
CO2浓度升高与增温对半干旱区马铃薯光合特性的协同影响
2018-06-07 09:32姚玉璧雷俊牛海洋张秀云肖国举
生态环境学报 2018年5期
姚玉璧 ,雷俊*,牛海洋,张秀云,肖国举
1. 中国气象局兰州干旱气象研究所/甘肃省干旱气候变化与减灾重点实验室/中国气象局干旱气候变化与减灾重点开放实验室,甘肃 兰州 730020;2. 甘肃省定西市气象局,甘肃 定西 743000;3. 宁夏大学,宁夏 银川 750021
政府间气候变化专门委员会(IPCC)第五次评估报告(AR5)认为,近百年来全球气候变暖毋庸置疑。1880—2012年,全球平均地表温度升髙了0.85 ℃;1951—2012年,全球平均地表温度的升温速率为 0.12 ℃·(10a)-1,几乎是 1880年以来的两倍;1983—2012年是自1850年以来最暖的3个10年,气候变暖的趋势特征几乎在全球各地均可以观测到(IPCC,2013)。1951—2016年,中国地表年平均气温呈显著上升趋势,增温速率为0.23 ℃·(10a)-1(中国气象局气候变化中心,2017)。
预计 2016—2035年全球平均地表温度将继续升高0.3~0.7 ℃。由于人类活动加剧,自1750年以来,大气中CO2浓度不断增加,到2011年达到391 μmol·mol-1。按照典型浓度目标中低排放情景(RCP4.5),辐射强迫稳定在4.5 W·m-2,2100年后CO2当量浓度稳定在约 650 μmol·mol-1(IPCC,2013)。
温度改变导致植物叶片气孔导度、蒸腾速率和土壤蒸发速率发生变化,进而作用于作物水分循环和蒸散发过程(Rawson,1988;Zhou et al.,2011)。模拟大气增温的红外线辐射器田间增温试验表明,增温使马铃薯(Solanumtuberosum L.)生理生态和产量形成过程均发生了显著变化(Xiao et al.,2013a;Xiao et al.,2013b)。春秋季气温增高有利于马铃薯生长发育及产量形成,而夏季气温增高导致马铃薯生育更加脆弱(姚玉璧等,2013;赵年武等,2015)。
增加CO2浓度对马铃薯总生物量和产量有积极的作用,在CO2浓度升高的情况下,马铃薯总生物量、产量和水分利用效率(WUE)均有所提高(Schapendonk et al.,2000;Sicher et al.,1999;Wheeler et al.,1999),当 CO2浓度高于环境 370~740 μmol·mol-1,则马铃薯块茎产量增加 27%~49%(Wheeler et al.,1991);然而,在试验中也观测到CO2浓度倍增对产量有轻度的负效应(Goudriaan et al.,1983);提高 CO2供应可以加速叶片衰老,加快开花期进程(Lawson et al.,2001)。增加CO2浓度减少了马铃薯叶片的蒸腾速率,提高了光合作用和水分利用效率(Ku et al.,1977),当开顶气室中增加 CO2浓度 350~700 μmol·mol-1,冠层光合作用提高 80%,但不同的生长季处理的结果是不同的(Schapendonk et al.,2000)。相对于对照而言,提高CO2浓度处理,马铃薯生长季叶片净光合速率增加(Sicher et al.,1999)。
研究大气增温、CO2浓度升高与植物光合生理生态的关系一直是植物生理和植物生态学研究的热点问题(Moreno et al.,2002;Awada et al.,2003;Damesin,2003)。植物光响应过程及其模拟研究可以明确净光合速率与光合有效辐射通量密度之间的定量关系,揭示植物光化学过程效率(Robert et al.,1984)。利用光响应曲线可以估算光补偿点、光饱和点、表观量子效率、最大净光合速率和暗呼吸速率等重要的光合参数(闫小红等,2013;叶子飘,2010)。国际上常用的光合光响应曲线模型有直角双曲线模型(Baly,1935;Miko et al.,1987)、非直角双曲线模型(Thornley,1976,1998)、指数函数(Prado et al.,1997)和直角双曲线的修正模型(叶子飘,2007;Ye,2007;Ye et al.,2012)等。光响应曲线的模型的选择与模拟决定着植物叶片光合能力重要参数的确定。
全球有157个国家种植马铃薯,2014年全球马铃薯种植面积 1920.5×104hm2,总产量达3.85×108t;中国马铃薯种植面积达 681.7×104hm2,产量1.27×108t,占全球产量的25.9%。马铃薯已成为继水稻、小麦、玉米之后的第四大主粮作物,是最有发展前景的高产作物之一(李子涵等,2016)。马铃薯耐旱、耐贫瘠,是中纬度半干旱区适宜种植的特色优势高产作物(张强等,2012;赵鸿等,2013)。马铃薯生长发育及其产量形成受气候变暖的影响也十分突出,大气增温和 CO2浓度倍增对中纬度半干旱区马铃薯光合特性研究尚缺乏试验基础和深入探讨。为此,开展 CO2浓度升高与大气增温对马铃薯光合生理生态特性的协同影响研究,分析马铃薯叶片光响应过程特征,建立马铃薯叶片光响应曲线模型,可为马铃薯生理生态模拟提供科学基础,为马铃薯栽培适应气候变化提供科学依据。
1 试验设计与方法
1.1 研究区域气候及马铃薯生长发育概况
1981—2010年,研究区域年平均气温为7.2 ℃,最热月7月平均气温为19.2 ℃,最冷月1月平均气温为-7.2 ℃。年平均降水量为377.1 mm,5—10月降水量为 328.5 mm,占全年降水量的87.0%。年平均日照时数为 2437.5 h,年平均连续无霜期为145 d。
试验区域马铃薯一般在5月上旬—5月中旬播种,出苗期在6月上旬—6月中旬,分枝期在6月下旬—7月上旬,花序形成期在7月上旬—7月中旬,开花期在7月中旬—7月下旬,可收期在10月上旬—10月中旬。播种—可收期全生育期有135~165 d。马铃薯全生育期≥0 ℃积温为2500~3000 ℃,降水量为300~350 mm,日照时数为 1100~1400 h。
1.2 试验设计
1.2.1 试验设计
试验于2016年4月—2016年10月在中国气象局定西干旱与生态环境野外试验基地内实施,进行CO2浓度升高与大气增温对马铃薯生理生态的协同影响试验。试验利用新型开顶式气室OTC(Open-top chamber),其为八面柱状透光玻璃结构,顶部向内斜伸20 cm,底面积18 m2,主体高3 m,顶部开口,因其更适宜于CO2浓度控制而优于FACE(Free-Air CO2Enrichment),是目前国际上认可的CO2倍增模拟试验设备。
根据“典型浓度目标”(Representative Concentration Pathways,RCPs)情景,假设人类努力减少温室气体排放的RCP4.5情景,辐射强迫稳定在4.5 W·m-2,21世纪末CO2当量浓度稳定在650 μmol·mol-1,全球地表温度升高不超过 2.0 ℃(IPCC,2013)。
试验设计2个处理及对照:一是增温处理(IT),采用温度监测仪,控制空气温度升高(2.0±0.5) ℃;二是增温(IT)+CO2浓度升高(IC)复合处理(IT+IC),采用CO2浓度监测控制仪,将CO2浓度控制在(650±20) μmol·mol-1;大田对照(CK),自然空气 CO2浓度约 370 μmol·mol-1。每个处理设置 3个重复。
以本地大面积栽培品种“新大坪”为供试材料。马铃薯播种时间为2016年4月29日,收获期为10月15日,行距40 cm,株距45 cm。
在马铃薯出苗—可收期全生育期利用CO2浓度监测控制装置进行熏气,熏气时间为每天 07:00—19:00,整个试验期间,气室内CO2浓度保持稳定。
试验期间水分、肥料均匀一致,无病虫害及杂草等限制因素。
1.2.2 测定项目和方法
在马铃薯分枝期、花序形成期、开花期测定马铃薯光合特征参数,观测时间设为晴天 09:30—11:30,进气速率为 500 μmol·mol-1,采用美国LI-COR公司生产的用Li-6400XT标准叶室测定马铃薯净光合速率(net photosynthetic rate,Pn)、气孔导度(conductance to H2O,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr)。光响应曲线的测定应用Li-6400-02B红蓝光源提供的不同光合有效辐射强度(photosynthetically active radiation,PAR,单位:μmol·m-2·s-1),分别在 2300、2000、1900、1700、1500、1300、1100、900、700、500、300、200、150、100、75、50、20、0 μmol·m-2·s-1共 18 个光点下测定马铃薯叶片净光合速率(μmol·m-2·s-1)。
马铃薯发育期、高度、密度、生物量、叶面积和产量的测定按照《农业气象观测规范》进行测定(国家气象局,1993)。
1.3 统计分析方法
1.3.1 气候变化特征分析
气候要素的趋势倾向率变化计算公式(魏凤英,2007)如下:

式中,Xi为气候要素变量;ti为 Xi所对应的时间;a为回归常数;b为回归系数;n为样本量。10b为要素的倾向率。
1.3.2 光响应曲线拟合
光响应曲线的拟合采用非直角双曲线模型(non-Rectangular hyperbola equation)进行拟合,其表达式为:

式中,Pn为净光合速率(μmol·m-2·s-1);I为光合有效辐射(μmol·m-2·s-1);α为植物光合作用对光响应曲线在I=0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率(μmol·m-2·s-1);Pnmax为最大净光合速率(μmol·m-2·s-1);θ 为曲线曲角(0≤θ≤1);Rd为暗呼吸速率(μmol·m-2·s-1)。
采用直线方程拟合弱光强条件下(≤200 μmol·m-2·s-1)的光响应数据得到表观量子效率(AQE,μmol·m-2·s-1),利用非线性最小二乘法估算Pnmax。光补偿点(LCP,μmol·m-2·s-1)及光饱和点(LSP,μmol·m-2·s-1)通过求解直线方程式(3)得到(叶子飘,2010):

当Pn=0时,计算得I的值为LCP;当Pn=Amax时,计算得I为LSP。
非直角双曲线模型应用 SPSS 15.0分析软件(SPSS Inc.Chicago,Ill,USA)中的最小二乘法进行拟合。
其中,大田对照(CK)光合光响应曲线完全应用非直角双曲线模型拟合;增温处理(IT)光合光响应曲线、增温(IT)+CO2浓度升高(IC)复合处理(IT+IC)光合光响应曲线采用非直角双曲线模型与Cubic函数分段拟合。数据处理及作图在Excel 2007中完成。
2 结果与分析
2.1 试验区气候变化特征
2.1.1 气温变化

图1 试验区逐年气候要素变化曲线Fig. 1 The annual climatic factors change with the experiment area
图1a所示为1958—2016年试验区逐年气温变化曲线,近 59年气温呈极显著上升趋势,气温线性回归倾向率为 0.42 ℃·(10a)-1(r=0.808,n=59,P<0.001)。20世纪60年代至80年代,气温距平值均为负距平,气温距平值60年代为-0.5 ℃,70年代为-0.4 ℃,80年代为-0.2 ℃;90年代为0.5 ℃,21世纪00年代为1.3 ℃,2011—2016年为1.7 ℃。
近59年冬季(12月至翌年2月)变暖速率最大,冬季气温倾向率为0.50℃·(10a)-1(P<0.001);秋季(9—11月)气温倾向率为 0.42 ℃·(10a)-1(P<0.001),变暖速率仅次于冬季;夏季(6—8月)气温倾向率为0.37 ℃·(10a)-1(P<0.001);春季(3—5月)变暖速率最小,其值为 0.35℃·(10a)-1(P<0.001);马铃薯生长发育阶段(5—10月)气温倾向率为 0.37 ℃·(10a)-1(P<0.001)。
2.1.2 降水量变化
1958—2016年试验区逐年降水量变化如图 1b所示,近 59年,年际降水量呈减少趋势,降水量倾向率为-12.17 mm·(10a)-1(r=0.25,n=59,P=0.05)。试验区域年际降水量气候平均值为 377.1 mm,在245.7~720.1 mm 之间振荡,降水量距平百分率为-34.8%~91.0%。20世纪60年代降水量偏多,年平均降水量447.6 mm;90年代偏少最多,年平均降水量365.7 mm;21世纪00年代降水量偏少,年平均降水量仅375.6 mm。
历年冬季降水量呈略增加变化趋势,增速为0.42 mm·(10a)-1(P>0.10),未通过显著性检验;春季和夏季降水量气候倾向率分别为-1.59 mm·(10a)-1(P>0.10)、-5.43 mm·(10a)-1(P>0.10),呈减少趋势,也未通过显著性检验。秋季降水量气候倾向率为-5.60 mm·(10a)-1(P<0.10),呈减少趋势;马铃薯生育期降水量气候倾向率为-10.12 mm·(10a)-1(P<0.10),也呈减少趋势,同时也是降水量减少最多的时段。
就降水量变化的稳定性而言,冬季降水量年际差异最大,极不稳定,变异系数达54.4%。其余季节年际变异系数在25.9%~44.8%之间。马铃薯生育期降水量年际差异最小,变异系数为20.7%。降水量年际变化较稳定时段与马铃薯生育期基本吻合,是马铃薯栽培降水量气候保证率相对高的时段。

图2 不同处理马铃薯净光合速率(a)和气孔导度(b)变化Fig. 2 Changes the net photosynthetic rate (a) and the stomatal conductance (b) of potato for different treatments
2.2 CO2浓度升高与大气增温对马铃薯光合特性的影响
2.2.1 马铃薯净光合速率变化特征
分别在马铃薯营养生长期、生殖生长期(开花期、块茎膨大期)测定马铃薯叶片净光合速率(图2a)。在营养生长期及块茎膨大前期,马铃薯叶片净光合速率表现为IT+IC>IT>CK,IT+IC处理马铃薯叶片的净光合速率在 18.6~23.8 μmol⋅m-2⋅s-1之间,IT 处理在 9.1~16 μmol⋅m-2⋅s-1之间,CK 在3.7~13.4 μmol⋅m-2⋅s-1之间,IT+IC 处理马铃薯叶片净光合速率较CK提高1~5倍,平均提高了2.1倍,IT处理较CK提高22%~140%,平均提高85%,均通过 0.01显著性检验。这表明,在马铃薯生育前期,大气温度偏低,未达到马铃薯生长发育的最适温度,增温补偿了因温度低而造成的马铃薯叶片光合速率低的状况,增温条件下马铃薯叶片的光合速率提高,在增温处理的同时,CO2浓度升高,叶片进行光合作用的原料增加,净光合速率进一步提高。因此,增温加CO2浓度升高复合处理的净光合速率远大于其他处理。
由于前期增温加 CO2浓度升高复合处理及增温处理马铃薯生长旺盛,在生育后期,植株叶片衰老加速,净光合速率的变化趋势表现为 CK>IT>IT+IC,存在显著或极显著差异,IT+IC处理较CK降低2.1~2.6倍,IT处理较CK降低1.1~1.13倍,净光合速率下降。
可见,在营养生长期,马铃薯叶片净光合速率表现为增温加 CO2浓度升高复合处理大于增温处理,且大于对照;在生育后期,由于前期增温加CO2浓度升高复合处理及增温处理马铃薯生长旺盛植株叶片衰老加速,增温加CO2浓度升高复合处理净光合速率下降更为迅速。
2.2.2 马铃薯叶片气孔导度的变化特征
马铃薯叶片气孔导度在整个生育期表现为先升后降的变化趋势(图2b)。对增温和CO2浓度升高背景下叶片气孔导度进行分析,结果表明,CO2浓度升高和大气增温处理,马铃薯叶片气孔导度较CK下降4.8%~49.0%,平均下降幅度为43.4%。在生育前期,气孔导度下降幅度较小,在生育后期,气孔导度下降幅度更大。与CK相比,生育前期单独增温处理气孔导度提高了17.4%~46.3%,在生育后期,随着增温处理时间的延长,气孔导度下降8.8%~63.1%,平均下降幅度为32%。进一步对不同处理之间气孔导度的变化进行分析,结果表明,生育前期气孔导度均表现为IT>CK>IT+IC,在生育后期,表现为CK>IT>IT+IC,CK、IT与IT+IC之间存在显著或极显著差异。可见,在马铃薯生育前期,增温处理使气孔导度大于对照,后期小于对照;随着环境CO2浓度升高,气孔导度逐渐降低,故增温加CO2浓度升高复合处理气孔导度小于对照。
2.2.3 马铃薯叶片蒸腾速率和水分利用效率的变化特征
分别在开花期与块茎膨大期不同阶段测定马铃薯叶片的蒸腾速率(图 3a),温度升高,马铃薯叶片蒸腾速率升高,IT处理最大,CK次之,IT+IC最小,IT处理叶片蒸腾速率较CK提高1.1%~12.1%之间,IT+IC较CK降低4.3%~27.2%。
叶片水平水分利用效率(WUE)是指蒸腾消耗单位重量的水分所同化的CO2的量,常用净光合速率与蒸腾速率的比值表示,即 WUE=Pn/Tr。在高CO2浓度下,马铃薯叶片的水分利用效率提高,其原因是净光合速率的提高和气孔导度的下降引起的蒸腾速率的降低,促进了水分利用效率的提高(图3b)。开花期、块茎膨大期马铃薯叶片WUE结果表明,CO2浓度升高,马铃薯叶片水分利用效率提高,处理间 WUE一致表现为 IT+IC>CK>IT,IT+IC处理较CK提高76.8%,IT较CK降低11.4%。生育期中后期,仍表现为IT+IC>IT>CK,且幅度较小;接近可收期,则CK>IT>IT+IC。
2.2.4 马铃薯叶片胞间CO2浓度的变化特征
试验结果表明,CO2浓度升高,叶片胞间CO2浓度显著高于 IT、CK(P<0.01),较 IT偏高31.3%~82.2%,平均增幅 67.5%,较 CK偏高40.3%~79.3%,平均增幅70.0%。IT细胞间CO2浓度略高于CK(图4)。

图4 不同处理马铃薯胞间CO2浓度变化Fig. 4 Changes the Intercellular CO2 concentration of potato with different treatments
2.2.5 马铃薯光响应曲线特征参数的响应特征
2.2.5.1 增温处理(IT)光响应曲线模型
增温处理(IT)光合光响应曲线采用非直角双曲线模型与Cubic函数分段拟合(图5a),其方程为:


图3 不同处理马铃薯蒸腾速率(a)水分利用效率(b)变化Fig. 3 Changes the transpiration rate (a) and the water use efficiency (b) of potato for different treatments
2.2.5.2 复合处理(IT+IC)光响应曲线模型

增温(IT)+CO2浓度升高(IC)复合处理(IT+IC)光合光响应曲线也采用非直角双曲线模型与 Cubic函数分段拟合(图5b)。其方程为:(R2=0.996,0≤I<1500 μmol⋅m-2⋅s-1);Pn=9×10-9x3-5×10-5x2+0.0894x-31.472(R2=0.897,1500 μmol⋅m-2⋅s-1≤I<2300 μmol⋅m-2⋅s-1)。
2.2.5.3 大田对照(CK)光响应曲线模型
大田对照(CK)光合光响应曲线完全应用非直角双曲线模型拟合(图5c),其拟合方程为:
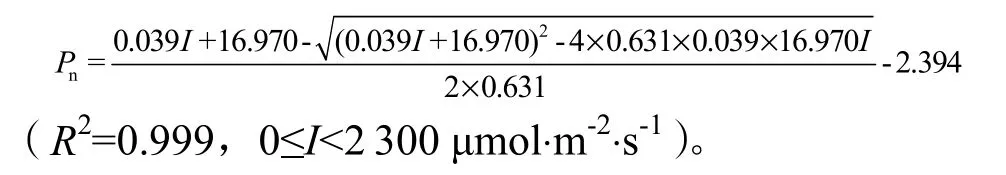
2.2.5.4 光响应曲线特征参数的响应特征及其阈值
对增温和CO2浓度升高背景下马铃薯光响应参数进行模拟(表 1),结果表明,单独增温(IT)、增温+CO2浓度升高(IT+IC)背景下,马铃薯叶片光补偿点(LCP)、表观量子效率α、最大净光合速率(Pnmax)一致表现为 IT+IC>IT>CK,IT、IT+IC处理光补偿点较CK提高72.2%和82.7%,分别为39.59、41.992、22.988 μmol·m-2·s-1。增温(IT)、复合处理(IT+IC)表观量子效率(α)较CK分别提高 0.003 μmol·m-2·s-1和 0.040 μmol·m-2·s-1。IT 处理及IT+IC处理叶片最大净光合速率(Pnmax)分别较CK提高0.4%和39.8%。试验表明,增温和CO2浓度升高背景下马铃薯叶片光合作用过程中光合同化作用与呼吸消耗相当时的光合有效辐射通量密度阈值提高,叶片在弱光条件下的光合能力增加,叶片的最大光合能力显著增加。
光饱和点(LSP)则表现为IT+IC>CK>IT,增温+CO2浓度升高(IT+IC)处理光饱和点(LSP)升高,表明增温和CO2浓度升高最大同化作用时光合有效辐射通量密度阈值增高。单独增温(IT)条件下,光饱和点(LSP)降低。
暗呼吸速率Rd表现为IT>CK>IT+IC,表明温度升高,暗呼吸速率升高,马铃薯叶片异化作用(即分解代谢)增强;增温的同时,增加CO2浓度,一定程度上缓解了增温造成暗呼吸速率升高。
2.3 CO2浓度升高与大气增温对马铃薯产量的影响特征
对不同处理马铃薯产量进行分析(表 2),单独增温(IT)株薯块重较CK明显降低,降低幅度为 39%,主要是由于温度升高影响了光合酶的活性,导致马铃薯光合速率降低,干物质积累减少。在IT+IC处理协同影响下,马铃薯株薯块重较CK提高,增幅为54.9%。屑薯率IT+IC处理较CK与IT处理明显提高。鲜茎重的变化趋势为 IT+IC>IT>CK,IT+IC和 IT较 CK分别增加 40.6%和19.5%。实际产量表现为IT+IC>CK>IT,单独增温处理(IT)产量最低,其原因是马铃薯为喜凉作物,气温升高不利于块茎的生长,气温过高会造成块茎生长停止。

表1 不同处理马铃薯光响应模拟参数Table 1 Simulation parameters of potato light response with different treatments
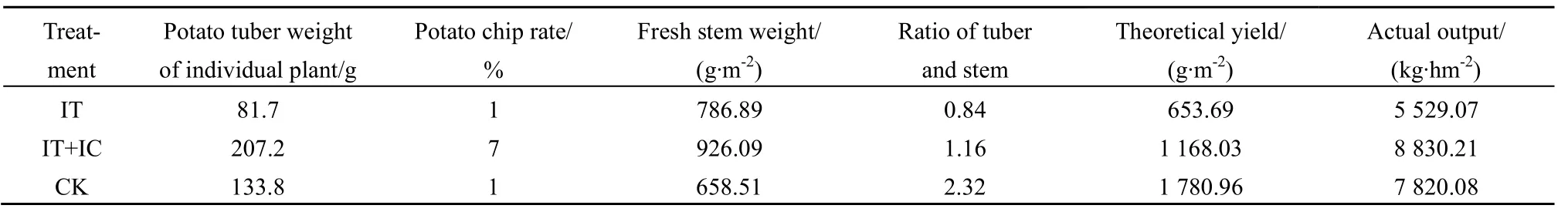
表2 不同处理下马铃薯产量结构变化Table 2 Potato yield structure change of different treatments
可见,在增温的同时增加CO2浓度,会使得叶片净光合速率和水分利用效率提高,鲜茎重和株块茎重表现为 IT+IC>IT>CK,实际产量也表现为IT+IC>CK>IT,干物重积累增多,实际产量增加。
3 讨论
净光合速率反映了植物在单位时间内积累有机物的能力,是作物生长发育最重要的指标。在营养生长期,马铃薯叶片净光合速率表现为增温加CO2浓度升高复合处理大于增温处理,且大于对照,此时段内胞间CO2浓度远大于单独增温处理和对照(陈根云等,2010);营养生长期处于马铃薯生育前期,此时段气温仍较低,尚未接近马铃薯生育的适宜温度,气温对马铃薯光合作用为正效应,增温加快了光合进程;在增温处理的同时,增加CO2浓度,则表现为胞间 CO2浓度远大于单独增温处理和对照,较高的胞间CO2浓度,为马铃薯叶片光合作用提供了充足的原料,使得作物净光合速率进一步提高(Fleisher et al.,2008)。增温和CO2浓度提高的叠加效应,使营养生长期增温加CO2浓度升高复合处理的净光合速率远大于其他处理和对照;同时马铃薯水分利用效率也相应提高,其值也高于单独增温处理和对照。(Carolina et al.,2017)。
由于前期增温加CO2浓度升高复合处理及增温处理马铃薯叶面积指数显著偏高,生长旺盛,在生育后期,叶面积指数的递减率也较快,叶片衰老速度加快(Miglietta et al.,1998;Lawson et al.,2001)),此时段内胞间CO2浓度较单独增温处理和对照亦有所降低,导致增温加CO2浓度升高复合处理净光合速率下降更为迅速,而马铃薯水分利用效率也随之下降。(Schapendonk et al.,2000)。
增温和CO2浓度升高背景下马铃薯叶片光合作用过程中光合同化作用与呼吸消耗相当时的光合有效辐射通量密度阈值提高,马铃薯叶片在弱光条件下的光合能力增加,马铃薯的最大光合能力也增加,最大同化作用时光合有效辐射通量密度阈值增高。单独增温条件下,光饱和点降低。由于温度升高,暗呼吸速率升高,马铃薯物异化作用(即分解代谢)增强;增温和CO2浓度升高协同影响,缓解了增温所造成的暗呼吸速率的升高。
通过试验研究了黄土高原半干旱区域增温与升高CO2浓度对马铃薯生理生态的影响,分析了马铃薯叶片光响应过程特征,建立马铃薯叶片光响应曲线模型,给出了叶片光响应过程的基本参数(叶子飘,2010)。然而,有关增温和 CO2浓度升高对马铃薯光合生理生态特征影响机理尚需在今后的研究中进行深化。
4 结论
(1)试验区近 59年气温呈极显著上升趋势,气温线性回归气候倾向率为 0.42 ℃·(10a)-1(r=0.808,n=59,P<0.001),20世纪70年代后气温持续上升,进入21世纪后升温尤为明显。近59年年际降水量呈减少趋势,降水量递减倾向率为-12.17 mm·(10a)-1(r=0.25,n=59,P=0.05)。
(2)在营养生长期,马铃薯叶片净光合速率表现为增温加CO2浓度升高复合处理大于增温处理,且大于对照;在生育后期,增温加CO2浓度升高处理净光合速率下降更为迅速。在马铃薯生育前期,增温处理气孔导度大于对照,后期小于对照;环境CO2浓度升高,气孔导度下降,马铃薯生育期CO2浓度升高处理气孔导度均小于对照。增温处理马铃薯叶片蒸腾速率升高,高于对照和其他处理,后期低于对照。增温加CO2浓度升高复合处理马铃薯叶片水分利用效率提高,明显高于对照和单独增温处理。增温加 CO2浓度升高复合处理叶片胞间 CO2浓度显著高于对照和增温处理。
(3)增温加CO2浓度升高复合处理马铃薯实际产量明显高于对照和单独增温处理。单独增温处理产量偏低,其原因是马铃薯为喜凉作物,气温升高不利于块茎的生长,气温过高会造成块茎生长停止,在增温的同时增加CO2浓度,可以使马铃薯叶片净光合速率和水分利用效率提高,干物重积累增多,经济产量增加。
AWADA T, RADOGLOU K, FOTELLI MN, et al. 2003. Ecophysiology of seedlings of three Mediterranean pine species in contrasting light regimes [J]. Tree Physiology, 23(1): 33-41.
BALY E C. 1935. The kinetics of photosynthesis [J]. Proceedings of the Royal Society of London Series B ( Biological Sciences), 117(804):218-239.
CAROLINA L X, AVILA A, TOLABA A, et al. 2017. Field responses of potato to increased temperature during tuber bulking: Projection for climate change scenarios, at high-yield environments of Southern Chile[J]. agricultural and forest meteorology, 239: 192-201.
DAMESIN C. 2003. Respiration and photosynthesis characteristics of current 2 year stems of Fagus sylvatica: from the seasonal pattern to an annual balance [J]. New Phytologist, 158(3): 465-475.
FLEISHER D H, TIMLIN D J, REDDY V R, et al. 2008. Interactive effects of CO2and water stress on potato canopy growth and development [J].Agronomy Journal, 100(3): 711-719.
GOUDRIAAN J, DE RUITER H E. 1983. Plant growth in response to CO2enrichment, at two levels of nitrogen and phosphorous supply. 1. dry matter, leaf area and development [J]. Netherlands Journal of Agricultural Science, 31: 157-169.
IPCC. 2013. Climate change 2013: the physical science basis [M].Cambridge: Cambridge University Press.
KU S B, EDWARDS G E, TANNER C B. 1977. Effects of light, carbon dioxide, and temperature on photosynthesis, oxygen inhibition of photosynthesis, and transpiration in Solanum tuberosum [J]. Plant Physiology, 59(5): 868-872.
LAWSON T, CRAIGON J, TULLOCH A M. et al. 2001. Photosynthetic responses to elevated CO2and O3in field-grown potato (Solanum tuberosum) [J]. Journal of Plant Physiology, 158(3): 309-323.
MIGLIETTA F, MAGLIAGO B, BINDI M, et al. 1998. Free air CO2enrichment of potato (Solanum tuberosum L.): development, growth and yield [J]. Global Change Biology, 4(2): 163-172.
MIKO U F K, GRAHAM D F. 1987. Investigation of the CO2dependence of quantum yield and respiration in Eucalyptus pauciflora [J]. Plant Physiology, 83: 1032-1036.
MORENO S A, WEISS A, PAPAROZZI E T, et al. 2002. Stability of leaf anatomy and light response curves of field grown maize as a function of age and nitrogen status [J]. Journal of Plant Physiology, 159(8):819-826.
PRADO C H B A, MORAES J A P V. 1997. Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field condition [J]. Photosynthetica, 33(1): 103-112.
RAWSON H M. 1988. Effect of high temperatures on the development and yield of wheat and practices to reduce deleterious effects [M]// KLATT A R. Wheat Production Constraints in Tropical Environments. Mexico,D.F, CIMMYT: 44-62.
ROBERT E S, MARK A, JOHN S B. 1984. Kok effect and the quantum yield of photosynthesis [J]. Plant Physiology, 75: 95-101.
SCHAPENDONK H C M, VAN OIJEN M, DIJKSTRA P, et al. 2000.Effects of elevated CO2concentration on photosynthetic acclimation and productivity of two potato cultivars grown in open-top chambers[J]. Australian Journal of Plant Physiology, 27(12): 1119-1130.
SICHER R C, BUNCE J A. 1999. Photosynthetic enhancement and conductance to water vapor of field-grown Solanum tuberosum (L.) in response to CO2enrichment [J]. Photosynthesis Research, 62(2):155-163.
THORNLEY J H M. 1976. Mathematical Models in Plant Physiology [M].London: Academic Press.
THORNLEY J H M. 1998. Dynamic model of leaf photosynthesis with acclimation to light and nitrogen [J]. Annals of Botany, 81: 421-30.
WHEELER R M, MACKOWIAK C L, YORIO N C, et al. 1999. Effects of CO2on stomatal conductance: do stomata open at very high CO2concentrations [J]. Annals of Botany, 83(3): 243-251.
WHEELER R M, TIBBITTS T W, FITZPATRICK A H. et al. 1991. Carbon dioxide effects on potato growth under different photoperiods and irradiance [J]. Crop Science, 31(5): 1209-1213.
XIAO G J, ZHANG F J, QIU Z J, et al. 2013a. Response to climate change for potato water use efficiency in semi-arid areas of China [J].Agricultural Water Management, 127(8): 119-123.
XIAO G J, ZHENG F J, QIU Z J, et al. 2013b. Impact of climate change on water use efficiency by wheat, potato and corn in semiarid areas of China [J]. Agriculture, Ecosystems & Environment, 181(181):108-114.
YE Z P, YU Q, KANG H J. 2012. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions [J].Photosynthetica, 50(3): 472-476.
YE Z P. 2007. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa [J]. Photosynthetica, 45: 637-640.
ZHOU J B, WANG C Y, ZHANG H, et al. 2011. Effect of water saving management practices and nitrogen fertilizer rate on crop yield and water use efficiency in a winter wheat-summer maize cropping system[J]. Field Crops Research, 122(2): 157-163.
陈根云, 陈娟, 许大全. 2010. 关于净光合速率和胞间CO2浓度关系的思考[J]. 植物生理学通讯, 46(1): 64-66.
国家气象局. 1993. 农业气象观测规范(上卷)[M]. 北京: 气象出版社:1-59.
李子涵, 杨晓晶. 2016. 世界及中国马铃薯产业发展分析[J]. 中国食物与营养, 22(5): 5-9.
魏凤英. 2007. 现代气候统计诊断与预测技术[M]. 北京: 气象出版社:175-181.
闫小红, 尹建华, 段世华, 等. 2013. 四种水稻品种的光合光响应曲线及其模型拟合[J]. 生态学杂志, 32(3): 604-610.
姚玉璧, 王润元, 赵鸿. 2013. 甘肃黄土高原不同海拔气候变化对马铃薯生育脆弱性的影响[J]. 干旱地区农业研究, 31(2): 52-58.
叶子飘. 2007. 光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J]. 生态学杂志, 26(8): 1323-1326.
叶子飘. 2010. 光合作用对光和 CO2响应模型的研究进展[J]. 植物生态学报, 34(6): 727-740.
张强, 王润元, 邓振镛. 2012. 中国西北干旱气候变化对农业预生态影响及对策[M]. 北京: 气象出版社: 148-152.
赵鸿, 王润元, 王鹤龄, 等. 2013. 半干旱雨养区苗期土壤温湿度增加对马铃薯生物量积累的影响[J]. 干旱气象, 31(2): 290-297.
赵年武, 郭连云, 赵恒和. 2015. 高寒半干旱地区马铃薯生育期气候因子变化规律及其影响[J]. 干旱气象, 33(6): 1024-1030.
中国气象局气候变化中心. 2017. 中国气候变化监测公报(2016年)[M].北京: 科学出版社: 1-10.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
新课程·下旬(2017年7期)2017-08-14
中国铸造装备与技术(2015年5期)2015-12-10
西北林学院学报(2015年5期)2015-01-03
