
谷子新品种(系)主要农艺性状及茎杆特性的遗传多样性分析
2018-07-04 08:01王可珍高小丽高金锋王鹏科冯佰利
干旱地区农业研究 2018年3期
屈 洋,宋 慧,刘 洋,王可珍,高小丽,高金锋,王鹏科,杨 璞,冯佰利
(1. 旱区作物逆境生物学国家重点实验室/西北农林科技大学,陕西 杨凌 712100;2. 宝鸡市农业科学研究院,陕西 岐山 722499;3.安阳市农业科学院,河南 安阳 455000)
作物表型性状是由遗传物质和环境综合作用的结果[1],是环境影响、自然选择和人工干预产生的稳定遗传变异[2]。作物表型性状的多样性对环境适应性和作物进化具有重要作用。谷子(SetariaitalicBeauv.)耐旱、耐瘠薄,主要分布在北方农牧交错地区,是我国干旱半干旱地区的主要作物[3]。了解谷子新品种(系)表型性状的多样性,研究农艺及茎杆特性与产量性状的关系,对提高谷子新品种(系)资源有效利用和谷子育种水平具有重要意义。表型性状的描述和多样性分析是作物种质资源有效利用的重要方法和途径[4],表型性状的多样性分析已应用于水稻、小麦、玉米、棉花、大豆种质资源或品种的分析[5-9]。此外,甜高粱[10],燕麦[11],荞麦[12],糜子[13]等也进行了表型性状的多样性分析,以提高作物种质资源的利用效率。近年来,谷子表型性状的相关性分析主要集中在产量性状上[14-15],例如出谷率、穗粒重、单穗重、千粒重、穗长、穗粗等。杨成元[16]利用灰色关联度法分析了农艺性状对产量的影响;黄英杰等[17]对谷子主要产量构成因素进行了稳定性分析;赵禹凯等[18]对谷子主要农艺性状进行了相关和通径分析;王海岗[19]对谷子核心种质进行了表型性状的多样性分析并对资源进行了综合评价。目前,谷子表型性状的多样性分析主要集中在产量性状上[20-21],茎杆特性方面的研究较少。本研究以31份谷子新品种(系)为材料,利用表型性状和茎杆性状的鉴定方法对农艺和茎杆性状的遗传多样性进行分析,揭示农艺和茎杆特性与产量性状之间的关系,为谷子品种资源的利用提供参考。
1 材料与方法
1.1 试验地点
试验于2016年在宝鸡市农科院刘家塬试验站(107.5°E, 34.5°N)进行。试验地所在地区为暖温带大陆性季风型半湿润气候,年平均气温12℃,极端最高温度36℃左右,极端最低温度-4℃左右,年平均日照时数2064.8 h,平均无霜期214 d,年平均降水量623.8 mm。
1.2 参试材料与试验设计
以31份谷子品种(系)为参试材料(表1),采用完全随机区组设计,3次重复,小区面积3.5m×4.6m=16 m2。4月29日播种,密度33万株·hm-2,行距50 cm,株距6 cm。播前施用磷酸二铵600 kg·hm-2。于拔节期盖防鸟网,防鸟食。各品种(系)进入蜡熟期开始取样收获,9月10日完成收获。
1.3 数据采集
调查每个小区的出苗期、拔节期、抽穗期、成熟期、株高、茎粗、节长、苗色、苗叶资、株型、米色、穗型等性状,参照谷子种质资源描述规范和数据标准进行[22],并对每个品种(系)的节长和茎粗进行全节位测量。每个小区内随机取样10株测量谷子品种(系)各性状值,各性状取平均值待分析。
1.4 数据处理
分级性状主要分析性状的频率分布和多样性指数,具体分级标准见表2。利用Excel 2007和SPSS 17.0进行描述性和频次分布统计,完成对农艺性状、茎杆特性的最大值、最小值、平均值、标准差、变异系数和多样性指数的计算。利用Shannon-Weanver遗传多样性指数衡量群体遗传多样性大小,计算公式为:H′=-∑Pi×lnPi[23]。其中H′为遗传多样性指数,Pi为某一性状第i级别内材料份数占总份数的百分比,ln为自然对数。利用SPSS 17.0进行主成分分析和差异性检验(Tukey法,P<0.05),并利用DPS7.05计算Euclidean(欧式)距离,并用Ward(离差平方和)法进行聚类。
2 结果与分析
2.1 谷子农艺性状的遗传多样性分析
2.1.1 分级性状的遗传多样性 8个分级性状间的多样性指数见表3。各分级性状间多样性指数差异较大,株型、穗型和穗密度频率在1、3、5、7级均有分布,离散性较高,且多样性指数分别为1.259,1.022和1.237;苗色、苗叶资和花药色频率在1、4、7级有分布,离散程度适中,多样性指数分别为0.512、0.689和0.579;谷粒颜色、米色频率在1和7级有分布,离散性较差,多样性指数最低分别为0.241和0.142。
2.1.2 谷子产量性状的遗传多样性 产量相关性状遗传多样性指数较高,变异系数范围为3.460~24.852,多样性指数范围为1.539~2.170。单穗重、穗粒重、株高的变异系数分别为23.794%、24.852%和11.855%,且遗传多样性指数分别为2.170、1.165和2.121,多样性丰富;穗粗、千粒重的变异系数为10.980%和20.862%,且遗传多样性指数为2.086和2.025,多样性较丰富;生育期、出谷率变异系数为5.135%和3.460%,且多样性指数为1.648和1.539,多样性较差(表4)。

表1 谷子新品种(系)名称及来源

表2 谷子8个分级性状及调查标准

表3 谷子8个分级性状的频率分布和多样性指数
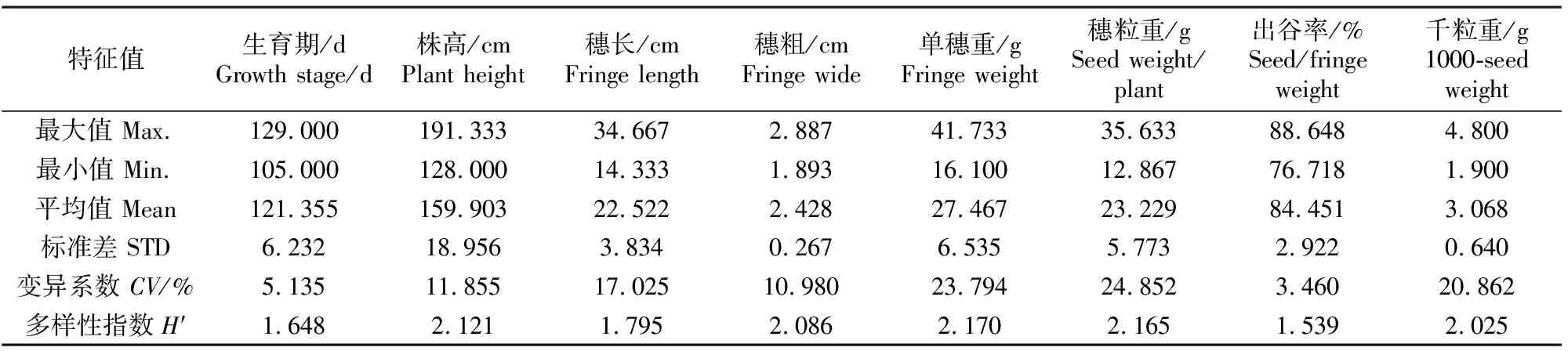
表4 谷子农艺性状的遗传多样性
2.2 谷子茎杆特性的遗传多样性
2.2.1 谷子基部和首部3节特性的遗传多样性 选取谷子品种(系)的基部和首部的3节,计算节长和节粗的遗传多样性指数(表5)。节长中,变异系数变化范围为23.294%~26.811%,多样性指数变化范围为1.860%~2.184%,首部第3节变异系数较大,多样性指数最高;基部第2节变异系数较低,多样性指数较低,多样性差。节粗中,变异系数范围为16.455%~25.610%,多样性指数变化范围为1.473%~2.023%,首部第1节变异系数中等,多样性指数较大,多样性丰富;基部第2节变异系数较高,多样性指数较低,多样性差。
2.2.2 谷子基部和首部3节主成分分析 对谷子茎杆特性的6个表型性状进行主成分(PV)分析,前3个主成分( PV(1)、PV(2)和PV(3))累计贡献率达99.577%(表6)。PV(1)主要包括首部节长第1、2、3节,首部和基部的节粗第1、2、3节;PV(2)主要包括首部第2节节长;PV(3)主要包括基部第3节节长。进一步以PC1为横坐标,PC2为纵坐标,绘制散点图。不同节位节粗具有一定的群集性,不同节位节长位点分布较为分散,但是首部和基部的第2、3节长具有一定的群集性(图1)。
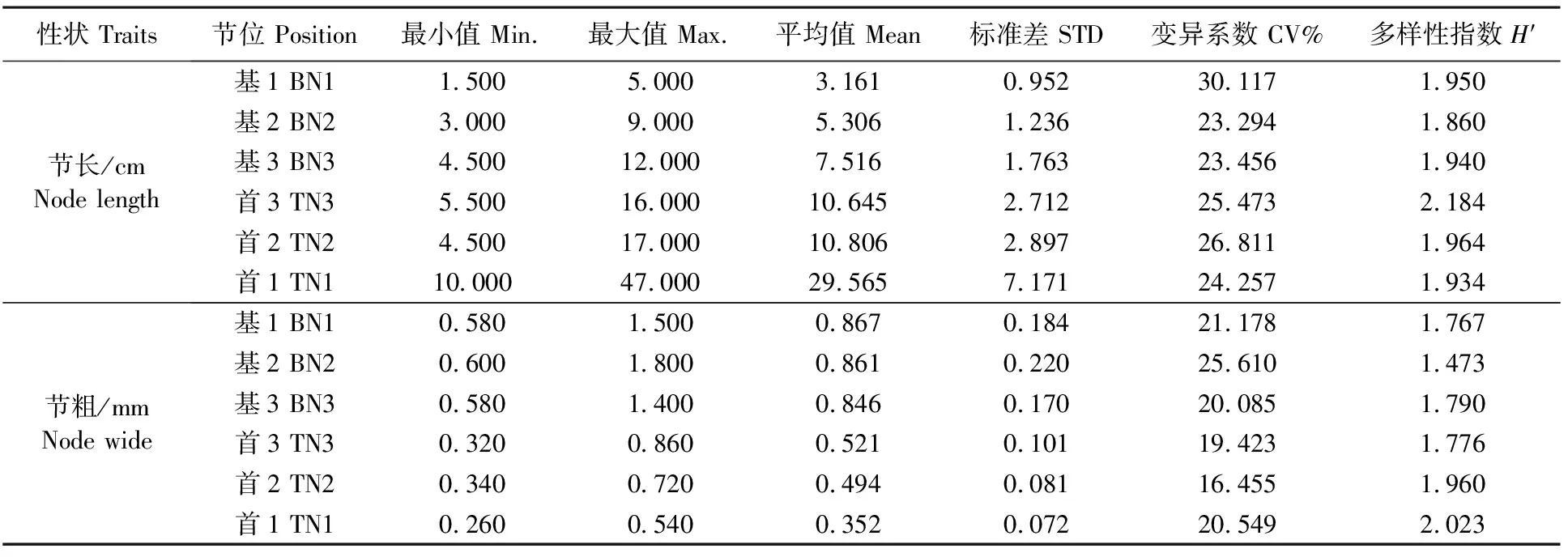
表5 谷子茎杆特性的遗传多样性
注:基1、2和3:为基部第一节、第二节和第三节;首1、2和3:为首部第一节、第二节和第三节。
Note:BN(1,2,3):The first(second,third) node from bottom;TN(1,2,3):The first(second,third) node from top.
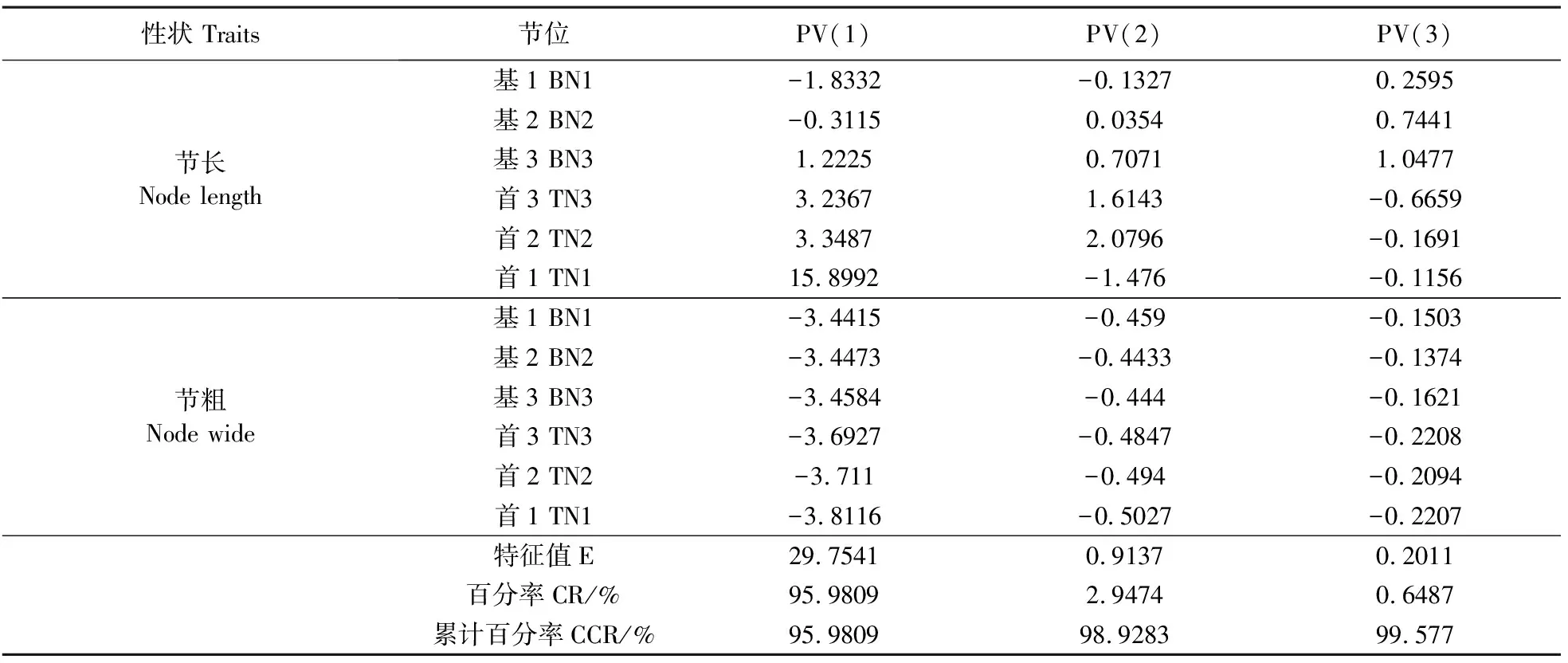
表6 基部和首部茎杆特性的前3个主成分的特征向量、主成分值,贡献率和累积贡献率
注:基1、2和3:为基部第一节、第二节和第三节;首1、2和3:为首部第一节、第二节和第三节。
Note:BN(1,2,3):the first(second,third) node from bottom;TN(1,2,3):the first(second,third) node from top.

注:BNL(1、2和3):基部第一节、第二节和第三节长;TNL(1、2和3):首部第一节、第二节和第三节长;BNW(1、2和3):基部第一节、第二节和第三节粗;TNW(1、2和3):首部第一节、第二节和第三节粗。Note:BNL(1,2,3):the first(second,third) node length from bottom;TNL(1,2,3):the first(second,third) node length from top;BNW(1,2,3):the first(second,third) node wide from bottom;TNW(1,2,3):the first(second,third) node wide from top.图1 谷子6个茎杆特性的主成分分析Fig.1 Principal component analysis of 6 node traits
2.3 基于茎杆特性的谷子品种多样性分析
2.3.1 基于茎杆特性的主成分分析 根据测量节位的节长和节粗对不同品种进行主成分分析,见表7。前4个主成分的特征值均大于1,累计贡献率达到79.84%。PV(1)的贡献率最大(28.20%),其次是PV(2),最后是PV(5)。品种1对PV(1)贡献较大,品种25对PV(2)贡献较大,品种18对PV(3)的贡献率较大,品种2对PV(4)的贡献率较大。进一步对31个对谷子品种(系)计算欧式距离,利用离差平方和法进行聚分析,如图2可将谷子品种分为两类。第一类包括品种1、16、18、5、29、17、6、10、2、9、30、26、27、12;第二类包括品种3、19、14、15、23、4、22、7、13、25、31、8、20、21、24、28,分类结果说明31个谷子新品种(系)在茎杆特性上存在差异,且不同品种之间茎杆特性存在一定的相似性,可作为31份谷子新品种(系)茎杆特性及抗倒伏能力差异的依据。

表7 不同品种茎杆特性的主成分分析

图2 谷子品种茎杆特性的聚类分析Fig.2 Cluster analysis of node traits
2.3.2 不同品种茎杆多样性与产量性状的分析 对参试材料的茎杆不同节位节粗和节长计算变异系数和多样性指数,并与单穗粒重进行相关分析,见表8。单穗粒重与茎杆粗H′和茎杆长CV呈正相关(P<0.05),且与茎杆粗CV和茎杆长H′呈负相关(P<0.05);茎杆粗H′与茎杆粗CV呈负相关(P<0.05),茎杆长CV与茎杆长H′呈负相关(P<0.05)。利用单穗粒重与茎杆特性进行回归分析(图3),茎杆长H′越高,单穗粒重越低(图3A),茎杆长CV越大,单穗粒重越大(图3B),茎杆粗H′越高,单穗粒重越高(图3C),茎杆粗CV越高,单穗粒重越低(图3D)。
3 讨 论
3.1 谷子农艺性状的多样性
谷子农艺性状的多样性研究主要集中在产量性状[24],例如单穗粒重、穗长、穗粗等,这些性状共同构成产量的主要因素,其遗传变异的信息可以作为亲本材料选择的依据[25]。本研究中31个谷子新品种(系)的8个分级性状和8个农艺性状表现出丰富的变异,各性状的变化幅度、变异系数和多样性指数均较高,其中单穗重和穗粒重与产量密切相关,变异系数为23.794%和24.852%,多样性指数分别为2.170和2.165,穗型和穗密度的多样性指数为1.022和1.237,说明31个谷子新品种(系)农艺性状变异丰富,多样性较高,可以进一步在育种中进行应用,提高谷子新品种(系)的效率。然而,31个新品种(系)的群体小,遗传基础狭窄,而且谷子产量等相关性状的变异受环境影响较大[26],不同的产区应根据农艺性状的实际水平进行种质改良和创制,有条件的产区可引入外来种质资源,拓宽遗传基础。

表8 茎杆多样性与产量性状的相关性分析
注:*代表在P<0.05水平差异显著,下同。
Note:* A significant level atP<0.05, the same as below.

图3 茎杆长和粗与单穗粒重的相关性分析(A:茎杆长H′,B:茎杆长CV, C: 茎杆粗H′,D:茎杆粗CV)Fig.3 Seed weight and genetic diversity of node from different varieties(A:H′ of node length, B: CV of node length, C: H′ of node wide, D: CV of node wide)
3.2 谷子茎杆性状的多样性
作物表型性状的多样性分析包括物候期、产量构成因素和植株性状[27-28]等,而对茎杆相关方面的研究重要集中在新种质创新上[29]。本研究中精确测量谷子新品种(系)不同节位节长和节粗,节长中平均变异系数25.586%,变化幅度较大,平均多样性指数1.972,多样性丰富;节粗中,平均变异系数20.550%,平均多样性指数1.798,多样性丰富;不同节位节粗群集性较强,节长群集性较差;不同品种的茎杆性状对31个谷子品种(系)进行聚类,可将谷子群体分为两类,说明谷子茎杆性状具有丰富的遗传多样性,且不同谷子新品种(系)在茎杆特性上具有一定的相似性。
3.3 谷子茎杆特性与产量性状
作物产量是由品种、环境、栽培管理和病虫害防控等因素综合作用的结果,而品种是影响产量的重要因素[30-31]。在品种农艺性状中,地上部分的植株性状和地下部分的根系作为重要的产量影响因素常被用作新品种选育的重要参考指标[32-33],其中株高、穗长、穗粗、穗密度、出谷率等是谷子育种中进行单株选择的重要依据[34]。本研究对产量主要性状单穗粒重与茎杆特性进行相关分析,单穗粒重与茎杆特性的H′和CV存在显著的相关关系(P<0.05),单穗粒重与茎杆粗H′和茎杆长CV呈正相关,单穗粒重与茎杆粗CV和茎杆长H′呈负相关。这些结果说明茎杆特性也是影响谷子产量构成的重要因素之一,区别于以往农艺性状与产量关系的研究[35-36],谷子茎杆特性在种质创制和育种实践中应给于足够的重视。值得注意的是31个谷子新品种(系)群体茎杆粗H′和CV呈负相关(r=-0.40,P<0.05),茎杆长H′和CV呈负相关(r=-0.41,P<0.05),可能与茎杆的遗传基础存在一定的关联,其内在的机制还需进一步的研究。
4 结 论
31个谷子新品种(系)8个分级性状(苗色、幼苗叶资、株型、花药颜色、穗型、穗码密度、籽粒颜色和米色)和8个农艺性状(生育期、株高、穗长、穗粗、单穗重、穗粒重、出谷率和千粒重)变异幅度较大,群体多样性丰富,依据茎杆特性可将谷子新品种(系)划分为两大类群,且茎杆长和粗具有较高的遗传变异水平,茎秆粗H′和茎秆长CV与单穗粒重呈正相关,茎秆粗CV和茎秆长H′与单穗粒重呈负相关,可作为未来谷子抗倒伏和高产育种重要农艺性状的补充。
参考文献:
[1] 李斌, 顾万春, 卢宝明. 白皮松天然种群种实性状表型多样性研究[J]. 生物多样性, 2002,10(2):181-188.
[2] 尹明宇,姜仲茂,朱绪春,等.内蒙古山杏种群表型变异[J].植物生态学报,2016,40(10):1090-1099.
[3] Jia G Q,Huang X,Zhi H,Diao X,et al. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setariaitalica) [J].Nature Genetic, 2013, 45: 957-961.
[4] 李润枝,陈晨,张培培.我国燕麦种质资源与遗传育种研究进展[J].现代农业科技,2009,17:44-45.
[5] 代攀虹,孙君灵,何守朴,等. 陆地棉核心种质表型性状遗传多样性分析及综合评价[J].中国农业科学,2016,49(19):3694-3708.
[6] 陈雪燕,工亚娟,雒景吾,等. 陕西省小麦地方品种主要性状的遗传多样性研究[J].麦类作物学报,2007,27(3):456-460.
[7] 王果,胡正,张保缺,等.山西省野生大豆资源遗传多样性分析[J].中国农业科学,2008,41(7):2182-2190.
[8] 胡标林,万勇,李霞,等.水稻核心种质表型性状遗传多样性分析及综合评价[J].作物学报,2012,38(5):829-839.
[9] 蔡一林,刘志斋,王天宇,等.国内部分玉米地方品种的品质与农艺性状的表型多样性分析[J].植物遗传资源学报,2011,12(1):31-36.
[10] 王继师,刘祖昕,樊帆,等.24个甜高粱品种主要农艺性状与品质性状遗传多样性分析[J].中国农业大学学报,2012,17(6):83-91.
[11] 南铭,马宁,刘彦明,等.燕麦种质资源农艺性状的遗传多样性分析[J].干旱地区农业研究,2015,33(1):262-267.
[12] 高金锋,张慧成,高小丽.西藏苦荞种质资源主要农艺性状分析[J].河北农业大学学报,2008,31(2):1-5.
[13] 董俊丽,王海岗,陈凌,等.糜子骨干种质遗传多样性和遗传结构分析[J].中国农业科学,2015,48(16):3121-3131.
[14] 栾素荣,王占廷,李青松.谷子产量与主要农艺性状的灰色关联度分析[J].河北农业科学,2010,14(11):115-116,118.
[15] 赵禹凯,王显瑞,张立媛,等.谷子产量与主要农艺性状的灰色关联度分析[J].吉林农业科学,2014,39(2):9-12.
[16] 杨成元.春谷产量与其相关性状的关联度分析[J]. 杂粮作物,2002,22(5):259-261.
[17] 黄英杰,张岩.谷子品种产量及主要产量构成因素稳定性的分析[J].作物杂志,2002,5:43-44.
[18] 赵禹凯,王显瑞,陈高勋,等.谷子主要农艺性状的相关和通径分析[J].内蒙古农业大学学报,2014,35(2):35-38.
[19] 王海岗,贾冠清,智慧,等.谷子核心种质表型遗传多样性分析及综合评价[J].作物学报,2016,42(1):19-30.
[20] 刁现民. 基础研究提升传统作物谷子和黍稷的科研创新水平[J].中国农业科学,2016,49(17):3261-3263.
[21] 孟庆立,关周博,冯佰利,等.谷子抗旱相关性状的主成分与模糊聚类分析[J].中国农业科学,2009,42(8):2667-2675.
[22] 陆平.谷子种质资源描述规范和数据标准[M].北京:中国农业出版社,2006.
[23] 屈洋,周瑜,王钊,等.苦荞产区种质资源遗传多样性和遗传结构分析[J].中国农业科学,2016,49(11):2049-2062.
[24] 田伯红.谷子地方品种和育成品种的遗传多样性研究[J].植物遗传资源学报,2010,11(2):224-228.
[25] 杨天育,黄相国,何继红,等.谷子遗传资源多样性研究进展[J].西北农业学报,2003,12(1):43-47.
[26] Qu Y, Zhou Y, Chao G, et al. Water use efficiency of foxtail millet (PanicumitalicumL.) under climate change conditions in Northwest regions of China[J]. Agrociencia,2016, 50: 665-676.
[27] 刁现民,程汝宏.十五年区试数据分析展示谷子糜子育种现状[J].中国农业科学,2017,50(23):4469-4474.
[28] 张艾英,郭二虎,刁现民,等.2005-2015年西北春谷中晚熟区谷子育成品种评价[J].中国农业科学,2017,50(23):4486-4495.
[29] 刘正理,程汝宏,黄文胜,等.甜秆多穗型超早熟谷子新种质的创新[J].中国农业科学,2005,38(1):17-21.
[30] 刘正理,程汝宏,李香月,等.不同类型谷子品种叶片对产量形成效应的研究[J].中国农学通报,1995,11(3):12-16.
[31] 夏雪岩,师志刚,张婷,等.不同粒色谷子品系的鸟害程度研究[J].河北农业科学,2014,18(2):4-6.
[32] 张文英,智慧,柳斌辉,等.豫谷1号和青狗尾草RIL群体根系变异和垂直分布[J].作物学报,2014,40(10):1717-1724.
[33] 刘正理,程汝宏.谷子目标性状基因库育种技术体系的构建及其应用[J].中国农业科学,2005,38(7):1306-1311.
[34] 程汝宏,籍贵苏,李香月.夏谷高产育种单株选择模式[J].华北农学报,1996,11(1):123-127.
[35] 陈素省,赵国顺,郝艳敏.应用灰色关联度法评价谷子新品种[J].河北农业科学,2006,10(2):62-65.
[36] 刘正理,程汝宏,张凤莲,等.华北夏谷区主要谷子品种遗传基础分析及改良途径的探讨[J].云南农业大学学报,2006,21(增刊):84-90,99.
猜你喜欢
西北农业学报(2022年9期)2022-10-08
西南农业学报(2022年1期)2022-03-15
辽河(2021年12期)2021-12-24
中国农业气象(2021年7期)2021-07-28
麦类作物学报(2021年3期)2021-05-25
北京农学院学报(2020年1期)2020-03-09
麦类作物学报(2018年10期)2018-12-12
三联生活周刊(2016年35期)2016-08-29
人间(2015年25期)2015-01-17
文苑·感悟(2008年4期)2008-07-07
