
山西历山连香树种群动态研究
2018-06-08 02:43:24张志娟毕润成
山西农业大学学报(自然科学版) 2018年6期
张志娟,毕润成
(山西师范大学 生命科学学院,山西 临汾 041000)
种群年龄结构可以反映植物的生物学特性与环境因素之间的相互关系,既可以体现濒危植物的生存现状,又能体现植物种群的演变历程及未来的发展方向[1]。而静态生命表能展现种群的年龄结构现状和发展趋势[2], 还可了解濒危种群与环境抗争的关系, 对保护和管理濒危物种具有重要的理论指导意义[3]。研究连香树种群的动态, 可以预测种群的未来发展趋势,可对保护连香树提供一定的理论依据。
连香树(Cercidiphyllumjaponicum) 为单科单属稀有种,别名山白果、五君树、紫荆叶木,系连香树科,连香树属的落叶大乔木。连香树被列为国家二级保护植物,连香树有较高的经济价值,在化工、食品、医药、园林、林业等方面有重要的作用;具有很高的科研价值,对于研究第三纪植物区系起源以及中国与日本植物区系关系等具有重要的研究价值[4],目前连香树零星的分布于我国的皖、湘、鄂、川、豫、晋、陕、甘、赣、浙等省以及日本的本州、九州、四国,其多分布于海拔在600~2 000 m的沟谷两侧地区,种群数量与规模已相当有限,甚至有的呈单株生长[5], 当前国内外研究学者对于连香树的研究工作涉及到植物区系特征[6]、资源现状调查[7~9]、种子萌发[10]、传粉生物学[11]等方面,对山西历山自然保护区地区连香树种群动态方面的研究比较少,通过比较分析山西历山连香树集中分布的两种不同生存坏境条件下的种群动态,旨在阐明其种群的未来发展趋势,探讨其濒危的主要原因,进而为山西历山地区连香树的保护提供一定的科学依据。
1 研究区概况
历山国家级自然保护区位于山西省南部中条山山脉的东段(35°16'30″N—35°27'20″N,111°51'10″ E—112°5'35″E),是运城、晋城、临汾三市的翼城、沁水、阳城、垣曲四县的交界地带。自然保护区总面积24 200 hm2,森林覆盖率为80.9%,最高海拔(舜王坪) 2 358 m,相对高差 2 058 m。该地区属于暖温带大陆性季风气候,年均温度8~12 ℃,年均相对湿度 70 %,年降水量 600~800 mm,一般集中在7-8月,无霜期 150~180 d。地带性土壤为褐土,土壤类型随海拔、地形、坡度和植被类型的变化而变化,由山顶到山基依次为山地草甸土、棕色森林土、山地淋溶褐土和山地褐土[12]。
2 研究方法
2.1 野外调查
在2016年8月,在连香树分布集中的地区沁水县和垣曲县设置样方,根据目标物种连香树,共建立了24个20 m×20 m 样方进行统计, 把每个样方划分成2个10 m×10 m的小样方,在小样方的对角线设置2个5 m×5 m 灌木样方和4个1 m×1 m的草本样方,乔木层记录其高度, 胸径(乔木层的起测胸径是5 cm, 胸径小于5 cm的乔木幼苗归到灌木层)、冠幅、枝下高等指标;灌木层记录种名、高度、盖度等;草本层记录种名、高度、盖度等。同时记录生长地区的相关环境因子,包括海拔、坡度、坡向等(表1)
2.2 年龄结构
采用“空间代替时间”的方法,并根据连香树生物学特性,将其胸径分为 12个径级,采用表2所建立龄级分类标准,分析其年龄结构。
2.3 生命表
本研究采用“空间代替时间”的方法,把树木的径级从小到大的顺序看作时间顺序,统计各龄级的株数[13],编制山西历山地区连香树种群的生命表,并对其进行分析。
2.4 时间序列预测模型
采用以下模型对种群数量动态进行预测[14]:
式中,Mt:在未来的n年时,t龄级种群的存活数;n:预测时间;t:龄级:Xk:当前k龄级的种群存活数。
3 结果与分析
3.1 连香树年龄结构分析
种群的年龄结构可以反映种群大小个体的组配状况,也可以解释种群的生存状况和繁殖策略[14],由图1可以看出,在沁水县,第III、IV径级(14 表1历山连香树种群基本情况 调查区Survey region样方号No.of samples海拔/mAltitude坡向Aspect坡度/°Slope沁水县垣曲县11210.3111.1°SW<521218.990.8°SW<531220.822.8°SW⫺541339.880.3°SW1051430.330.2°SE1461434.9181.4°SW1871451.6280.3°SW2081460.533.2°SW2291468.310.8°SE23101466.214.8°SW23.811595.111.2°SE⫺521598.820.3°SE⫺1031598.348.8°SW1541599.491.0°SW1651610.1190.1°SE1861614.350.1°SW2271615.192.2°SW2381618.981.0°SE2491622.228.8°SW22101632.88.3°SW10111632.032.8°SW28121633.818.1°SW28131638.8100.8°SW14141639.1100.8°SW30 根据生命表的数值能了解该种群的现存状态,分析该种群的结构及受干扰程度,预测该物种未来的数量动态[15],由表3可知,沁水县中第I、第II径级和垣曲县中第III、第IV径级死亡率qx为负值,这是由于种群中幼苗和幼树少而导致的。其中,沁水县种群中第VIII、第XII径级的死亡率较高,垣曲县中第VII、第径级XI的死亡率较高。这表明两种群随着个体的生长,种群生存环境的变化,各种群中的个体对光照,水分等资源的需求量增大,种内、种间竞争加剧,且连香树多沿沟谷生长,其本身对坏境条件的要求苛刻,导致种群死亡率升高,而后个体数量减少到一定程度后,竞争减少,使死亡率保持到相对稳定状态。 表2历山地区连香树种群龄级表 龄级Age class高度/cmHeight胸径/cmDiameter at breast height生长期Period of growthIH≤50DBH⫹4幼苗期II50 图1 历山地区连香树种群径级结构Fig.1 Size structure of Cercidiphyllum japonicum in Lishan 由表3可知,两地的消失率Kx和死亡率qx的变化趋势基本一致,波动较大,消失率和死亡率先升高,后下降,而后又升高,由于连香树的幼苗繁殖率低、成活率低,但随着种群的生长,生存下来的个体抵抗恶劣环境的能力增强,种群的消失率有所下降,而后又随着种群的老化、种内种间的竞争、生态环境的变化、人类活动的干扰等,使得消失率再次升高。 表3 历山地区连香树种群生命表Table 3 Life table of Cercidiphyllum japonicum population in Lishan 注:ax:存活数;lx:存活量;dx:死亡量;qx:死亡率;Lx:区间寿命;Tx:总寿命;ex:期望寿命;Kx:消失率;Sx:存活率。 由表3可知,在沁水县,第V径级的生命期望值ex较高;在垣曲县,第II径级的命期望值较高;总体上,两地的幼苗期及幼树期的平均生命期望值较高。这表明,在沁水的第V径级和垣曲的第II径级的生命质量较高,两地的连香树种群的幼苗和幼树有较高的生命期望值,随着个体的生长,中老龄期生命期望值略有波动,但总体上呈逐渐降低趋势。 存活曲线能反映生存率及死亡率随年龄变化的情况,对反映种群动态有着重要的意义[16]。以连香树径级为横坐标,标准化存活数为纵坐标分别绘制垣曲县和沁水县连香树种群的存活曲线。由图2可知,两个种群存活曲线基本呈 DeeveyⅢ型。即幼苗死亡率高,种群数量急剧减少,随着年龄的增加,死亡率逐渐下降,种群规模开始稳定,达到生理年龄后,死亡率开始上升。连香树种群幼年个体的种内竞争激烈,造成很高的死亡率,经过环境筛的选后,死亡率降低,种群实现了定居,直至其生理衰退年龄,死亡率升高。在沁水县和垣曲县,连香树种群都呈现出中老龄占据大多数,而幼龄占据少数的状态。整体上,种群的个体数量少,呈衰退趋势,和年龄结构的分析结果相符。 图2 历山地区连香树种群存活曲线Fig.2 Survival curve of Cercidiphyllum japonicum population in Lishan 由图3可知,横向来看,沁水县和垣曲县连香树种群,各径级种群数量有的增加,有的减少,在未来10年、20年,将呈现出随着种群的生长,径级的增大,植株数量先增多,而后又减小的趋势。在预测序列中,两种群在X径期均出现数量峰值;纵向来看,随着时间的变化,幼苗和幼树急剧减少,老龄植株增多,两种群的个体数量都将减少,种群衰退加剧。纵横向分析都验证了种群的衰退趋势。由此可知,如果不采取人工措施增加种群的幼苗和幼树,单靠连香树天然种群的自我繁殖,在10年、20年后,随着生态环境的变化、人类的干扰,连香树现有个体将逐渐减少,甚至消失。 图3 历山地区连香树种群数量动态预测Fig.3 The prediction of Cercidiphyllum japonicum dynamics of population in Lishan 年龄结构可体现濒危植物与其生境间的关系,也可体现其生态位的大小;生命表可以体现濒危种群过去和未来的数量存亡动态[17]。两地连香树的种群年龄结构都呈现衰退趋势,且两种群年龄结构都出现了断续。种群衰退表明连香树的生存环境在恶化。种群衰退还表明,连香树在激烈的种内及种间的竞争中都处于弱势状态。年龄结构出现断续表明种群的衰退程度较大,种群消失的可能性大,亟需保护。在生命表中出现了死亡率qx为负值,虽然与数学假设不相符,但能表明种群是在不断变化的,是处在迅速的发展或衰退之中并非静止不动的,仍有重要的生态学记录意义[18]。沁水县中第I、第II径级和垣曲县中第III、第IV径级的死亡率出现了负值, 表明两种群幼苗和幼树少,种群在迅速衰退。 种群的存活曲线能反映出种群的动态趋势,可以体现出种群在生长发育过程中各个龄级个体的死亡率变化[19]。 研究结果表明,两种群的存活曲线呈现DeeveyⅢ型,种群个体数目较少,连香树多沿沟谷两侧生长,本身对生存坏境的要求苛刻,导致种群生长地狭小,种群数量有限。这和野外实地调查的连香树实际情况相符。种群时间序列的分析能适用于多种复杂条件下的种群数量动态预测,且预测准确性高。研究结果表明,两种群随着时间的的变化,幼苗急剧减少,老化植株增多,种群个体数目都将减少。其主要原因是幼苗繁殖不足,且成活率低。幼苗繁殖不足的原因一方面是因连香树以小种子进行繁殖[20],对生存坏境的要求较高,且结实率低,成活率低;另一方面是人类活动对种子萌发及生长的干扰。能够为濒危植物种子的萌发、幼苗的定居及生长发育创造适宜环境条件,是保证种群稳定的关键性因素[21],因此,若幼苗繁殖不足,加之人类活动对种子萌发及幼苗的生长环境的干扰,将加快种群衰退甚至消失。种群的年龄结构、静态生命表、存活曲线以及种群数量动态预测都一致表明,山西历山地区连香树种群呈衰退趋势,幼苗不足,其繁殖特性及其对生境苛刻要求是导致连香树濒危的主要原因。 针对连香树濒危的原因,对山西历山地区连香树的保护工作,对垣曲县来说,测重点是要尽可能的采取人工措施,通过嫁接、植物组织培养等技术手段,建立幼苗繁殖基地,繁殖和培育连香树幼苗;而对沁水县来说,因近年来旅游业的快速发展,且连香树刚好处在旅游景点附近,受人类活动干扰强,除了采取人工措施, 增加连香树幼苗外,还要重点采取措施限制人类活动影响,可通过建立连香树保护区,减少人类活动对幼苗萌发、生长的的影响以及对连香树原始生存环境的破坏。 参 考 文 献 [1]李文良,张小平,郝朝运,等.湘鄂皖连香树种群的年龄结构和点格局分析[J].生态学报, 2009,29(6): 459-468. [2]Harcombe PA. Tree life tablest[J]. Bioscience, 1987,37(8): 557-568. [3]吴承祯,洪伟,谢金寿,等.珍稀濒危植物长苞铁杉种群生命表分析[J].应用生态学报, 2000,11(8):333-336. [4]易雪梅,张悦,王远遐,等.长白山水曲柳种群动态[J].生态学报, 2015,35(1):91-97. [5]吴甘霖.连香树的生物学特性及其资源保护[J].安庆师范学院学报(自然科学版), 2006,12(2): 106-109. [6]黄绍辉,方炎明.安徽歙县连香树群落的植物区系特征分析[J].徐州工程学院学报, 2007,22(10):44-45. [7]付素静,高宇琼,龙梅珍,等.珍稀树种连香树资源现状与园林应用分析及建议[J].现代园艺, 2012(1): 28-29. [8]刘胜祥,黎维平,杨福生,等.神农架国家级自然保护区连香树资源现状及其保护[J].植物资源与环境, 1999,8(1):33-37. [9]程丽芬.历山自然保护区连香树资源调查[J].山西林业科技, 2014,43(3):43-44. [10]杨欣超,王瑞辉,曹基武,等.湖南省野生连香树生境调查及濒危原因分析[J].湖南林业科技, 2012,39(2):38-41. [11]熊丹,陈发菊,梁宏伟,等.珍稀濒危植物连香树种子萌发的研究[J].福建林业科技, 2007,34(1):36-39. [12]袁丽洁,方向民,崔波,等.濒危植物连香树的传粉生物学研究[J].河南农业大学学报, 2007,41(6):647-650,654. [13]铁军,李燕芬,王霞,等.山西历山国家级自然保护区猕猴栖息地森林群落物种多样性[J].生态学杂志,2015,34(11):3009-3015. [14]张志祥,刘鹏,蔡妙珍,等.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1146-1156. [15]郭微,上官铁梁,王志明,等.灵空山油松种群年龄结构与动态分析[J].植物科学学报,2013,31(2):130-135. [16]张志祥,刘鹏,蔡妙珍,等.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1146-1156. [17]肖宜安,何平,李晓红,等.濒危植物长柄双花木自然种群数量动态[J].植物生态学报2004,28(2):252-257. [18]张文辉,许晓波,周建云,等.濒危植物秦岭冷杉种群数量动态[J].应用生态学报2005,16(10):1799-1804. [19]于大炮,周莉,董百丽,等.长白山北坡岳桦种群结构及动态分析[J].生态学杂志,2004,23(5):30-34. [20]张文辉,祖元刚,刘国彬.十种濒危植物的种群生态学特征及致危因素分析[J].生态学报, 2002,22(9): 1512-1520. [21]安静,吴玲,王海娟,等.不同干扰程度下沙生植物异翅独尾草的种群结构与动态特征[J].生态学报, 2017,37(6):2023-2032.
Table1 The basic status ofCercidiphyllumjaponicumpopulation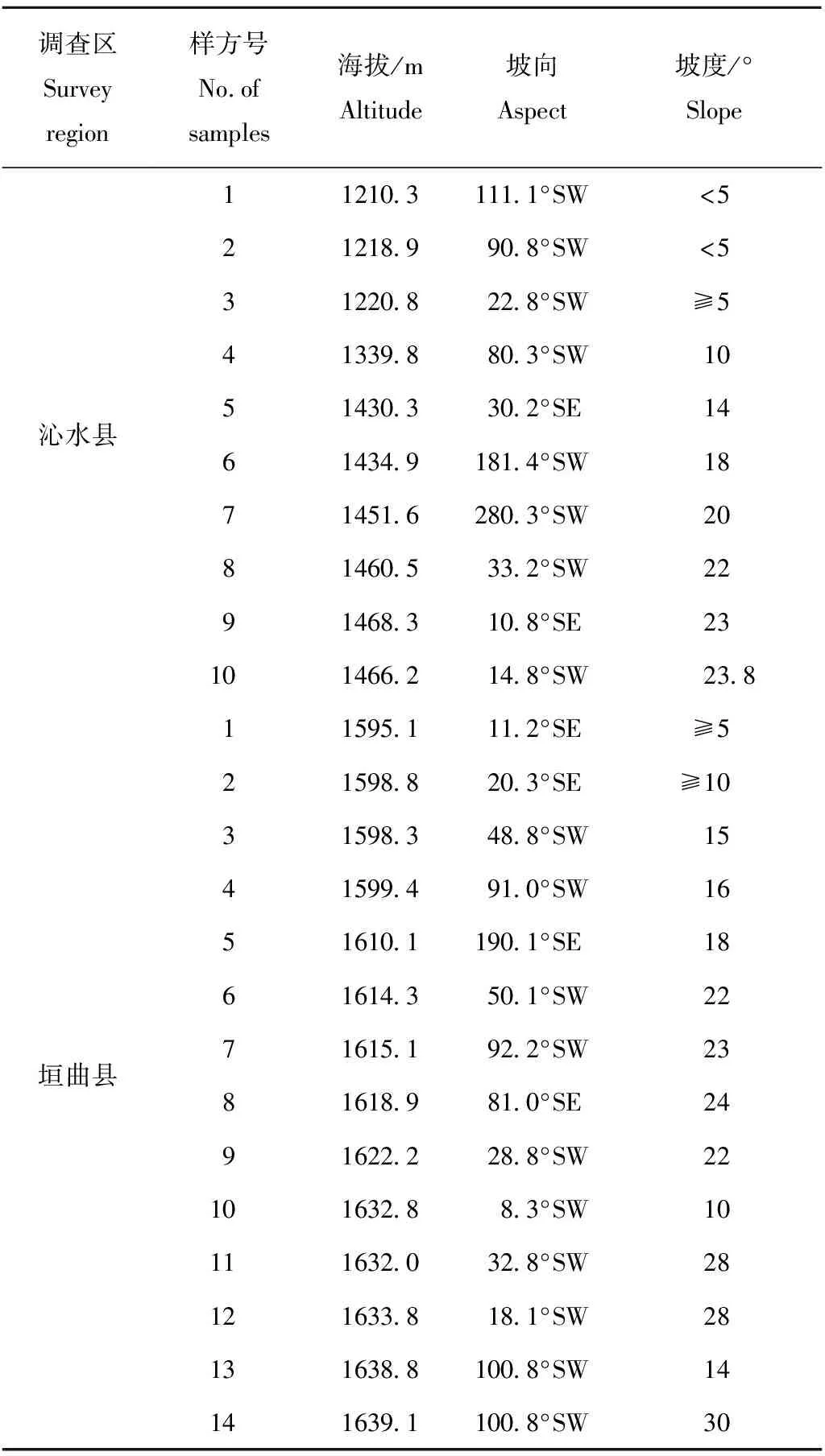
3.2 种群静态生命表
Table2 Age class ofCercidiphyllumjaponicumpopulation in Lishan region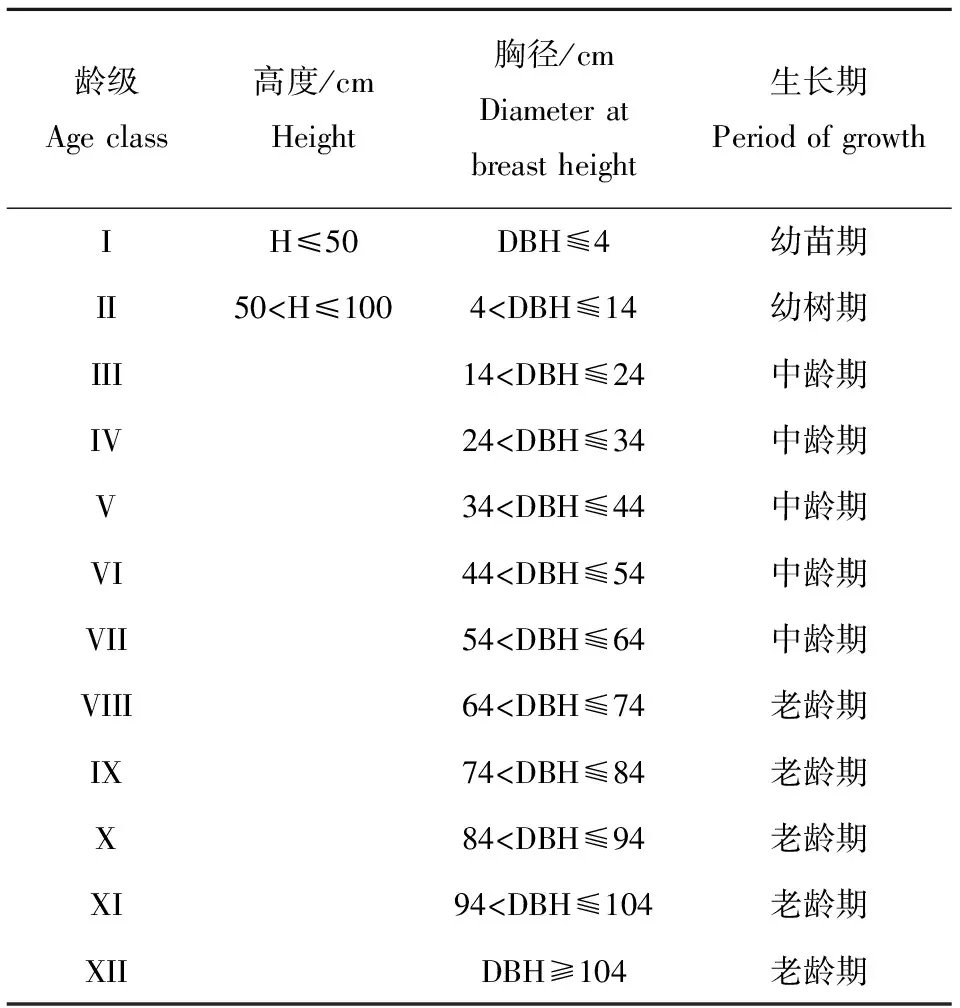

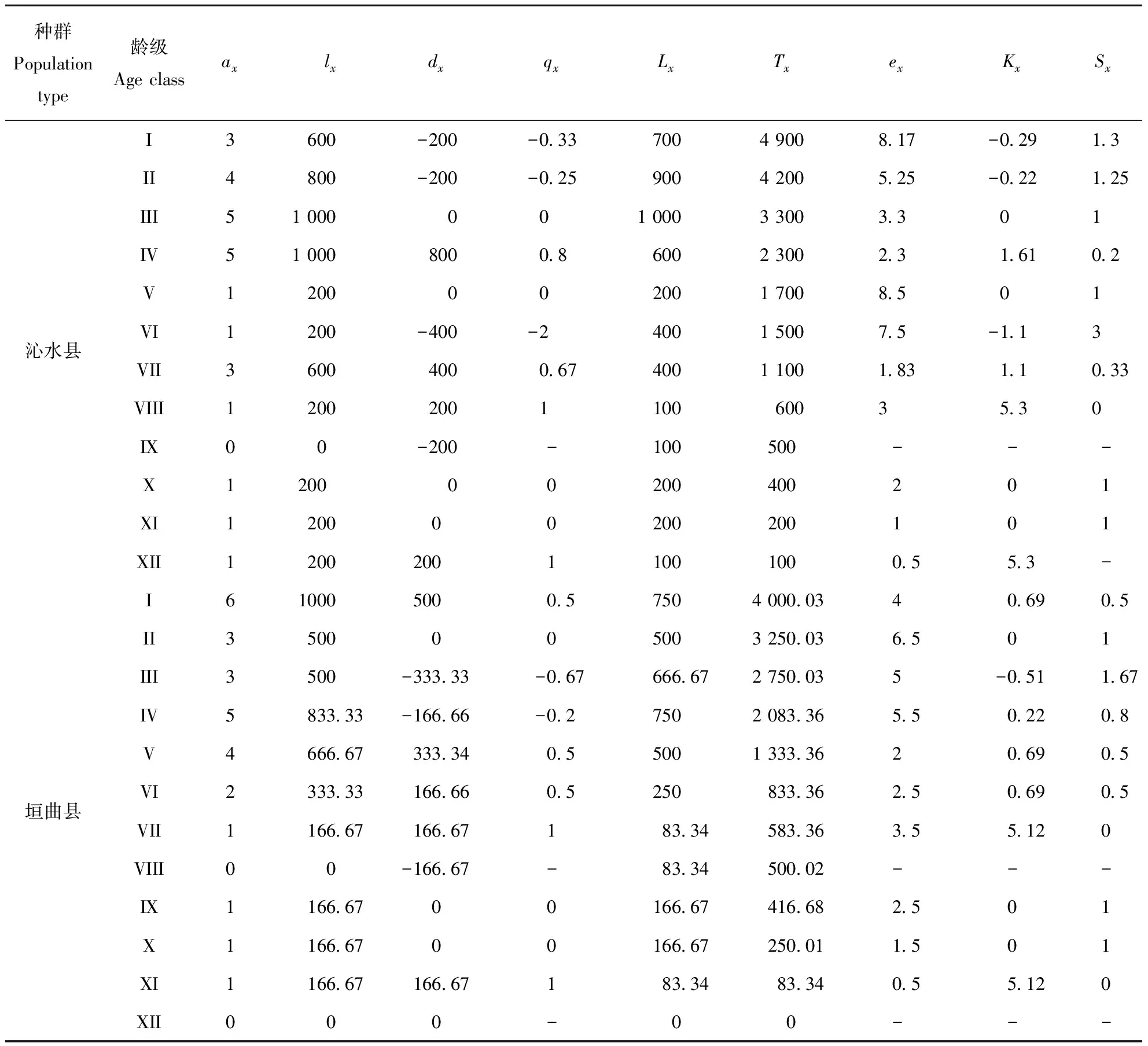
Notes:ax: Survival number;lx: Survival quantity;dx:Death number;qx:Mortality rate;Lx:Span life;Tx:Total life;ex:Life expectancy;Kx:Vanish rate;Sx:Survival rate.3.3 种群的存活曲线
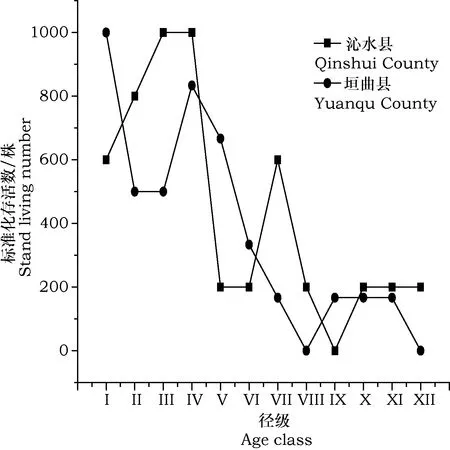
3.4 时间序列预测

4 讨论
猜你喜欢
今日农业(2022年13期)2022-09-15 01:19:16今日农业(2021年19期)2021-11-27 00:45:49今日农业(2020年22期)2020-12-25 02:30:40中华戏曲(2020年2期)2020-02-12 05:18:12中华戏曲(2020年2期)2020-02-12 05:17:58青年生活(2019年32期)2019-09-10 04:25:38小火炬·阅读作文(2019年4期)2019-08-06 04:33:28支部建设(2019年36期)2019-02-20 13:21:22今日农业(2018年14期)2018-08-11 06:15:12畅谈(2018年24期)2018-02-02 02:31:16
