且有明显的加和效应;株高和植株地上部干重的增加量Yb1>Yb2,根系干重的增加量Yb2>Yb1,这可能与菌株促生机制的主要作用部位和产生量有关。
根际是病原微生物和有益微生物的运动场和战场[44],根际病原微生物和有益微生物的数量和密度对植物生长和健康有非常重要的影响[45-46]。健康的根际体系,微生物群体相对平衡,这种平衡一旦被打破,有益微生物减少,病原微生物增加,从而导致植物病害产生及作物产量下降[47]。在土壤中添加有益微生物,不仅改善根际微生物群体的相对平衡,而且有利于防治植物病害。盆栽试验的结果表明:与CK相比,Yb1、Yb2及Yb1+Yb菌悬液处理尖孢镰刀菌及真菌数量明显减少,细菌和放线菌的数量增加,根际微生物区系明显改善,结果与其防治效果一致,两种菌的加和处理对尖孢镰刀菌的抑制效果更好。
菌株Yb1和Yb2不仅具有多种拮抗促生因子,而且在实践中表现了抗病促生能力,因此菌株Yb1和Yb2具有生防潜力,两株菌配合应用效果更好。
参考文献:
[1] 韦巧婕.黄瓜土传枯萎病拮抗菌的筛选鉴定及其生物学效应研究[D].南京:南京农业大学,2012.
[2] 赵瑛,罗俊杰,王方,等. 微生物菌肥在保护地黄瓜栽培上的应用研究进展[J].甘肃农业科技,2015,(06):62-65.
[3] 中华人民共和国农业部种植业管理司.中国种植业信息网-蔬菜数据库[EB/OL].http://zzys.agri.go.v.cn, 2014.
[4] Shen W, Lin X, Zhang H,et al. Land use intensification affects soil microbial populations, funcional diversity and related suppressiveness of cucumber Fusarium wilt in China’s Yangtze River Delta [J]. Plant and Soil, 2008,306(2):117-127.
[5] 薛超,黄启为,凌宁,等.连作土壤微生物区系分析、调控及高通量研究方法[J].土壤学报,2011,48(03):612-618.
[6] Pu Z J, Zhang Y J, Liu D, et al. Research progress inbiological control strategies for Fusarium wilt of cucumber(in Chinese) [J]. China Vegetables (中国蔬菜),2011,(6):9-14.
[7] 郭晋云,胡晓峰,李勇,等. 黄瓜枯萎病对黄瓜光合和水分生理特性的影响[J].南京农业大学学报,2011,34(01):79-83.
[8] 王惟萍,石延霞,赵一杰,等.土壤环境条件对威百亩熏蒸防治黄瓜枯萎病的影响[J].植物保护学报,2017,(1):159-167.
[9] 李新文,袁清. 黄瓜枯萎病及其综合防治技术[J].现代园艺,2009,(3):30-31.
[10] 赵帅,杜春梅,田长彦.黄瓜枯萎病综合防治研究进展[J].中国农学通报,2014,34(7):254-259.
[11] 闫霜,吴洪生,周晓冬,等. 黄瓜枯萎病生物防治研究进展[J].山东农业科学, 2011(1): 86-92.
[12] 乔胜伟. 一株黄瓜枯萎病拮抗细菌的分离鉴定及其相关特性的研究[D].南京:南京农业大学,2010.
[13] 王敏. 土传黄瓜枯萎病致病生理机制及其与氮素营养关系研究[D].南京:南京农业大学,2013.
[14] Uppal A K, Hadrami A E I, Adam L R, et al. Pathogenic variability of Verticillium dahlia isolates from potato fields in Manitoba and screening of bacteria for their biocontrol [J]. Can J Plant Pathol,2007 29(2):141-152.
[15] Adhikari T B, Joseph C M, Yang G, et al. Luation of bacteria isolated from rice for plant growth promotion and biological control of ceedling disease of rice [J].CanadianJournal of Microbiology,2001, 47: 916-924.
[16] Eilenberg J, Hokkanen H M T. An Ecological and Societal Approach to Biological Control[M]. Springe,2006.
[17] 王玲平.黄瓜感染枯萎病菌后生理生化变化及其与抗病性关系的研究[D].太谷:山西农业大学,2011.
[18] 袁玉娟. Bacillus subtilis SQR9的黄瓜促生和枯萎病生防效果及其作用机制研究[D].南京:南京农业大学,2011.
[19] 程凯,江欢欢,沈标,等. 棉花黄萎病拮抗菌的筛选及其生物防治效果[J].植物营养与肥料学报,2011,17 (1):166-174.
[20] 俞鲁,凌宁,张楠,等. 香蕉枯萎病拮抗菌的筛选鉴定及其生防效果[J].南京农业大学学报,2012,35(4):81-86.
[21] 赵佳,黄静,陈哲.西瓜枯萎病拮抗菌Lh-1的鉴定及生物防治效果研究[J].生物技术通报,2017,339(4):1- 7.
[22] Perze-Vicente L, Batlle A, Fonseca J,et al.Fusariumoxysprumf.sp. Cuba: reaction of cultivars and biocontrol method[C]//Picq C, Vezina A. 2nd International Symposium on Fusarium Wilt on Banana. Salvador de Bahia, 2003: 22-26.
[23] 江欢欢,沈标,杨兴明,等. 辣椒青枯病拮抗菌的筛选及其生物防治效应[J].土壤学报,2010(6):1225-1231.
[24] 凌宁,王秋君,杨兴明,等. 根际施用微生物有机肥防治连作西瓜枯萎病研究[J].植物营养与肥料学报,2009, 15(5):1136-1141.
[25] 程丽娟,薛泉宏.微生物学实验技术(第二版)[M].北京:科学出版社,2012.
[26] 东秀珠,蔡妙英.常见细菌系统鉴定手册[M].北京:科学出版社,2001.
[27] Eden P Z, Schmidt T M, Blakemore R P, et al. Phylogenetic analysis of Strain Yb1(a) and Yb2(b) of using polymerase chain reaction-amplified 16S Rrna-SPECIFIC DNA [J]. International Journal of Systeamtic Bacteriology, 1991, 41:324-325.
[28] Hsieh F C, Lin T C, Meng M, et al. Comparing methods for identifying Bacillus strains capable of producing the antifungal lipopeptide iturin A [J]. Current Microbiology, 2008, 56:1-5.
[29] 高晓星,满百膺,陈秀荣,等.东祁连山线叶嵩草内生细菌X4的产吲哚乙酸、解磷、抗菌和耐盐特性研究及分子鉴定[J].草业学报,2013,22(4):137-146.
[30] 姚拓.高寒地区燕麦根际联合固氮菌研究Ⅱ固氮菌的溶磷性和分泌植物生长素特性测定[J].草业学报,2004,13(3):83-90.
[31] Smith K P, Goodman R M. Host variation for interactions with beneficial plant associated microbes [J].Annal Review of Phytopathology, 1999, 96:4786-4790.
[32] 丁传雨.土传马铃薯青枯病的生物防控及机理研究[D].南京:南京农业大学,2012.
[33] 杨晓云,陈志谊,蒋盼盼,等. 解淀粉芽孢杆菌B1619对番茄的促生作用[J].中国生物防治学报,2016,32(3):349356.
[34] 李晶.黄瓜枯萎病高效拮抗枯草芽孢杆菌的筛选及生防机制研究[D].哈尔滨:哈尔滨工业大学,2010.
[35] 梁银,张谷月,王辰,等. 一株拮抗放线菌的鉴定及其对黄瓜枯萎病的生防效应研究[J].土壤学报,2013,50 (4):810- 817.
[36] Huang X F, Zhou D, Guo J, et al.Bacillusspp. from rainforest soil promote plant growth under limited nitrogen conditions[J]. Applied Microbiology, 2015, 118:672- 684.
[37] 孙冰冰,李伟,魏军,等. 生防芽孢杆菌的研究进展[J].天津农业科学,2015,(12):102-107.
[38] 武志江,李业燕,王亚军,等.百合枯萎病拮抗细菌的筛选、鉴定及其抑菌物质研究[J].微生物学通报,2015,42(7):1307-1320.
[39] 陈莉. Bacillus atrophaeus B44产生iturin A及其对棉苗立枯病的防病促生作用[D].石河子:石河子大学,2015.
[40] 罗楚平,刘邮洲,吴荷芳,等. 脂肽类化合物bacillomycin L抗真菌活性及其对水稻病害的防治[J].中国生物防治学报,2011,27(1):76-81.
[41] Saran S, Isar I, Saxena R. Amodified method for the detection of microbial proteases on agar plate using tannic acid [J].Journal of Biochemical and Biophysical Methods, 2007, 70:697-699.
[42] 彭研,陈相艳,裘纪莹,等. 生防芽孢杆菌的研究进展[J].山东农业科学,2013,( 7):138-140.
[43] 康贻军,程洁,梅丽娟,等. 植物根际促生菌作用机制研究进展[J].应用生态学报,2010,21(1):232-238.
[44] Jos M. Raaijmakers, Timothy C. Pauilitz, Christian Steinberg, et al. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms [J].Plant Soil, 2009, 321: 341-361.
[45] Lynch J M. Resilience of the rhizosphere to anthropogenic disturbance [J]. Biodegradation, 2002, 13(1):21-27.
[46] Avis T J, Gravel V. Antoun H. Tweddell R J. Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity [J]. Soil Biol Biochem, 2008, 40(7):1733-1740.
[47] 张慧,杨兴明,冉炜,等. 土传棉花黄萎病拮抗菌的筛选及其生物效应[J].土壤学报,2008, 45(6):1095-1101.
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
现代园艺(2017年23期)2018-01-18 06:57:48
中国蔬菜(2016年8期)2017-01-15 14:23:38
中国酿造(2016年12期)2016-03-01 03:08:15
中国果业信息(2016年2期)2016-01-29 03:17:16
湖南农业科学(2014年1期)2014-02-27 14:25:50
中国果业信息(2012年9期)2012-01-23 07:30:01
植物营养与肥料学报(2011年3期)2011-10-24 06:14:54
植物营养与肥料学报(2010年4期)2010-11-06 07:29:18
