
喀斯特白云岩石漠化区域不同生境条件下苔藓植物群落特征及演替模式研究
2018-07-02 11:20庞嘉鹏王智慧张朝晖
生态科学 2018年3期
庞嘉鹏, 王智慧,*, 张朝晖
1. 贵州师范大学生命科学学院, 贵阳 550001
2. 贵州师范大学贵州省山地环境信息系统与生态环境保护重点实验室, 贵阳 550001
1 前言
石漠化是喀斯特地貌典型的脆弱生态环境, 主要集中在以贵州为中心(北纬23°—32°, 东经102°—111°)的中国西南地区[1-2], 具有岩石裸露、易水土流失、生态环境恶劣等特点[3-4]。而生态环境是植物的生长所依赖的一个重要条件, 这种特殊的环境使得植被演替放缓, 生态效率降低[5]。苔藓植物作为植物界从水生向陆生过渡的高等植物中最原始的类群,即使在这样养分限制和总体生产力较低的喀斯特恶劣生态环境下, 依然可以持续生长。由于本身植株微小, 加上物种鉴定困难以及大部分研究者对苔藓植物缺乏了解, 对于苔藓植物的功能往往被忽视[6]。国内关于喀斯特苔藓植物的研究主要集中在在分类、区系、多样性、生态功能等方面[7-13]。喀斯特地形这种特殊的岩溶地质, 石漠化较严重, 水土保持能力较弱, 苔藓群落作为石漠化环境的先锋植物群落[14], 具有水土保持、环境保护、重金属富集等生态功能[15]。陈云等采用多元回归树得出小秦岭自然保护区苔藓植物多样性随海拔的上升呈显著的上升格局[16]。刘荣相等对贵州贞丰县喀斯特石漠地区峰丛不同山段苔藓植物群落特征的研究, 说明人类影响及植被覆盖率成为了影响苔藓分布的重要因素[17],籍烨研究不同演替阶段中苔藓植物的多样性, 发现随着演替阶段的发展, 苔藓植物多样性呈现先下降、在藤刺灌丛出现反弹、最后再次降低的变化趋势[18]。通过对群落多样性的研究, 不仅有利于了解一定区域范围内系统本身抵御外界不确定性因素的干扰和干扰后恢复的能力, 而且还有助于探究喀斯特白云岩石漠化区域生态系统中苔藓群落的生产力水平[19-22], 因此本文以苔藓群落作为对象, 通过对喀斯特白云岩石漠化区域不同生境条件下的苔藓群落的种类组成、群落多样性特征及其演替模式的研究,揭示了苔藓群落在生境条件下的适应环境、维持生存的策略, 为喀斯特石漠化的生物治理提供研究依据。
2 研究区域和方法
2.1 研究地点
研究地区位于贵州省贵阳市城北黔灵山脉的鹿冲关森林公园(北纬 26°37′31.01″—26°38′09.80″, 东经 106°42′09.50″—106°43′16.27″), 该区域是发育在白云岩上的十分典型的喀斯特地形[23], 处于亚热带湿润温和型气候带, 有充沛的水资源、较稳定且适宜的温度以及较大的湿度, 使得这里的森林覆盖率达87%, 共有植物约190科844属2037种, 主要植被群落类型有以马尾松(Pinus massoniana)为优势的森林群落, 主要植物种类有马尾松、枫香树(Liquidambar formosana)、油茶(Camellia oleifera)、杜鹃(Rhododendron simsii)和小果珍珠花(Lyonia ovalifolia)等; 以华山松(Pinus armandii)为优势的群落, 主要植物种类有华山松、盐肤木(Rhus chinensis)、水杉(Metasequoia glyptostroboides)、朴树(Celtis sinensis)、栾树(Koelreuteria paniculata)等; 以黑松(Pinus thunbergii)为优势的群落, 主要植物种类有黑松、小果蔷薇(Rosa cymosa)、金丝梅(Hypericum patulum)、火棘(Pyracantha fortuneana)等。包括掌叶木(Handeliodendron bodinieri)、云南拟单性木兰(Parakmeria yunnanensis)、银杉(Cathaya argyrophylla)和柔毛油杉(Keteleeria pubescens)等濒危稀有植物约41种[24]。由于人类活动的影响, 样地内有局部地区出现了喀斯特石漠化现象, 同时经初步调查,区域内有较为丰富的典型植被、生境条件, 苔藓植物生长良好、种类丰富, 为研究苔藓群落多样性提供了优越的条件。
2.2 调查及采样
于2015年10月—2016年4月间采用样方调查法进行调查。根据植物类群和环境选取裸露白云岩、灌草丛、藤刺灌木丛、针阔混交林四种生境条件, 设置的样地见图1。同时记录每个样地海拔、温度、湿度、光照强度、郁闭度、盖度等生态因子(表1)。每个样地随机取5个1 m×1 m大样方, 每个大样方利用五点取样法取5个10 cm×10 cm小样方中全部苔藓, 共收集群落样方(样本)100份。实验室内使用HWG-1型双筒解剖镜、XSZ-107TS型光学显微镜观察并绘图。参照《中国苔藓志》(第二、四、七、八卷)进行苔藓物种鉴定[25-28], 并对每个小样方苔藓植物群落利用优势种原则命名。参照《贵州植物志》[29](第一、四、五、七、九卷)对样地植物进行分类鉴定。
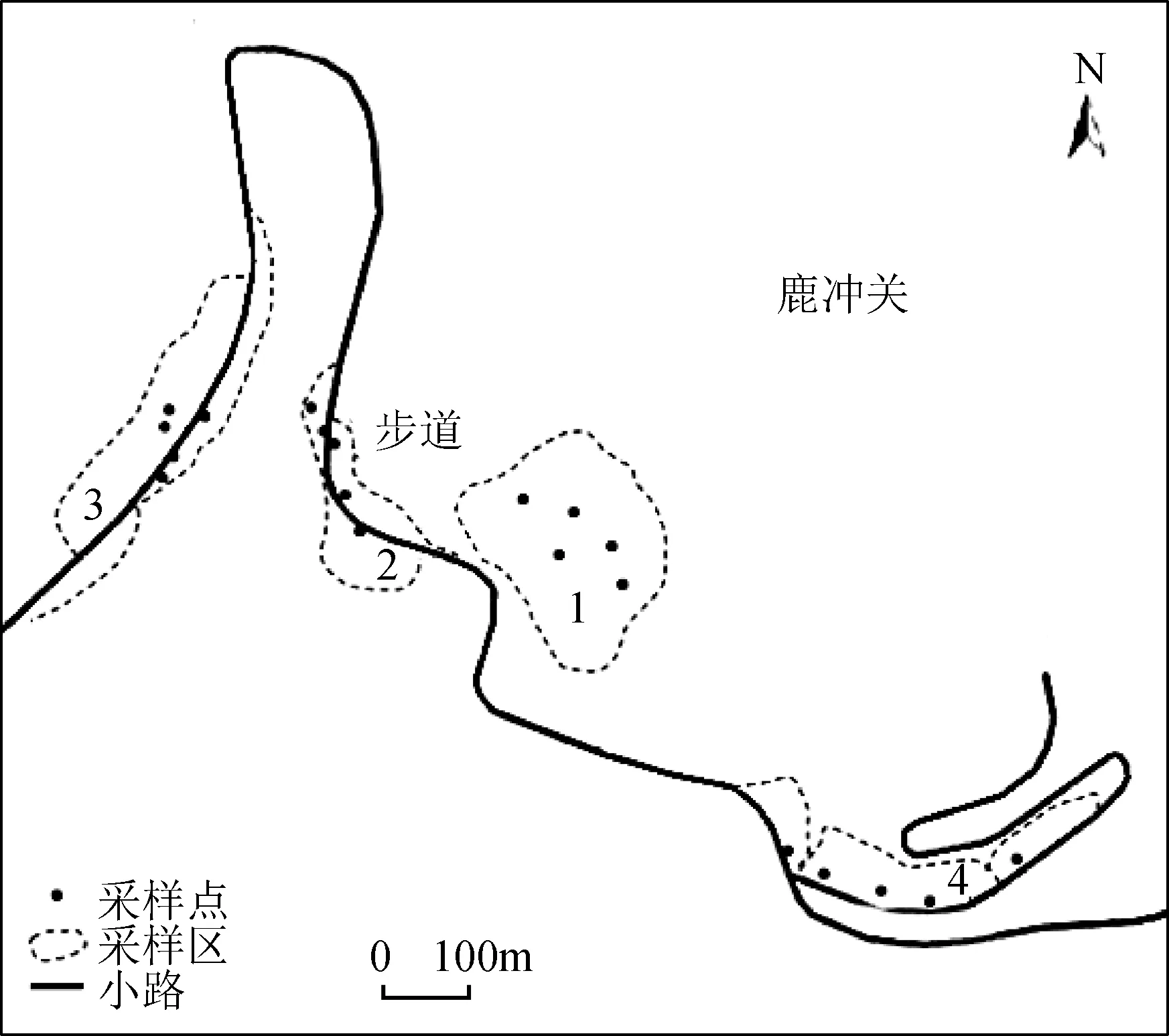
图1 研究区采样点设置Fig. 1 Sample location of area
2.3 数据分析
2.3.1 重要值(IV)
苔藓植物的重要值:IV(%)=(Fr+Cr)/2
式中,Fr为苔藓植物在各样地的相对频度,Cr为苔藓植物在各样地的相对盖度。
2.3.2 多样性指数
(1)α多样性指数
Shannon-Wiener多样性指数(H)
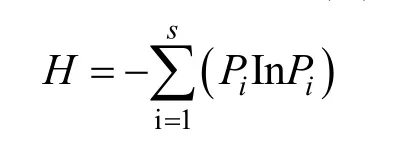
式中,H为各小样方物种多样性之和,Pi为第i个物种的重要值,S为每个生境条件内的苔藓物种总数。
Pielou均匀度指数(J)

式中,H′为Shannon-Wiener多样性指数。
(2)β多样性指数
Sorenson多样性指数(Cs)

式中,j为两个生境条件下苔藓植物的共有种,a和b分别为两个生境条件下苔藓植物的物种总数。
3 结果与分析
3.1 群落物种组成
经调查, 通过对各样地内苔藓群落的统计, 共记录到组成物种11科30属82种, 其中藓类植物10科28属80种, 苔类植物1科1属2种。裸露白云岩共有苔藓植物27种, 隶属于3科11属; 灌草丛共有苔藓植物27种, 隶属于4科14属; 藤刺灌木丛共有苔藓植物34种, 隶属于6科16属; 针阔混交林共有苔藓植物42种, 隶属于7科21属, 其中优势科为丛藓科(Pottiaceae)、真藓科(Bryaceae), 优势属为扭口藓属(Barbula)、真藓属(Bryum)、小石藓属(Weissia)和石灰藓属 (Hydrogonium)等。就总体而言, 随着生境条件逐渐良好, 物种丰富度逐渐增加。裸露白云岩和灌草丛仅有27个种, 物种丰富度最小。针阔混交林苔藓植物十分丰富, 其科、属、种相较其他三个植被类型也是最大的。
3.2 各生境条件苔藓植物重要值

表1 样地生境Tab. 1 Environmental features of sampling sites
从群落物种组成上来看, 短叶小石藓、北地扭口藓和尖叶扭口藓普遍存在于各生境条件下, 为各生境条件的共有种。仅在针阔混交林出现的有卵叶青藓、侧枝匐灯藓和短尖美喙藓(Eurhynchium angustirete)等21种, 仅出现在藤刺灌木丛的包括有净口藓(Gymnostomum calcareum)、宽叶真藓(Bryum funkii)和中华短月藓(Brachymenium sinense)等13种,比拉真藓(Bryum billarderi)、短肋羽藓(Thuidium kanedae)等9种仅出现在灌草丛, 仅出现在裸露白云岩的最少, 仅阔叶小石藓、尖叶平蒴藓(Plagiobryum demissum)、沙氏真藓(Bryum sauteri)等8种。

图2 各生境条件苔藓植物科属种数Fig. 2 The number of family and genus in different habitats
针阔混交林下的优势苔藓植物为北地扭口藓、卵叶青藓、侧枝匐灯藓、毛口藓(Trichostomum brachydontium), 其中北地扭口藓出现最多, 为建群种, 其重要值为 14.65, 藤刺灌木丛下的长尖扭口藓、卷叶石灰藓、尖叶扭口藓、净口藓为优势种, 其重要值分别为 8.23、7.65、6.99、5.48, 灌草丛中的长尖扭口藓重要值达到 10.82, 其次是黑扭口藓(Barbula nigrescens)和银叶真藓, 重要值为 7.59和7.24, 其余种类重要值介于1.48—6.53之间, 裸露白云岩中北地扭口藓重要值为 9.86, 其次是阔叶毛口藓和长尖扭口藓, 重要值为 9.85和 9.42, 小石藓(Weissia controversa)的重要值最小仅0.85。
3.3 各生境条件苔藓群落多样性分析
各生境条件苔藓植物的Shannon-Wiener多样性指数(H)和Pielou均匀度指数(J)如图3所示。由图可知, 各生境条件多样性指数总体呈现由裸露白云岩到针阔混交林不断递增的趋势, 与丰富度的关系最为密切, 针阔混交林物种多样性指数最高, 为 3.37;藤刺灌木丛和草灌丛次之, 分别为 3.31和 3.13; 裸露白云岩的物种多样性指数最低, 仅为 2.99, 这与该生境条件较为恶劣有关。总体来说, J值均大于0.9,说明整体研究区域的生境较好, 而在裸露白云岩和针阔混交林两个“极端”生境中均匀度最小, 说明石漠化生境和高等植物的顶级群落, 均对苔藓的生长产生了影响。其中藤刺灌木丛、灌草丛、裸露白云岩三者之间无显著性差异(P>0.05); 针阔混交林与藤刺灌木丛二者之间也无显著性差异; 而针阔混交林与灌草丛、裸露白云岩之间具有显著差异(P<0.05)。
各生境条件苔藓群落物种组成的相似性如表 3所示。各生境条件苔藓植物群落物种相似性相对较低(26.09%—45.90%), 均未达到50%。其中藤刺灌木丛和裸露白云岩之间的相似性最高。达到了45.90%,表明其苔藓物种组成最为相似。针阔混交林与裸露白云岩相似性系数最低, 为 26.09%, 灌草丛和裸露白云岩相似性也较高, 达到了40%。
3.4 各群落生境条件苔藓植物生活型
根据 Magdefrau的概念和区分系统[30], 通过统计各生境条件苔藓植物的生活型, 各生境条件下共有4种类型, 其中矮丛集型达到了苔藓总数的50%,主要包括丛藓科、真藓科等, 高丛集型占苔藓总数的 33.33%, 主要包括金发藓属、扭口藓属等, 交织型有 11种, 占苔藓总数的 13.10%, 主要包括青藓科、羽藓科等, 平铺型和树状型最少,仅地钱属 2种和大叶藓属 1种, 分别占苔藓总数的 2.38%和1.19%。该区域内矮丛集型、高丛集型、交织型是苔藓植物主要生活型, 反映出该区域各生境条件环境复杂, 各生境条件多样化特点。
各生境条件的生活型也具有差异(图4), 由图可以看出, 矮丛集型、高丛集型在各生境条件中均有出现, 交织型除了藤刺灌木丛均有出现, 平铺型仅少量出现在藤刺灌木丛和针阔混交林, 树状型只出现于灌草丛其余生境条件均未出现该生活型, 针阔混交林相较于其他生境, 交织型的比例增加, 高丛集型比例减少。裸露白云岩, 植被类型单一, 生存环境较恶劣, 分布有3种生活型, 物种种类较单一;针阔混交林, 植被郁闭度大, 被乔木重新分配的湿度、光照等资源使得苔藓植物种类达到最多, 同时苔藓植物生活型也最丰富, 被视为潮湿环境指示的交织型苔藓比例在这个生境明显增加。
3.5 各生境条件苔藓群落结构
如表 4所示, 以优势种命名, 各生境条件共有99个苔藓群落, 其中纯群落16个, 混合群落83个,混合群落数量远远大于纯群落, 说明该区域受到人类的干扰较小, 苔藓植物种类丰富。其中灌草丛纯群落: 混合群落之比最高, 表明在该生境下, 环境
群落结构相对稳定。可以看出扭口藓属的苔藓在不同的生境条件均可以构成优势群落, 表明其适应能力强, 而不同的环境条件下的优势群落也有区别,对于环境较为恶劣的裸露白云岩, 就会产生如银叶真藓的矮丛集型耐旱苔藓植物群落; 而对于针阔混交林环境湿润但同时荫蔽弱光, 就会产生青藓属等匍匐型的苔藓植物群落。

表2 各生境条件苔藓植物群落物种组成及重要值Tab. 2 Composition and importance value in different habitats

图3 各生境条件物种多样性指数比较Fig. 3 Diversity index comparison in different habitats

表3 各生境条件β多样性指数差异Tab. 3 β-diversity index of bryophytes in different habitats %

图4 各生境条件物种生活型Fig. 4 Percentages of different life form in different habitats
3.6 苔藓群落演替模型
以各苔藓物种重要值以及植物样方使用CANOCO 4.5 for Windows进行PCA排序, 排序结果较好的反映出各生境条件的生态关系和种类组成的变化(图5)。图中不同的箭头代表不同的生境条件,TH为针阔混交林,TB为藤刺灌木丛,SG为灌草丛,DB为裸露白云岩; 每一个圆形代表一个物种, 图中左侧的为小于平均贡献的种, 右边为大于平均贡献的种, 因此仅讨论盖度和频度均较高的群落组成。从图中可以看出, 在各个生境条件下, 可以明显的划分为3个区域, 对应3个演替阶段, 每个阶段具有与其相关性较高的群落物种组成, 其中与针阔混交林相关性较高的包括以北地扭口藓为建群种的群落、卵叶青藓、侧枝匐灯藓、毛口藓 (Trichostomum brachydontium)构成的群落, 灌草丛有长尖扭口藓、黑扭口藓、银叶真藓、短叶小石藓、丛本藓; 丛本藓; 与裸露白云岩相关的苔藓植物有尖叶扭口藓、剑叶扭口藓; 藤刺灌木丛仅阔叶毛口藓一种。即随着生境条件的改善, 在植被、水热条件等的影响下,各阶段建群种演替规律为: 尖叶扭口藓→长尖扭口藓→北地扭口藓。在生境较为恶劣的阶段以尖叶扭口藓为建群种, 与剑叶扭口藓、银叶真藓等伴生种组成混合群落, 伴生种比较丰富; 过渡阶段随着环境改善,以长尖扭口藓为主要建群种的群落分化更为明显;顶级的针阔混交林下, 各个种类基本各自成群, 除北地扭口藓的优势度最高以外, 卵叶青藓、侧枝匐灯藓、毛口藓等也可以形成相应的群落。可见在群落演替模式上, 受到生境条件的影响比较明显。

表4 各生境条件苔藓群落结构Tab. 4 Community structure of bryophytes in different habitats

图5 样方和物种PCA排序Fig. 5 PCA ordination of species and habitats
4 结论与讨论
喀斯特白云岩石漠化区域苔藓群落种类组成多样化。本研究区域苔藓群落组成物种共有11科30属82种, 其中藓类植物10科28属80种, 苔类植物1科1属2种。构成了喀斯特石漠化区域的苔藓群落, 占优势的科是丛藓科和真藓科; 苔藓植物群落共99个, 其中混合群落占83.8%, 纯群落仅占16.2%,说明总体区域环境良好。随着生境条件的改善, 苔藓群落的物种丰富度不断增加, 显示了苔藓植物生活环境随各类大型高等植物的改变不断改善。
苔藓群落特征与各生境条件密切相关。在本研究区域内, 矮丛集型占苔藓种类的 50%, 且在各个生境条件均有出现, 这与其植株短小, 对环境适应性强有关, 反应了矮丛集型苔藓较好的耐受性, 其中短叶小石藓存在数量大、在各生境条件均有出现,表明其相较于其他苔藓具有更强的生存能力和更宽的生态位, 这种在各生境条件具有较强竞争力的苔藓种类对喀斯特地区石漠化的恢复治理及水土保持具有重要意义[31-32]。而针阔混交林生境, 郁闭度大,被乔木重新分配的湿度、光照等资源使得苔藓植物生活型种类达到 4种, 被视为潮湿环境指示的交织型苔藓比例明显增加, 达到了到 16.36%, 这也与Glime的研究观点相同[33]。
生境条件变化对于苔藓植物相较于其他高等植物的影响更为明显[34-36]。苔藓植物特有的生活史以及不同的苔藓植物也由于生态适应对策不同对生境条件有不同的响应策略, 决定了苔藓植物群落的组成和演替模式。多项研究表明, 苔藓植物对于生境的变化并非具有非常吻合的变化方式, 在生境从恶劣到较好的过渡阶段, 物种多样性随着生境条件的改善呈正相关, 但随后会又会出现下降趋势[18,37]。本研究 Shannon-Wiener多样性指数(H)在各生境条件多样性指数总体呈现由裸露白云岩到针阔混交林不断递增的趋势, 与其丰富度密切相关; 在裸露白云岩和针阔混交林两个“极端”生境中均匀度最小, 在灌草丛最大, 分化出以优势种占据的群落增加, 从而导致群落间相同种类减少的现象。表明灌草丛这一生境条件, 为苔藓提供了一个优势的改造环境生境, 多样性增加; 而针阔混交林顶级的高等植物群落出现, 环境条件得到再分配, 苔藓植物净光合作用减弱[38], 种间竞争加剧, 使苔藓多样性趋于减小,适应环境的优势种、建群种逐渐被自然选择脱颖而出。由此可见灌草丛在石漠化生境条件中的重要地位, 可以考虑作为苔藓石漠化修复的背景环境, 重点关注, 这也为石漠化治理提供了新思路。在群落演替模式上, 本研究区域四种生境条件下的演替模式尖叶扭口藓、长尖扭口藓、北地扭口藓成为了演替模式种, 但在各生境条件下还是有差异, 可以预见随着生境条件继续改善, 多种优势种将趋于平均分配生态空间, 形成以多种苔藓为优势种少量伴生种的群落模式。
[1] 陈洪松, 聂云鹏, 王克林. 岩溶山区水分时空异质性及植物适应机理研究进展[J]. 生态学报, 2013, 33(2):317-326.
[2] 李阳兵, 侯建筠, 谢德体. 中国西南岩溶生态研究进展[J]. 地理科学, 2002, 22(3): 365-370.
[3] 张信宝, 王世杰, 曹建华, 等. 西南喀斯特山地水土流失特点及有关石漠化的几个科学问题[J]. 中国岩溶, 2010,29(3): 274-279.
[4] 何永彬, 张信宝, 李豪. 喀斯特白云岩坡地土壤异质性特征与土壤保育模式研究[J]. 地球与环境, 2013, 41(1):77-81.
[5] 姚长宏, 蒋忠诚, 袁道先. 西南岩溶地区植被喀斯特效应[J]. 地球学报, 2001, 22(2): 159-164.
[6] 田维莉, 孙守琴. 苔藓植物生态功能研究新进展[J]. 生态学杂志, 2011, 30(6): 1265-1269.
[7] 曹同, 郭水良. 长白山主要生态系统苔藓植物的多样性研究[J]. 生物多样性, 2000, 8(1): 50-59.
[8] 唐景儒, 杨比伦, 卢廷, 等. 铜壁关自然保护区陇川片区苔藓植物资源调查[J]. 西南林学院学报, 2006, 26(2):24-28.
[9] 汪庆, 贺善安, 吴鹏程. 苔藓植物的多样性研究[J]. 生物多样性, 1999, 7(4): 332-339.
[10] 郝占庆, 叶吉, 姜萍, 等. 长白山暗针叶林苔藓植物在养分循环中的作用[J]. 应用生态学报, 2005, 16(12): 2263-2266.
[11] 白学良, 赵连梅, 孙维, 等. 贺兰山苔藓植物物种多样性、生物量及生态学作用的研究[J]. 内蒙古大学学报(自然科学版), 1998, 29(1): 121-127.
[12] TILMAN D, REICH P B, KNOPS J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment[J]. Nature, 2006, 441(7093): 629-632.
[13] IVES A R, CARPENTER S R. Stability and diversity of ecosystems[J]. Science, 2007, 317(5834): 58-62.
[14] 王圳, 张金池, 于水强, 等. 退化喀斯特地区植被恢复过程中苔藓的先锋作用[J]. 南京林业大学学报(自然科学版), 2011, 35(3): 137-140.
[15] 贾少华, 李军峰, 王智慧, 等. 喀斯特山区公路石漠化边坡苔藓生态功能[J]. 生态学杂志, 2014, 33(7):1928-1934.
[16] 陈云, 冯佳伟, 牛帅, 等. 小秦岭自然保护区苔藓植物群落数量分类、排序及多样性垂直格局[J]. 生态学报, 2017,37(08): 2653-2664.
[17] 刘荣相, 王智慧, 张朝晖. 贵州贞丰喀斯特石漠峰丛苔藓植物群落生态特征[J]. 植物研究, 2009, 29(6):734-741.
[18] 籍烨, 张朝晖. 喀斯特石漠生态系统不同自然演替阶段中苔藓植物多样性特征分析[J]. 植物科学学报, 2014,32(06): 577-585.
[19] NAEEM S, THOMPSON L J, LAWLER S P, et al.Declining biodiversity can alter the performance of ecosystems[J]. Nature, 1994, 368(6473): 734-737.
[20] KENNEDY T A, NAEEM S, HOWE K M, et al.Biodiversity as a barrier to ecological invasion[J]. Nature,2002, 417(6889): 636-638.
[21] 王长庭, 龙瑞军, 丁路明, 等. 草地生态系统中物种多样性、群落稳定性和生态系统功能的关系[J]. 草业科学,2005, 22(6): 1-7.
[22] FELLY M T, STEVEN M B, JEFF W B. Diversity and distribution of bryophytes in three forest types of Bwindi Impenetrable National Park, Uganda[J]. African Journal of Ecology, 2007, 45(3): 79-87.
[23] LAI Wenting, WANG Zhihui, ZHANG Zhaohui. The bryophytes found in artificial trench in karst mountainous area in Guizhou province of China[J]. Chenia, 2016, 1(12):109-114.
[24] 储蓉, 郭鸿英. 贵州省植物园珍稀濒危保护植物的迁地保护初报[J]. 贵州林业科技, 2004, 32(2): 38-42.
[25] 高谦. 中国苔藓志: 第 2 卷[M]. 北京: 科学出版社, 1996:1-293.
[26] 高谦. 中国苔藓志: 第 4 卷[M]. 北京: 科学出版社, 2006:1-263.
[27] 胡人亮, 王幼芳. 中国苔藓志: 第 7 卷[M]. 北京: 科学出版社, 2005: 1-288.
[28] 吴鹏程, 贾渝. 中国苔藓志: 第 8 卷[M]. 北京: 科学出版社, 2004: 1-482.
[29] 贵州植物志编委会. 贵州植物志: 第 1、4、5、7、9 卷[M].贵阳: 贵州人民出版社, 1982.
[30] MAGDEFRAU K. Bryophyte ecology[M]. London:Chapmanand Hall, 1982: 45-58.
[31] 李军峰, 贾少华, 王智慧, 等. 喀斯特石漠化过程中苔藓植物多样性及分布与环境关系[J]. 生态科学, 2015, 34(1):68-73.
[32] 李军峰, 王智慧, 张朝晖. 喀斯特石漠化山区苔藓多样性及水土保持研究[J]. 环境科学研究, 2013, 26(7):759-764.
[33] WANG Zhe, LIU Xin, BAO Weikai. Higher photosynthetic capacity and different functional trait scaling relationships in erect bryophytes compared with prostrate species[J].Oecologia, 2016, 180(2): 359-369.
[34] DA C D P. Epiphytic bryophyte diversity in primary and secondary lowland rainforests in southeastern Brazil[J].The Bryologist, 1999, 102(2): 320-326.
[35] 田晔林, 王文和, 颜亭玉, 等. 北京百花山自然保护区不同植被地面生苔藓植物物种多样性[J]. 植物研究, 2013,33(4): 398-403.
[36] 田晔林, 李俊清, 石爱平, 等. 北京百花山自然保护区树附生苔藓植物物种多样性[J]. 生态学杂志, 2013, 32(4):838-844.
[37] ODUM E P. The strategy of ecosystem development[J].Science, 1969, 164(3877): 262-270.
[38] 郭磊, 韦博良, 胡金涛, 等. 基于两个不同资源轴上苔藓植物生态位分析[J]. 生态学报, 2017, 37(21):1-11.
猜你喜欢
军事文摘(2021年18期)2021-12-02
润·文摘(2021年11期)2021-11-20
今日中国·西班牙文版(2021年4期)2021-04-09
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
现代园艺(2018年2期)2018-03-15
儿童故事画报·自然探秘(2017年7期)2018-03-14
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
