
不同生态环境胁迫胡桃楸的演化结构比较研究
2018-06-27 06:59:56陈丹萍由继红刘丹丹王晓钰徐光照陆静梅
东北师大学报(自然科学版) 2018年2期
陈丹萍,由继红,刘丹丹,王晓钰,李 岩,徐光照,陆静梅
(东北师范大学生命科学学院,吉林 长春 130024)
胡桃楸(JuglansmandshuricaL.)为胡桃科(Juglandaceae)胡桃属(Juglans)植物,是我国珍贵树种[1].其种子内含可供食用的油脂,木材性质优良;又因其青果、树皮及叶内含有的生物活性物质和有效的化学成分较丰富,并对森林生态系统有绿化功能,故被誉为“果材兼备、药赏两佳”的经济树种[2-6].胡桃楸极高的经济价值、药用价值和生态价值,使其有着广阔的开发应用前景,1992年被列为国家二级珍稀树种和中国珍稀濒危树种的三级保护植物[7].近年来,越来越多的国内外学者致力于胡桃楸的栽培技术、化学成分以及生物学活性等方面的研究[8-9].随着城市建设的蓬勃发展和生态环境的恶化,土地资源量相对匮乏问题日益严重,很多土地因处于建筑包围之中而形成荫蔽环境,楼宇旁、树荫下的植物无法正常生长,因此研究和利用植物耐荫性,选育不同耐荫性的树种,相对增加绿化面积,将从根本上解决荫胁地的利用问题,同时可以有效改善人们的生活质量及周围的生态环境.环境决定植物的形态建成,结构又是功能的基础,植物结构的变化必然影响到生理生态功能的改变[10-11].
目前,对于同一种植物颉颃不同生态环境的演化结构研究少有问津,因此,本文通过植物形态解剖研究,探索了植物颉颃逆境适应机制和相应对策的基础;通过系列解剖结构实验,探讨了阴生与阳生环境胁迫下胡桃楸叶片和叶柄解剖结构的演化规律,以为胡桃楸的抗逆研究以及优良品种的选育提供理论基础和科学参考依据.
1 材料与方法
1.1 实验材料
供试材料胡桃楸分别采自吉林省长春市吉林大学校园(阴生环境,43.857°N,125.333°E)和东北师范大学校园(阳生环境,43.862°N,125.331°E).
取材时间均为2016年7月.供试材料为茎的顶端下数第5片复叶和小叶柄.
1.2 实验方法
1.2.1 石蜡切片法
将叶片切成0.3 cm×0.6 cm大小,截取叶柄0.5 cm放入FAA中固定48 h,经不同浓度梯度的酒精逐级脱水,二甲苯逐级透明,浸蜡包埋待凝固后进行修块切片、展片及粘片,烘干后经二甲苯逐级脱蜡,番红-固绿复染法染色制片,最后用中性树胶封片.
1.2.2 显微观察及摄影
使用Olympus光学显微镜镜检.挑选合格的装片在Nikon Eclipse 80i显微镜下观察,并选取目标视野进行不同倍数的显微拍摄.
1.2.3 数据处理及分析
采用Nis-Elements图像分析系统中的Nis-ElementsD2.20,SP2(Build 243) Imaging Software 软件测量胡桃楸叶片和叶柄相关结构特征参数,取10次重复实验的平均值,结果用“平均值±标准差”表示.使用SPSS19.0软件进行数据统计学分析[12].
2 结果与分析
2.1 阴生环境的胡桃楸叶片解剖结构
阴生环境中生长的胡桃楸叶片较薄(见图1-1、图1-7),平均厚度为92.088 μm.表皮细胞参差不齐,平均直径为12.393 μm(见图1-3),外切向壁角质化程度较低,偶见单细胞表皮毛集生在表皮突起的部位(见图1-1、图1-3),腺毛由倒圆锥状的多细胞头部和多细胞柄组成,着生在叶的远轴面(见图1-9).叶主脉的机械组织中有沙晶分布(见图1-3).叶主脉木质部导管孔径较大(见图1-5),排列疏松,其管孔链平均长度为57.590 μm.韧皮部不发达(见图1-1),平均厚度为66.780 μm.叶片栅栏组织厚度较小,平均厚度为44.696 μm,胞间隙中出现不规则的晶体状结构且体积较大,形似钟乳状(见图1-7).海绵组织细胞形状不规则且不发达,平均厚度为21.175 μm,有通气组织存在(见图1-7).
2.2 阴生环境的胡桃楸叶柄解剖结构
阴生胡桃楸叶柄表皮细胞平均直径为6.984 μm.表皮下基本组织细胞较大、细胞壁较薄,特化程度较低,由3~4层细胞组成(见图1-13).维管束排列较松散,为异形维管束,数目较少(见图1-11).木质部导管分子数量少,导管直径大,形成的管孔链长度为91.560 μm,由大小参差不齐的导管分子组成的导管团数量极少(见图1-13).韧皮部的平均厚度为146.736 μm,筛管分子不发达,韧皮纤维特化不显著(见图1-13),平均厚度为72.671 μm.叶柄维管束的中部基本组织细胞呈心形排列,薄壁细胞数量少、直径大(见图1-11).
2.3 阳生环境的胡桃楸叶片解剖结构
阳生胡桃楸叶片厚度为125.834 μm(见图1-2、图1-8).叶表皮细胞平均直径为10.781 μm,外切向壁角质层较厚(见图1-4),表皮毛数量多且粗壮,包括单细胞簇生表皮毛和多细胞表皮毛(见图1-2),腺毛亦由倒圆锥状的多细胞头部和多细胞柄组成(见图1-4、图1-10).叶主脉维管束的机械组织发达,细胞内具有沙晶,木质部导管孔径小、数量多且排列紧密(见图1-6),管孔链长度为66.469 μm.韧皮部较发达(见图1-2),平均厚度为73.571 μm,薄壁细胞中晶体较多(见图1-4).叶片栅栏组织由3层细胞组成,平均厚度为71.698 μm,在栅栏组织细胞中形成了肥大的异形胞,其内具有大量晶体(见图1-8).海绵组织平均厚度为29.064 μm,细胞间隙较大(见图1-8).
2.4 阳生环境的胡桃楸叶柄解剖结构
叶柄表皮细胞平均直径为6.252 μm,表皮下的基本组织由8~9层细胞组成,基本组织细胞的细胞壁厚,细胞小且数量多(见图1-14).叶柄中维管束木质部导管分子数量多且排列紧密,叶柄中的异形维管束大小参差不等(见图1-12).导管分子形成了数量较多的管孔链,平均长度为150.041 μm,管孔团较多(见图1-14),叶柄中部的基本组织呈心形排列,其周围被木质部的导管分子等包围(见图1-12).薄壁细胞体积变小且数量增多.阳生胡桃楸韧皮部发达,平均厚度为146.736 μm,韧皮纤维木质化显著,其平均厚度为57.513 μm(见图1-14).

1 阴生环境胡桃楸叶的单生毛(×10)

3 阴生胡桃楸单细胞表皮毛、薄壁细胞、表皮细胞、角质层(×20)
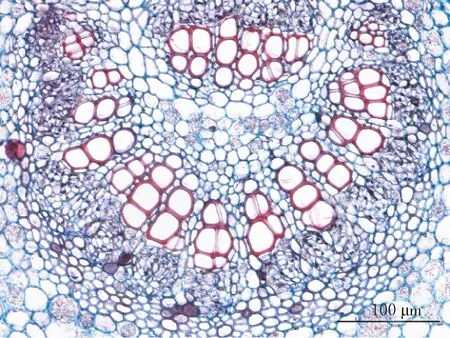
5 阴生胡桃楸主叶脉维管束、薄壁细胞、晶体(×20)

7 阴生胡桃楸栅栏组织、海绵组织、晶体(×40)
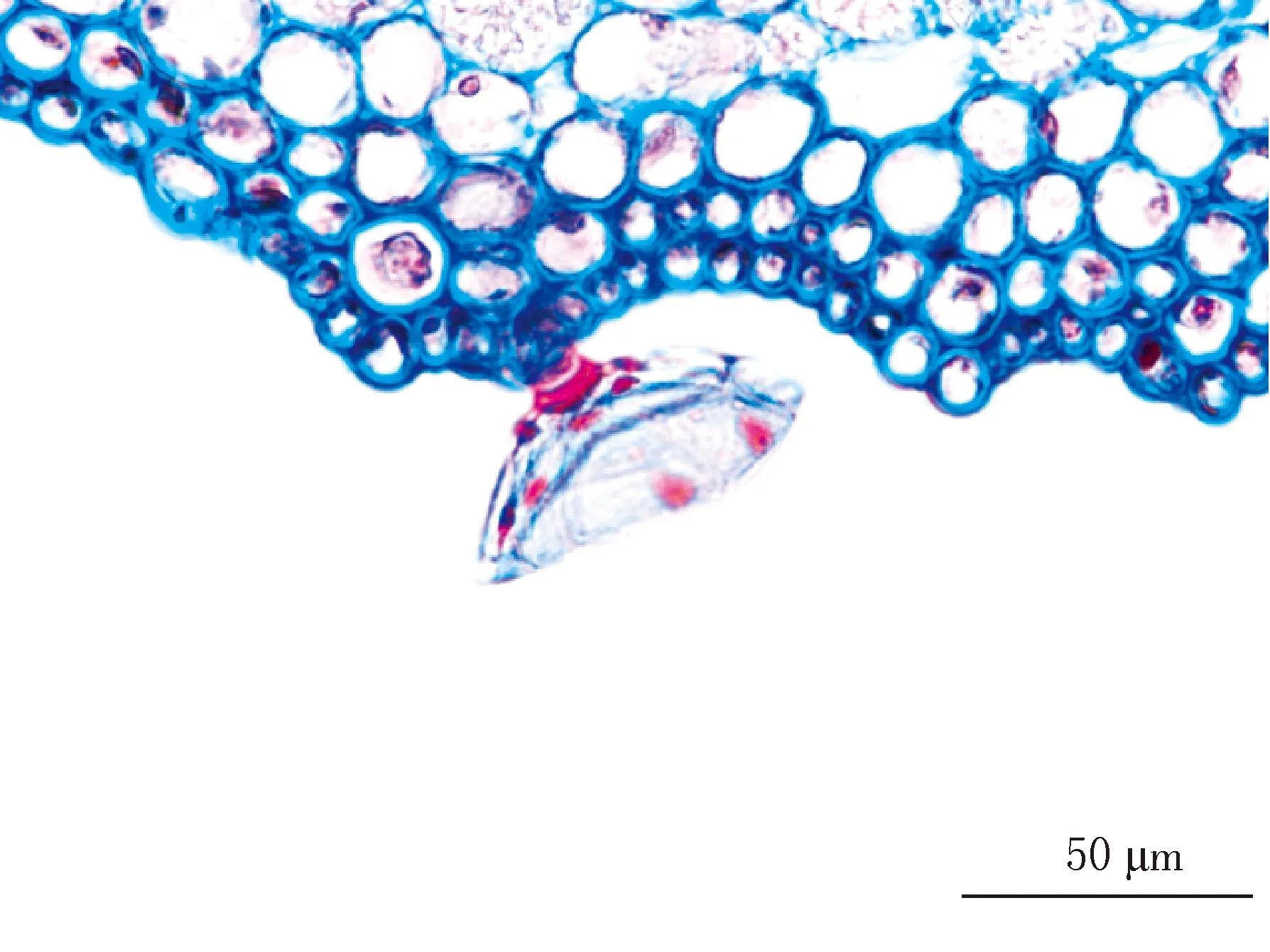
9 阴生胡桃楸腺毛、表皮细胞(×40)
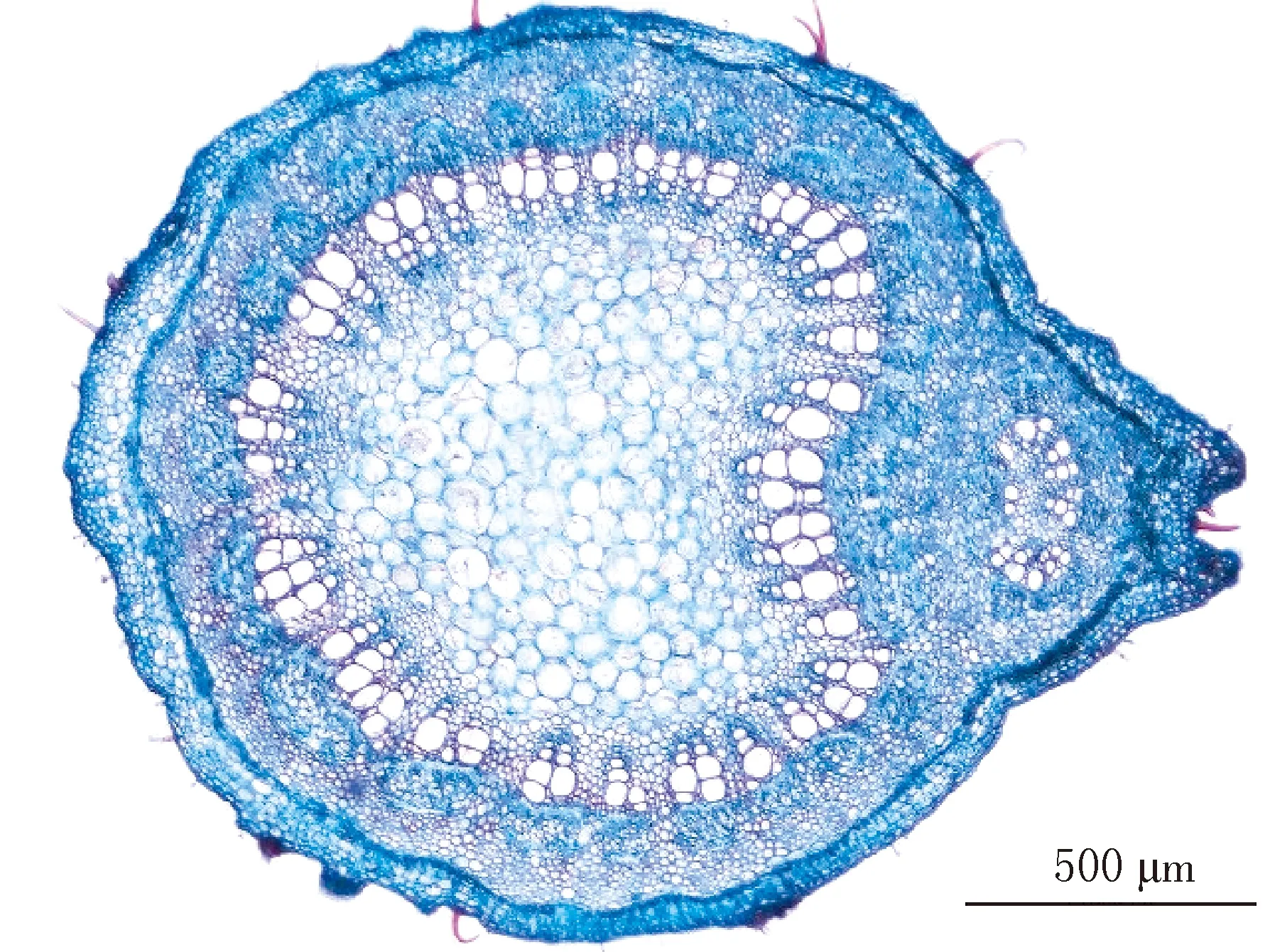
11 阴生胡桃楸叶柄的解剖结构(×4)

13 阴生胡桃楸表皮细胞、基本组织、维管束(×10)
3 讨论
本文的实验结果表明,阳生胡桃楸和阴生胡桃楸叶与叶柄的基本结构组成仍保持一致,代表了种的基本特征不变,即:稳定性和保守性.但与阳生胡桃楸相比,阴生胡桃楸颉颃弱光逆境胁迫后其叶和叶柄演化出了明显的差异.阴生胡桃楸叶片表皮细胞外切向壁角质层较薄,表面簇生毛数量显著减少,利于降低植物叶片因反射和遮挡而造成的光能损耗,从而增加植物叶片对低光量子的有效吸收率.阴生胡桃楸叶肉细胞的胞间隙和叶表皮细胞相对较发达,海绵组织较疏松,增加了植物叶片对低光量子的透过率从而促进了植物的光合作用,有助于植物抵御弱光胁迫.阳生和阴生胡桃楸叶片栅栏组织间隙中均出现了不规则的异形细胞,其内布满了大量的晶体结构,具有解毒作用,增强了胡桃楸的抗逆能力.晶体的化学特性尚待进一步的鉴定、研究.
阴生胡桃楸叶柄基本组织细胞较大,细胞间的接壤表面积明显小于阳生的,因此阴生环境的胡桃楸叶柄细胞之间的物质交流功能下降,因为细胞体积小数量多其表面积就会变大.其异形维管束数量少是颉颃阴湿环境的演化结构[13].阳生胡桃楸叶柄中演化出较多的管孔团,代表了其较为进化的特征.
阴生胡桃楸的叶片面积大、薄,颜色呈深绿色,为典型的阴生植物特征,是植物颉颃弱光逆境的演化结果:叶面积大有利于捕捉弥散光,叶片薄有利于在弱光下进行光合作用,叶片颜色深绿是因为叶绿体的长轴面与叶表面呈平行分布的结果.这也是阴生环境生长的胡桃楸为维持其正常的生理代谢需求,生理活动的活跃程度保持在相对较低水平的特殊形态和结构.阴生胡桃楸另一个颉颃逆境的演化结构为木质部导管分子直径变大,这是因为在阴生环境中水分充足,进而增强了导管输导水分和溶解在水中的无机盐的功能,是用进废退的重要结构演化特征.
从图1-3,1-4,1-11,1-12的对比可以看出,阳生胡桃楸的导管分子直径明显小于阴生环境的.由于阳生环境相对干燥缺水,多且直径小的导管分子,扩大了导管分子纵向和横向的输导面积,增强了输导水分的功能,利于颉颃干旱逆境.
数理统计分析可以看出,二者的叶片、栅栏组织、海绵组织、叶韧皮部、叶韧皮纤维、叶柄韧皮纤维厚度、叶柄木质部管孔链长度及叶与叶柄表皮细胞直径等特征参数均存在极显著差异(P<0.01),而叶主脉厚度、叶主脉木质部管孔链长度及叶柄韧皮部厚度等特征参数间差异显著(P<0.05)(见表1、表2).
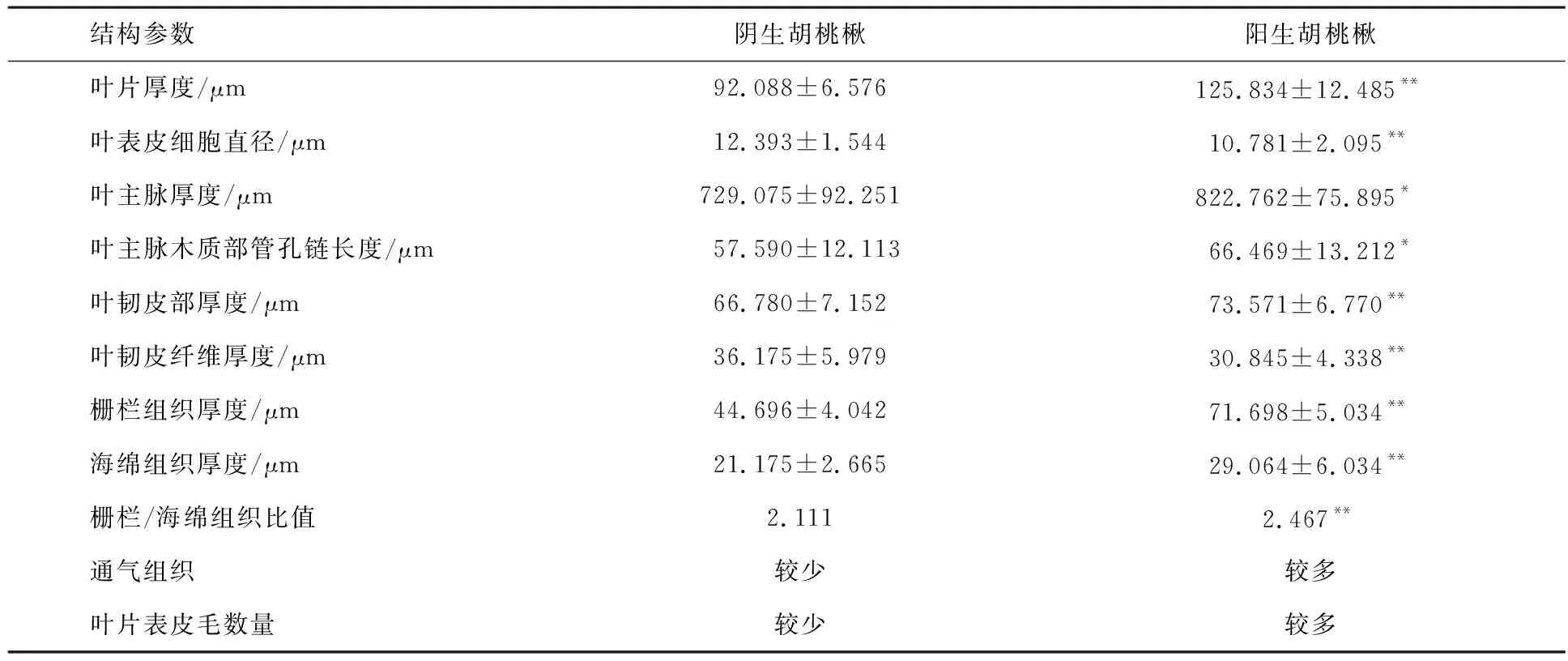
表1 不同生态环境胡桃楸叶片解剖结构参数
注:表中数值为平均值±标准差(n=20);*P<0.05,**P<0.01,下同.

表2 不同生态环境胡桃楸叶柄解剖结构参数
综上所述,本文的实验结果证明了不同生态环境的胡桃楸适应各自逆境演化出来的形态、结构和功能之间的辩证统一;同时,在一定程度上阐明了阴生胡桃楸叶和阳生胡桃楸颉颃逆境的结构演化规律.
[参 考 文 献]
[1] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1979:32-33.
[2] 宁宇,蔺立杰,王丰俊.超声波辅助提取核桃楸油及其脂肪酸分析[J].食品科技,2014(10):184-188.
[3] 王瑞,于洪侠,祁永会.胡桃楸加工和利用的木材指标测试与评定[J].现代农业研究,2016(2):34-35.
[4] YANG B Y,JIANG Y Q,MENG Y,et al.Studies on chemical constituents inn-butanol extracts from epicarp of green fruit ofJuglansmandshurica[J].Chinese Traditional & Herbal Drugs,2015,46(4):481-485.
[5] 崔玉强,张咏莉,林桂芳,等.核桃楸树皮化学成分及药物活性研究进展[J].海峡药学,2010,22(1):3-6.
[6] YAO D L,ZHANG C H,LUO J,et al.Chemical constituents from the leaves ofJuglansmandshurica[J].Chemistry of Natural Compounds,2016,52(1):93-95.
[7] 隋元杰,宋卫东,杨辉,等.加强保护与培育珍贵树种资源的建议[J].吉林林业科技,2009,38(1):59-61.
[8] 黄桂龙,毛立仁,刘玉凤,等.核桃楸的经济价值及栽培技术[J].辽宁林业科技,2010(3):52-53.
[9] 孙墨珑.核桃楸化学成分及生物活性研究进展[J].东北林业大学学报,2004,32(3):85-86.
[10] 谷安根,陆静梅.维管植物演化形态学[M].长春:吉林科学技术出版社,1993,182-186.
[11] 王勋陵,王静.植物形态结构与环境[M].兰州:兰州大学出版社,1989:99-110,154-165.
[12] 张力.SPSS19.0在生物统计中的应用[M].厦门:厦门大学出版社,2013.
[13] 胡正海.植物异常结构解剖学[M].北京:高等教育出版社,1993:124-125.
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31 06:38:50
小天使·四年级语数英综合(2022年6期)2022-06-21 12:49:36
建材发展导向(2021年14期)2021-08-23 00:56:24
作文小学中年级(2021年4期)2021-05-07 01:17:50
疯狂英语·初中天地(2019年10期)2019-10-23 05:25:20
祝您健康·文摘版(2019年4期)2019-06-11 10:20:34
学与玩(2017年4期)2017-02-16 07:05:44
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:54
小学教学研究(2016年36期)2016-05-17 05:51:58
中国房地产业(2016年9期)2016-03-01 01:26:33
