
广东康禾自然保护区地面苔藓种群分布与格局分析
2018-06-07 09:32蒋甜甜杨学成钟泳林徐明锋唐启明苏志尧
生态环境学报 2018年5期
蒋甜甜,杨学成,钟泳林,徐明锋,唐启明,苏志尧
华南农业大学林学与风景园林学院,广东 广州 510642
植物种群的分布格局是植物生态学研究的经典主题,对研究物种的形成、群落构建及物种与环境之间的关系具有重要的指导意义(贾成朕等,2017;张金屯,2011;Fibich et al.,2016)。植物种群的分布格局是指种群个体在水平空间的配置方式,不仅反映了种内个体在水平空间上的相互关系,也是种群个体自身生物学特性、种群关系及其对生境长期适应和综合利用的空间表现形式,也反映出种间关联及与其他种群之间的相互作用和影响过程(赵峰侠等,2007;张瑾等,2013;Nguyen et al.,2016)。因此,种群分布格局分析已被认为是解释物种与生境的关联、回答物种生存策略、物种共存及其在群落构建中的作用等生态学问题的强有效工具(Ebert et al.,2016;Fibich et al.,2016)。研究植物种群的分布格局主要从两方面开展,一是通过离散指数或拟合分布模型对种群分布类型进行判定,普遍认为有3种分布类型,即随机分布、均匀分布和集群分布(梅军林等,2017);二是对植物种群集群分布的规模和强度进行研究(兰国玉等,2003)。
截至目前,研究植物种群分布格局的检验指标已有10余种(兰国玉等,2003)。在对集群分布的格局规模进行描述方面,Greig-smith利用区组样方法差法(Blocked Quadrat Variance Method,BQV)进行格局分析,克服了传统单一取样分析难以全面客观反映种群分布格局的不足(李海涛,1995;郭连金,2014;Dai et al.,1997)。后来出现的双项轨迹方差分析(Two-term local quadrat variance technique,TTLQV),则可不受 n/2的分组限制,对研究连续样方内植物种群的空间分布格局尤为有效(Dai et al.,1997);该方法自提出后,在陆地生态学,尤其在森林、草原植物群落的研究中已得到大量运用(李素清等,2006;李素清等,2011;Hanewinkel,2004),但至今其在苔藓植物研究中的应用还鲜见报道。
苔藓植物是一类体型细小,结构相对简单,以孢子繁殖,可代表水生向陆生进化的重要类群(Miller,1982),其种类数量仅次于种子植物的高等植物(Song et al.,2016),在植物的演化中具有特殊的地位(Miller,1982;卜兆君等,2009)。目前全世界记录的苔藓植物超过 20000种(Song et al.,2015),根据最新的资料统计,中国苔藓植物共记录 151科 602属 3045种,占世界多样性的18.8%,是生物多样性的重要组成(王利松等,2015)。随着近几年对苔藓植物生态功能的研究报道越来越多,人们越来越多的关注和重视苔藓植物(田维莉等,2011;陈勤等,2013;姜苹红等,2015)。中国对苔藓植物生态学的研究起步相对较晚,不像对维管植物尤其是木本植物的群落特征及其与环境间相互作用的研究已经趋于成熟,苔藓植物的生态学研究仍处于初步探索阶段(郭磊等,2016;陈云等,2017),国内对苔藓植物的研究以不同研究地区苔藓物种多样性报道居多,偶见附生苔藓空间分布格局的报道(许宁等,2017),但地面苔藓种群分布格局和规模变化的研究基本处于空白状态。本文利用数学模型 Poisson分布和方差均值方法来检验广东康禾自然保护区地面苔藓植物群落的分布格局类型,采用TTLQV来揭示格局规模的变化,以此探讨亚热带常绿阔叶林中苔藓植物的分布格局及格局规模,并探讨其成因,旨在丰富亚热带森林苔藓植物的研究,为今后研究保护苔藓植物多样性提供参考意见。
1 材料与方法
1.1 研究地概况
研究区域位于广东东源康禾省级自然保护区内 , 地 理 坐 标 为 115°04′27″E~115°09′41″E ,23°44′37″N~23°52′16″N,面积 6484.8 hm2;保护区属半山区半丘陵地带,总体呈东北-西南走向,最高峰为白石岗(海拔893.3 m)。区内土壤主要有黄壤、红壤、赤红壤等,成土母岩主要有花岗岩和砂岩。气候属亚热带南缘季风气候,年平均温度 20.3~21.1 ℃,年平均降水量2142.6 mm,光热充足,降雨充沛。地带性乔、灌木植被是以壳斗科、山茶科树种为主的亚热带常绿阔叶林(He et al.,2017)。
1.2 样地设置及植物调查
于2016年8月在保护区内选择苔藓植物分布较为丰富的区域,设置2 hm2样地,并进行群落学调查,样地海拔 209.1~349.49 m,坡度 5°~70°,坡向为西南方向。利用全站仪(Total Station)把样地划分50个20 m×20 m的网格样方,分别在每个网格样方的中心处、1/4处、3/4处设置1个2 m×2 m样方单元,共 5个,在每个样方单元中设置 4个1 m×1 m小样方对地面苔藓进行调查取样,为确保实际调查时准确记录苔藓植物,实际测量时将20 cm×20 cm网格铁筛(与小样方大小一致)置于小样方上,记录相同种类苔藓出现在网格交叉处的次数,用于计算每种苔藓的盖度,并记录每个小样方中苔藓植物种名、网格样方的生境。对于实地无法确定的苔藓,采集其标本,装入事先准备好的牛皮封口信封中,带回实验室,借助显微镜进行形态学鉴定,分类学鉴定参考《中国生物物种名录——苔藓植物》和《广东苔藓志》(贾渝等,2013;吴德邻,2013)。样地内主要苔藓植物有东亚拟鳞叶藓(Pseudotaxiphyllum pohliaecarpum)、细指苔(Kurzia gonyotricha)、三裂鞭苔(Bazzania tridens)、双齿裂萼苔(Chiloscyphus latifolius)、细叶小羽藓(Haplocladium microphyllum)、粗肋凤尾藓(Fissiden slaxus)和变异多褶苔(Spruceanthus polymorphus)等。
1.3 分析方法
本研究采用苔藓植物的频度计算种群分布和聚集强度,并对苔藓植物的盖度百分比进行等级划分,用于双向轨迹方差分析以揭示苔藓植物种群分布格局的大小和斑块尺度。苔藓植物的盖度百分比等级依次为:0(<5%),1(5%~15%),2(15%25%),3(25%~35%),4(35%~45%),5(45%~55%),6(55%~65%),7(65%~75%),8(75%~85%),9(85%~95%),10(≥95%)(Usher,1983)。
研究植物种群分布的方法有很多,判别种群分布的数学模型主要有 Poisson分布、负二项分布和正二项分布(马钦彦,2009)。在研究植物种群分布中,最常用的是 Poisson分布模型,但为了避免不同模型的局限性,研究者常采用多种检验方法来判定分布类型。本文采用了6种指标对研究区域的苔藓植物进行种群分布格局分析,其中偏离系数和χ2值均通过拟合检验获得P值来验证Poisson分布模型,以更准确地判断种群的分布格局;聚集强度则采用了丛生指数I、平均拥挤度m*指数和聚块性指数m*/m、标准化的Morista’s分散指数进行测定。丛生指数I和平均拥挤度m*主要受到种群密度的影响,是种群数量和密集程度的重要表征(董秋莲等,2013),特别是平均拥挤度m*,表征了样方单元内每个植物个体的邻体数,邻体数越大,植物个体受到的拥挤效应越强,相邻个体对其自身生存资源的竞争越强烈。m*/m值越大,聚集强度越大。标准化的Morista’s分散指数与聚块性指数m*/m类似,均不受种群密度和样本大小的影响,仅考虑空间格局自身的性质,既能作为集群程度的衡量指标,也能表示物种对空间的扩展和延伸程度(赵峰侠等,2007;李亚等,2017)。
1.3.1 分布格局类型的确定
Poisson分布模型对应的是随机分布。其函数为:

式中,P(X)为N个抽样中出现的X个个体的概率;X为物种个体数,X=0, 1, 2, 3…….;m为每个样方中的平均个体数。
方差均值比是根据对 Poisson分布的偏离程度来确定种群分布格局。V代表方差,代表均值,方差比为,若=1,则个体完全符合Poisson分布,呈随机分布;若V>1,为集群分布;若<1,为均匀分布(张金屯,2011)。
检验通过检验不同个体的样方频度的观测值(Vobs)和Poisson分布预测值(Vpre)之间的差异来判断种群分布类型,若χ2与存在显著差异,为非随机分布,反之则为聚集分布(张金屯,2011),公式如下:
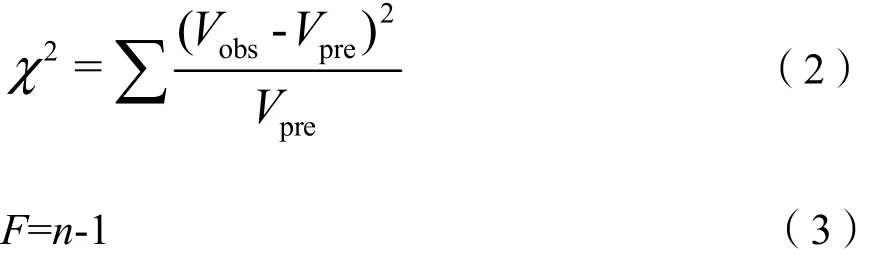
式中,F为自由度;n为组数。

式中,S2为样本方差;X为样本均值;I为丛生指数。当I=0时,呈随机分布;当I<0时,呈均匀分布;当I>0时,呈聚集分布。
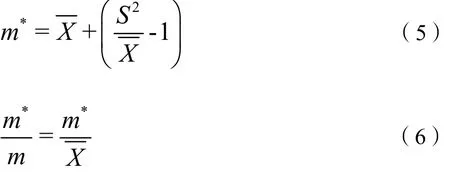
当m*/m=1时,为随机分布;当m*/m<1时,为均匀分布;当m*/m>1时,为聚集分布。
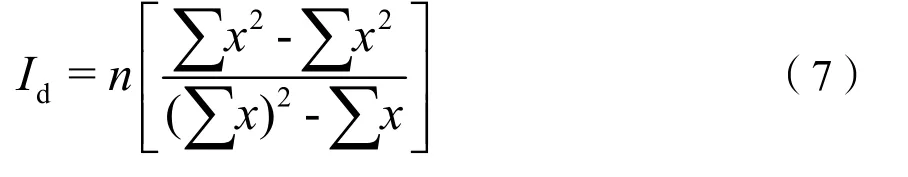
式中,表示总样方数;表示样方数的平方和;n表示样方数;Id为标准化Morista’s分散指数。若Id=0时,为随机分布;若Id<0时,为均匀分布;若Id>0时,为聚集分布(Szmyt,2014)。
1.3.2 双项轨迹方差分析法(Two-term local quadrat variance technique,TTLQV)
双项轨迹方差分析法是研究植物种群分布格局规模的主要方法之一,不仅能反映植物单个种群的斑块大小,当其与除趋势对应分析(Detrended correspondence analysis,DCA)联合时,还是研究植物种群组分布格局的重要手段(张金屯,2005;张金屯等,2006)。公式如下:
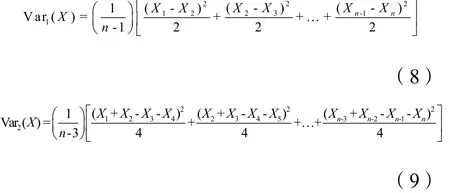
式中,Var1(X)表示区组1的方差计算;Var2(X)表示区组2的方差计算;n为样方数;X1为样方1的植物个体数,在此表示为苔藓植物盖度百分比的等级值,其他区组的计算依次类推。
除趋势对应分析在软件PC-ORD中完成,分布格局分析在软件 Ecological Methodology 7.2中完成,图形制作在软件STATISTICA 8.0中完成。
2 结果与分析
2.1 苔藓植物的种群分布格局
样地中共记录到苔藓植物37种,隶属于23科33属,其中重要值最高的物种是东亚拟鳞叶藓和细指苔;其次是三裂鞭苔、双齿裂萼苔。对单个物种的格局分析不仅能表示物种的数量特征,还可以进一步对物种水平结构做出定量描述(张瑾等,2013)。研究区域苔藓植物的分布类型与聚集强度见表 1,结果表明:苔藓植物单种的分布格局总体呈集群分布,仅有藓类植物桧叶白发藓(Leucobryum juniperoideum)的χ2检验结果表现不同,呈随机分布。苔类植物中细指苔受到的拥挤效应最高(m*=10),而在藓类植物中东亚拟鳞叶藓受到的拥挤效应最高(m*=13.04),细叶小羽藓(m*=11.85)次之。在仅考虑空间格局自身性质的情况下,标准化的Morista’s分散指数值与m*/m呈相同的增长趋势,其中苔类植物树生扁萼苔(Radula obscura)和长叶羽苔(Plagiochila flexuosa)种群的聚集程度最大,分别为14.79和14.45;而优势物种细指苔、三裂鞭苔、双齿裂萼苔的聚集强度分别为 1.99、1.89、1.93。与苔类植物相比,藓类植物的优势种和非优势种在聚集强度分布上具有相似的分布特征,样地的藓类植物非优势物种日本扁枝藓(Homalia trichomanoides var. japonica)的聚块性指数(11.05)是绝对优势物种东亚拟鳞叶藓的聚块性指数(1.15)的 9.6倍。说明非优势物种的聚集强度比优势物种的聚集强度更高,更喜欢集中分布在某些生境中,而优势物种在该区域分布较广,生存能力更强,生态位幅度更宽,但与其他个体的资源竞争更强烈。总体而言,该区域苔类植物种群和藓类植物种群的总聚集强度差异不明显,但藓类植物种群间相邻个体的拥挤度高于苔类植物种群,其物种间对样地生态资源的竞争更加强烈。
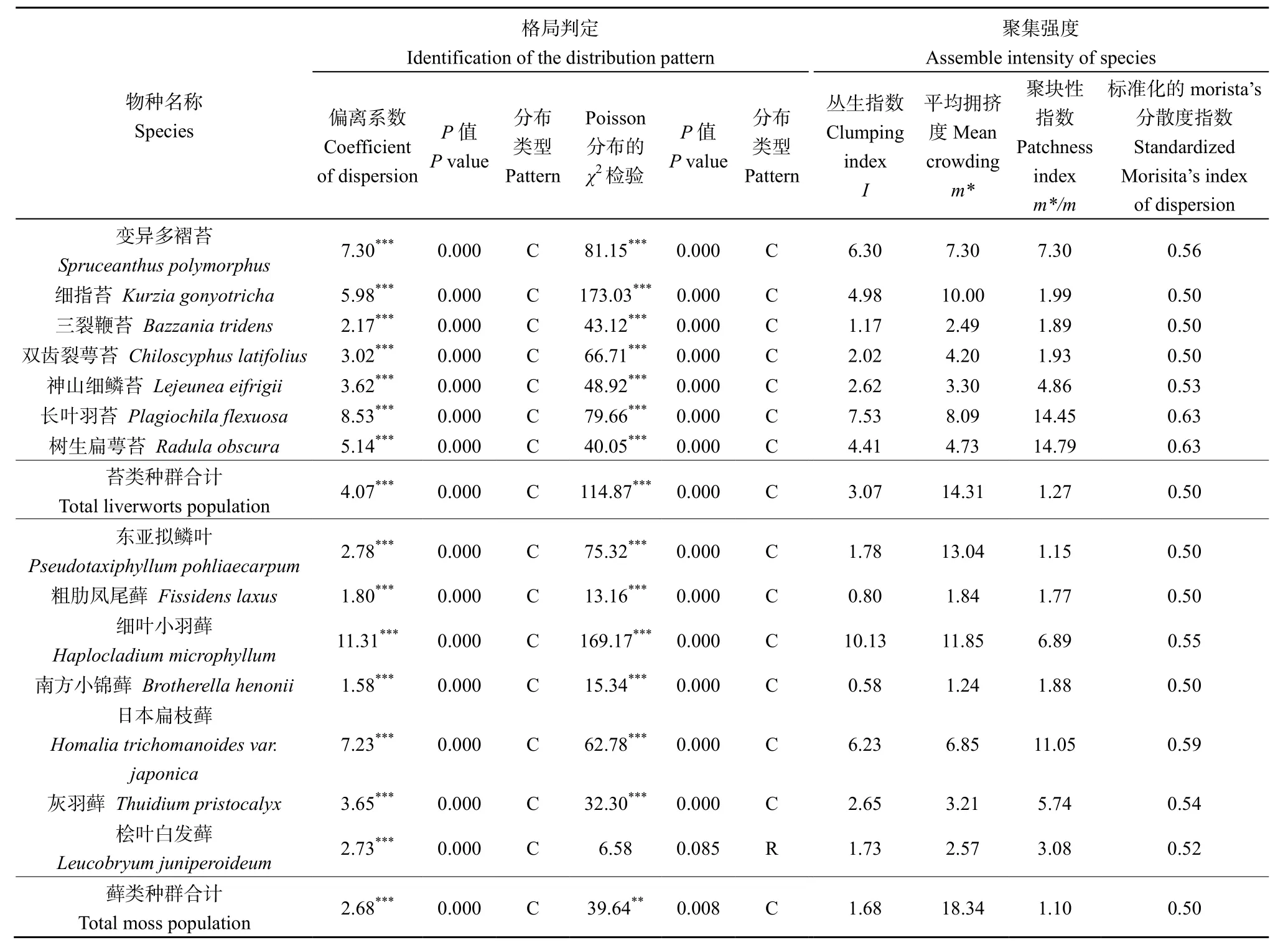
表1 样地苔藓植物的格局分布判定与聚集强度Table 1 Identify distribution pattern and Assemble intensity of bryophytes
2.2 苔藓植物的TTLQV格局分析
集群分布的种群个体通常聚集成大小不等的斑块镶嵌在一起,这种镶嵌组合在一起的斑块被称为种群格局规模(张维等,2016)。本研究TTLQV以区组大小为横坐标,盖度百分比得分值的方差为纵坐标作图(图1、图2、图3)。图上曲线区组所对应出现的峰值是解释分布格局强度和斑块尺度的重要指标,即峰值对应的区组大小代表了植物种群的格局规模(Szmyt,2014;王继和等,2010)。在方差区组图中,若出现的方差峰值急剧增加,则表明聚集强烈且呈大斑块聚集;若峰值相对较低,则表示为低强度的聚集,聚集斑块为小斑块的聚集规模(Szmyt,2014;Krebs,1989);若同时出现多个峰值则表示该研究对象有多个分布格局或有多个小格局镶嵌于大格局中(Yang et al.,2017)。研究区域的苔类植物和藓类植物的方差区组值总体均呈先增加后减少的趋势(图1、图2、图3)。
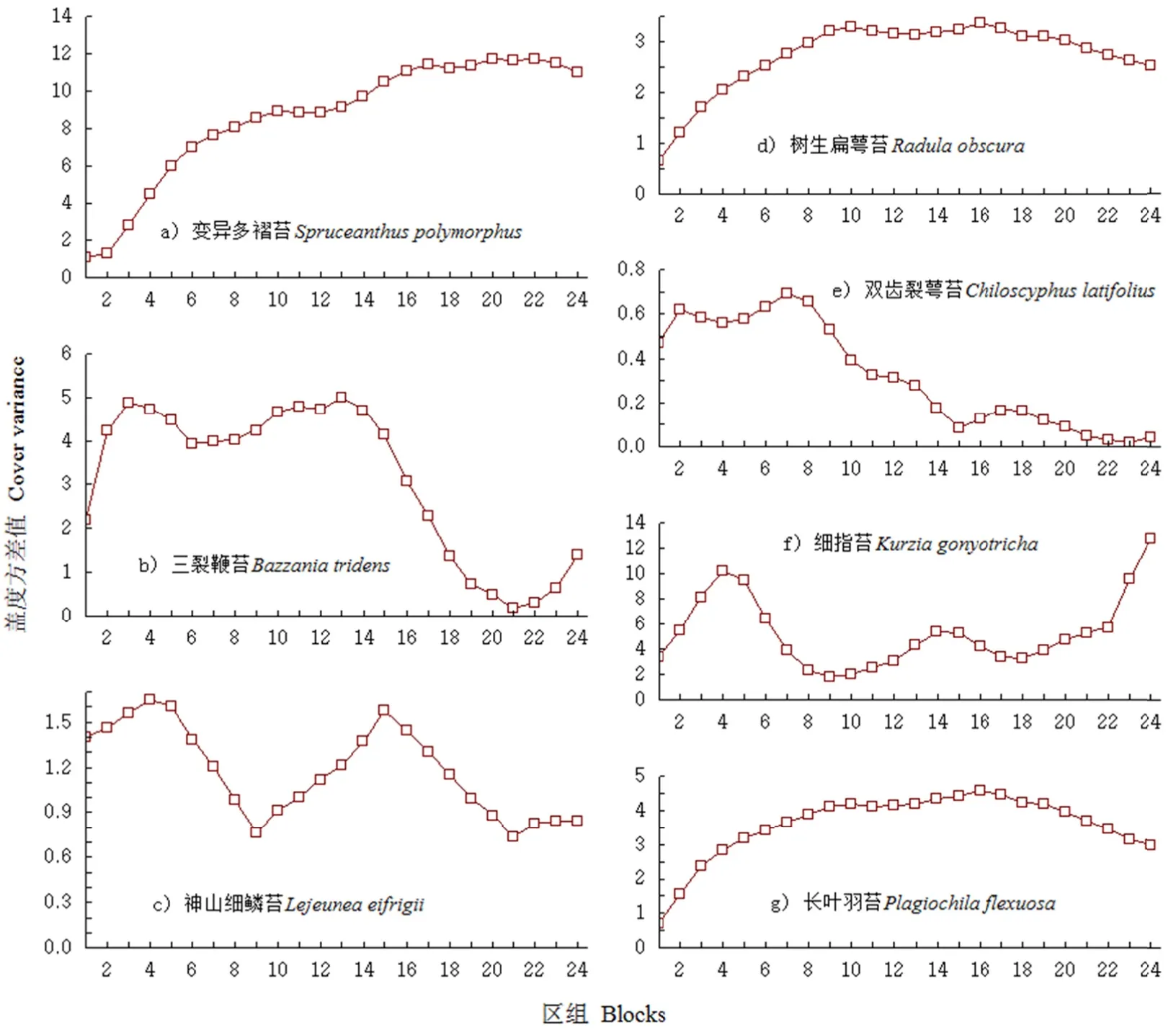
图1 苔类植物种群分布格局的变化Fig. 11 Changes in poppulation distributioon patterns of liverrw orts
图1所示为7种苔类植物的格局规模变化,树生扁萼苔(Radula obscura)和长叶羽苔(Plagiochila flexuosa)的总体增长趋势基本一致,树生扁萼苔和长叶羽苔均在区组 16出现轻微上升达到峰值后下降,呈小斑块的聚集规模,未发现大斑块的聚集规模(图1d,图1g)。变异多褶苔的格局规模未能明显展现,仅在区组20~22,呈现轻微波动(图1a)。细指苔作为研究区域的优势物种,在区组4和区组24呈现大斑块聚集规模,在区组14和区组15呈现两个小斑块聚集规模。神山细鳞苔与细指苔相似,在14、15、24都呈现斑块聚集,不同的是神山细鳞苔在区组4和区组15呈现大斑块聚集规模,在区组24呈现小斑块聚集(图 1c,图 1f)。三裂鞭苔的大斑块出现在区组3和区组13,小斑块出现在区组24(图 1b)。双齿裂萼苔的大斑块出现在区组2和区组7,小规模的斑块出现在区组17和区组18(图11e)。从物种分布格局规模看,物种间不同尺度的格局规模,说明了不同的生物个体对样地环境的选择不同,而多个物种在区组24出现斑块聚集,说明随着研究尺度的增加,生境变得多样,适宜物种生存的机会增加,故多个物种在区组24出现斑块。
图2所示为7种藓类植物的格局规模,日本扁枝藓在区组17和区组119出现轻微的峰值波动,但整体格局规模不明显(图 2a);东亚拟鳞叶藓是研究区域中分布最广的物种,出现多个聚集斑块,最大的斑块规模出现在区组19,小斑块规模依次出现在区组 2、8、9、22、24(图 2dd)。南方小锦藓(Brotherellla henonii)与灰羽藓(Thuiddium pristocalyx)出现斑块的区组一致,格局规模正好相反,南方小锦藓在区组4呈大斑块规模,在区组22、23呈小斑块规模;而灰羽藓在区组4呈小斑块的规模,在区组 22、23呈大斑块规模。细叶小羽藓与灰羽藓的斑块规模相似,区别仅在于细叶小羽藓在区组5有1个小斑块聚集规模,在区组22呈大斑块聚集,区组23未出现斑块聚集(图 2e,图2f,图 2g)。桧叶白发藓在区组 5呈大斑块聚集规模,小斑块聚集规模不明显。粗肋凤尾藓在区组 66呈大斑块聚集规模,在区组14和区组24呈小斑块聚集规模(图22b,图 2c)。从藓类植物单种的聚集规模看,藓类单个物种间的聚集区组更加接近,聚集规模更加明显,且3个单种出现斑块的区组基本一致,出现的格局规模略有差异,结合表1可以发现,3个物种中细叶小羽藓的平均拥挤度m*最高,聚块性指数m*/m也最大,此结果表明这3种藓类植物可能对研究区域某一生境较喜爱,但由于细叶小羽藓对生态位的竞争能力强于其他两个物种,虽然细叶小羽藓在区组4呈小斑块聚集,但从盖度方差值可以看出,其格局规模比灰羽藓、南方小锦藓大。因此,这33种藓类植物的种间竞争能力由大到小依次为细叶小羽藓、灰羽藓、南方小锦藓。
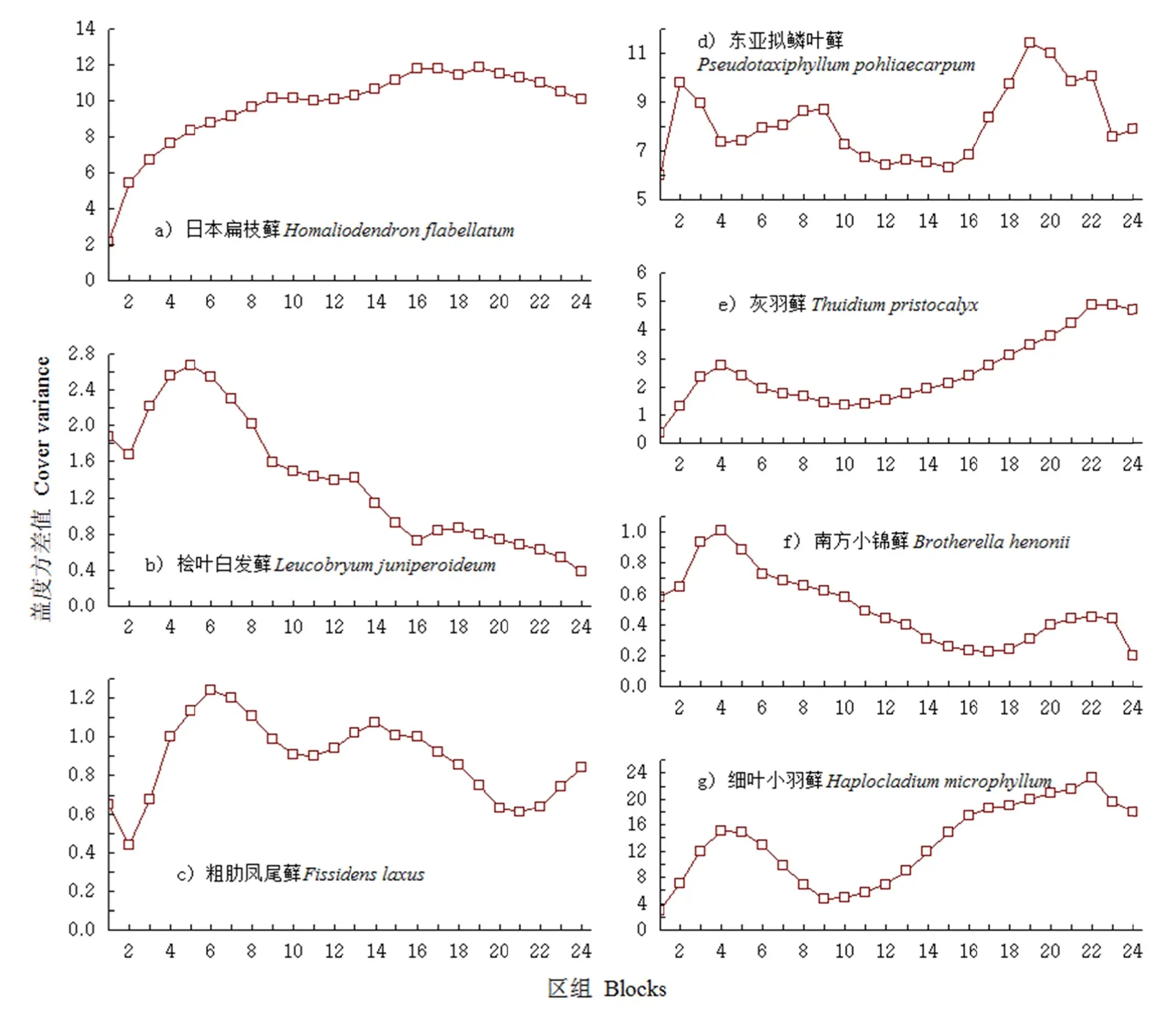
图2 藓类植物种群分布格局的变化Fig. 2 Changes in population distributtion patterns of moosses
2.3 苔藓植物种群组的TTLQV格局分析
图3 所示为苔类种群组、藓类种群组和研究区域总苔藓植物群落的格局规模。种群组的格局是指研究区域内所有苔类或藓类植物物种联合形成的格局,分别先对苔类种群组、藓类种群组和样地总的苔藓植物个体的盖度百分比进行DCA排序,得到3个轴的DCA排序轴值,分别以苔类、藓类和总苔藓植物的第一轴为基础对其种群组进行TTLQV格局分析(图33)。图3直观地展现了研究区域苔藓植物格局规模的变化,苔类植物在区组5呈大斑块聚集,在区组14、15呈小斑块聚集,总体分布格局呈小格局镶嵌于大格局中,其图形走势与苔藓植物更加接近。藓类植物在区组5、区组16、区组 22都呈现较大的峰值波动,呈大斑块聚集,其分布格局为大格局分布。苔藓植物在区组4和区组5的峰值接近,两者都呈大斑块聚集,在区组16呈小斑块聚集,其分布格局为小格局镶嵌于大格局中。从种群组的分布格局看,苔类植物与样地总苔藓植物的分布格局更加接近,都为小格局镶嵌于大格局中,这一结果说明苔类植物在样地苔藓植物群落分布格局中占主导作用。苔类植物和藓类植物种群组在不同的区组出现斑块聚集,说明苔类植物和藓类植物对样地生态环境具有不同的喜好,两者均在区组5出现斑块聚集(区组5的样方中具有良好的生态环境,适合苔藓植物生存),两者总体分布格局的差异可能由种群竞争和物种对资源生态位的侵占效益不同造成。这一结果解释也可由表1平均拥挤度得以证明,虽两者的侵占效应未见明显差异,但藓类植物受到相邻个体的拥挤效应比苔类植物高,说明在同一生态环境下,藓类植物的种内竞争强于苔类植物。
3 讨论
植物种群分布格局的形成一方面受物种自身的生态特性和种群关系的影响,另一方面由栖息生境的环境因子所决定(许宁等,2017;张旭等,20115)。研究结果显示,区内的苔藓植物总体呈集群分布,群落中重要值大的几个优势种(东亚拟鳞叶藓、细指苔、三裂鞭苔、双齿裂萼苔等)聚集指标相对较低,特别是在研究区域内占绝对优势地位的东亚拟鳞叶藓,其聚块性指数最低;而非优势物种长叶羽苔和树生扁萼苔的聚快性指数最高,这与植物的种间竞争和生境适宜性密切相关。在同一生态环境下,优势物种在群落中占据绝对的优势地位,竞争环境资源的能力更强,优胜劣汰,强势的物种生长更加旺盛,对环境适应能力更强,种群分布更广,导致聚集强度较低。
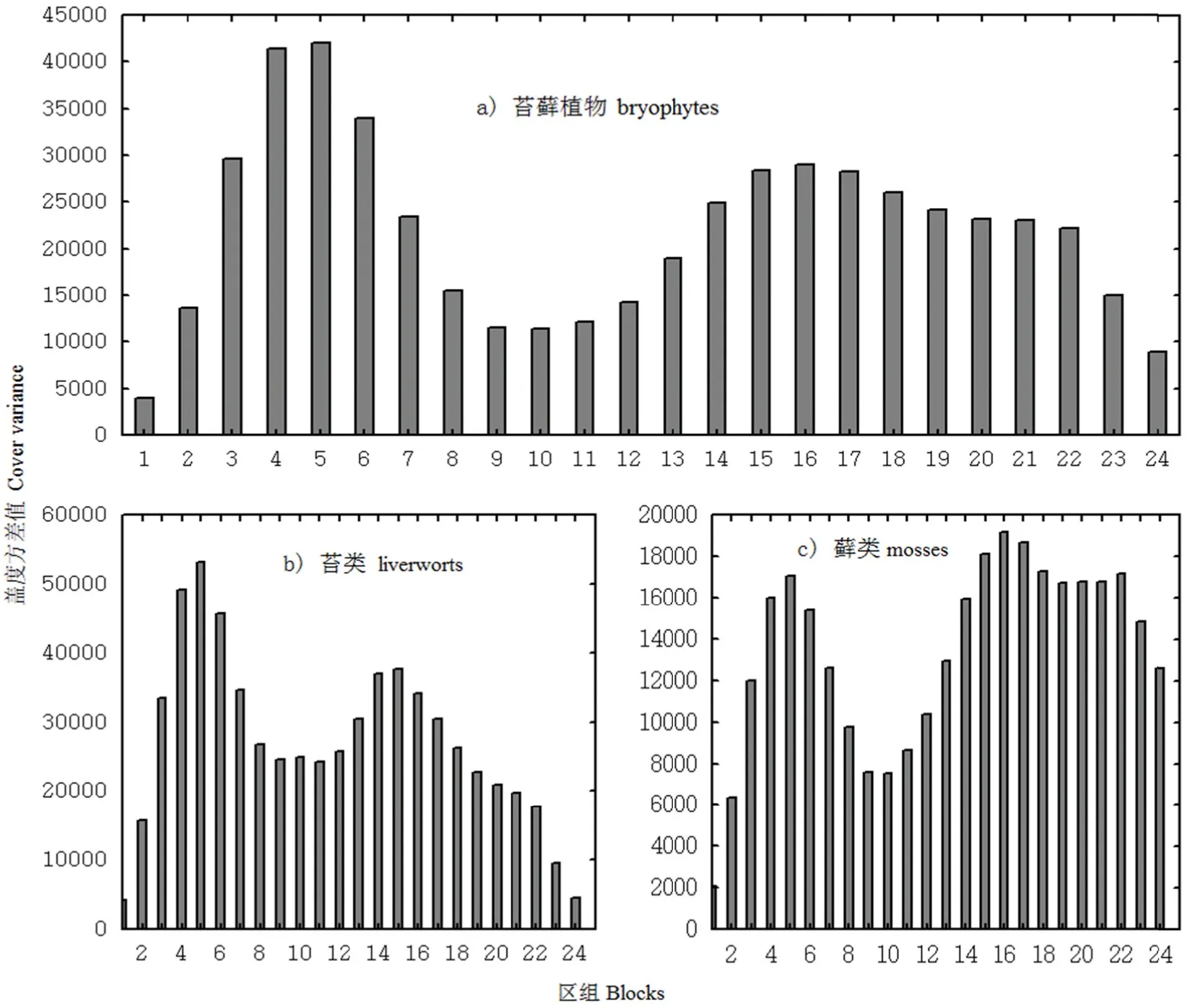
图3 不同类群苔藓植物种群组的格局变化Fig. 33 Change in bryoophyte populationdistribution patterrn s by groups
种群分布格局规模的变化显示,多个种群(细指苔、神山细鳞苔、三裂鞭苔等)在区组4、5、15、22、24均有斑块聚集,其规模有所差异,这与植物的生物学特性相关,如细指苔和三裂鞭苔属于同科植物,具有相似的生活特征,喜欢生长在树干基部;而神山细鳞苔则喜欢生长在腐木和石面上。苔类植物变异多褶苔、树生扁萼苔、长叶羽苔与藓类植物日本扁枝藓几乎具有相似的分布特征,整体分布格局变化不显著,这可能与其生活习性相关,均喜欢生长在岩石表面或树干上。一般认为,植物种群自身特性是决定小规模聚集的主要原因,如通过无性繁殖和种子传播的方式形成小斑块聚集;大斑块的聚集主要是植物从原分布中心扩散,多受生境异质性的影响(张维等,2016;张旭等,2015;赵峰侠等,2007)。本研究结果显示,不论是苔类植物还是藓类植物物种,其聚集规模以小斑块居多,仅有重要值大的单种多呈现大斑块聚集规模,这一方面可能与苔藓植物的自身生理特性紧密相连,另一方面可能受生境和种群关系的影响。苔藓植物是一类特别喜阴喜湿的物种,多分布在潮湿、阴暗的环境下(艾应伟等,2010),作为原始的高等植物,其精巧的繁殖方式高度依赖外界环境,在生存资源受限的条件下,苔藓植物的有性生殖受到限制,无性繁殖成为苔藓植物延续与扩张的主要方式(He ett al.,2016;王中生等,20003)。研究表明,在热带和亚热带森林中,基质类型、凋落物、光、热、海拔、坡度等地形因子是影响苔藓植物生长重要因子(Coorrales et al.,2010;Manndl et al.,20009)。对地面苔藓而言,土壤是其生长的主要基质,凋落物是阻碍其生长的主要物质。本研究样地枯枝落叶层较厚,凋落物是地面苔藓竞争资源环境的主要竞争者,故样地苔藓植物的格局规模以小斑块居多。也有研究表明,海拔是限制苔藓植物组成与分布的主要因子,坡度越陡峭的地方苔藓植物生长得越好(Soong et al.,20015;刘蔚秋等,2007)。研究区域部分样方单元位于山脚林缘处,海拔相对较低,受干扰多,易形成不同的局部小生境;部分样方单元位于山顶,地面相对平坦,不利于苔藓生长;有些样方单元位于沟谷内;这些多样的局部小生境为优势苔藓提供了多元的栖息生境。而非优势物种对生境要求高,故不同的苔藓种呈现不同的聚集斑块。综上,生境异质性和种群关系是优势物种呈大斑块聚集的主要原因。
苔类种群组的格局规模变化与总的苔藓植物格局变化基本一致,这是由于有较多重要值大的苔类植物,苔类植物种群组在样地总苔藓植物群落分布格局中占主导优势。藓类种群组的格局变化呈增加-减少-增加的趋势,充分地展现了对于生态习性相同的物种,优势物种在种内竞争中占优势(卜兆君等,2009)。藓类植物种群组的分布格局为大斑块聚集,这可能与藓类植物的生理形态有关。相关研究显示,藓类植物比苔类植物具有更多的形态特征,使其对环境的耐受力与容忍度更高,其生活型和配子形态更容易定植在开放的环境下,如暴露的岩石壁、土堆、溪流峡谷等岸边(Batista et al.,2018;Da Costa et al.,2015)。结合样地实境,藓类植物多生在沟壑和岩石壁上,其生境异质性对其影响更大,是其分布格局的主导因素。
4 结论
广东康禾自然保护区是典型的常绿阔叶林,属亚热带南缘季风气候,样地生境类型多样,沟谷、沟壑、岩石随处可见,为苔藓植物定植提供了良好的生态环境,选择苔藓植物生长分布丰富的地段设置样地进行调查,研究苔藓植物的种群分布类型及格局变化,结果显示在野外自然生境条件下,苔藓植物的分布类及规模变化主要受生境异质性和自身生物学特性的综合影响。生境异质性是导致苔藓植物集群分布的主要原因,而苔藓植物对样地沟壑、潮湿、阴郁等生态环境的偏爱和种内、种间竞争关系是导致规模变化的重要原因,优势物种在种内、种间竞争中占优势作用。
CORRALES A, DUQUE A, URIBE J, et al. 2010. Abundance and diversity patterns of terrestrial bryophyte species in secondary and planted montane forests in the northern portion of the Central Cordillera of Colombia [J]. Bryologist, 113(1): 8-21.
DA COSTA D P, DOS SANTOS N D, DE REZENDE M A, et al. 2015.Bryoflora of the Itatiaia National Park along an elevation gradient:diversity and conservation [J]. Biodiversity and Conservation, 24(9):2199-2212.
DAI X, EVANDER M. 1997. Transect-based patch size frequency analysis[J]. Journal of Vegetation Science, 8(6): 865-872.
EBERT A, DA COSTA R B, BRONDANI G E. 2016. Spatial distribution pattern of Mezilaurus itauba (Meins.) Taub. Ex mez. in a seasonal forest area of the southern Amazon, Brazil [J]. Iforest-Biogeosciences and Forestry, 9: 497-502.
FIBICH P, LEPS J, NOVOTNY V, et al. 2016. Spatial patterns of tree species distribution in New Guinea primary and secondary lowland rain forest [J]. Journal of Vegetation Science, 27(2): 328-339.
HANEWINKEL M. 2004. Spatial patterns in mixed coniferous even-aged,uneven-aged and conversion stands [J]. European Journal of Forest Research, 123(2): 139-155.
HE S Y, ZHONG Y L, SUN Y D, et al. 2017. Topography-associated thermal gradient predicts warming effects on woody plant structural diversity in a subtropical forest [J]. Scientific Reports, 7: 40387.
HE X L, HE K S, HYVONEN J. 2016. Will bryophytes survive in a warming world [J] .Perspectives in Plant Ecology Evolution and Systematics, 19: 49-60.
KREBS C. 1989. Ecological Methodology [M]. New York: HarperCollins publishers.
MANDL N A, KESSLER M, GRADSTEIN S R. 2009. Effects of environmental heterogeneity on species diversity and composition of terrestrial bryophyte assemblages in tropical montane forests of southern Ecuador [J]. Plant Ecology & Diversity, 2(3): 313-321.
MILLER H A. 1982. Bryophyte evolution and geography [J]. Biological Journal of the Linnean Society, 18: 145-196.
NGUYEN H H, URIA-DIEZ J, WIEGAND K. 2016. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam [J]. Journal of Vegetation Science, 27(2):318-327.
BATISTA W V S M, PÔRTO K C, SANTOS N D D. 2018. Distribution,ecology, and reproduction of bryophytes in a humid enclave in the semiarid region of northeastern Brazil [J]. Acta Botanica Brasilica,32(2): 303-313.
SONG L, LU H Z, XU X L, et al. 2016. Organic nitrogen uptake is a significant contributor to nitrogen economy of subtropical epiphytic bryophytes [J]. Scientific Reports, 6: 30408.
SONG L, MA W Z, YAO Y L, et al. 2015. Bole bryophyte diversity and distribution patterns along three altitudinal gradients in Yunnan, China[J]. Journal of Vegetation Science, 26(3): 576-587.
SZMYT J. 2014. Spatial statistics in ecological analysis: from indices to functions [J]. Silva Fennica, 48(1): 93-109.
USHER M B. 1983. Pattern in the Simple Moss-Turf Communities of the Sub-Antarctic and Maritime Antarctic [J]. Journal of Ecology, 71(3):945-958.
YANG F, ZHOU Y. 2017. Quantifying spatial scale of positive and negative terrains pattern at watershed-scale:Case in soil and water conservation region on Loess Plateau [J]. Journal of Mountain Science, 14(8):1642-1654.
艾应伟, 刘浩, 李伟, 等. 2010. 苔藓植物资源保护利用中的生态环境效应研究进展[J]. 生态环境学报, 19(1): 227-232.
卜兆君, 陈旭, 姜丽红, 等. 2009. 苔藓植物相互作用的研究进展[J]. 应用生态学报, 20(2): 460-466.
陈勤, 孙冲, 方炎明. 2013. 苔藓植物的生态环境指示作用[J]. 世界林业研究, 26(2): 19-23.
陈云, 冯佳伟, 牛帅, 等. 2017. 小秦岭自然保护区苔藓植物群落数量分类、排序及多样性垂直格局[J]. 生态学报, 37(8): 2653-2664.
董秋莲, 李毅, 单立山, 等. 2013. 张掖市龙首山红砂种群结构和分布格局研究[J]. 水土保持通报, 33(4): 284-288.
郭磊, 韦博良, 胡金涛, 等. 2016. 基于两个不同资源轴上苔藓植物生态位分析[J]. 生态学报, 21(37): 7266-7276.
郭连金. 2014. 濒危植物香果树幼苗空间格局及数量动态研究[J]. 西北植物学报, 34(9): 1887-1893.
贾成朕, 苗百岭, 姜威, 等. 2017. 阿拉善典型荒漠群落种群空间格局及种间关联的尺度依赖性[J]. 内蒙古大学学报(自然版), 48(2):142-148.
贾渝, 何思. 2013. 中国生物物种名录: 苔藓植物[M]. 北京: 科学出版社.
姜苹红, 罗远玲, 彭克俭, 等. 2015. 苔藓植物运用于大气重金属污染监测的研究进展[J]. 环境污染与防治, 37(7): 82-87.
兰国玉, 雷瑞德. 2003. 植物种群空间分布格局研究方法概述[J]. 西北林学院学报, 18(2): 17-21.
李海涛. 1995. 植物种群分布格局研究概况[J]. 植物学报, 12(2): 19-26.
李素清, 李斌, 张金屯. 2006. 基于 DCA-TTLQV 的云顶山亚高山草甸群落格局分析[J]. 草业学报, 15(6): 44-48.
李素清, 武冬梅, 王涛, 等. 2011. 山西长治湿地草本植物优势种群和群落的空间格局分析[J]. 草业学报, 20(3): 43-50.
李亚, 胡小柯, 魏怀东, 等. 2017. 腾格里沙漠南缘天然群落主要种群空间分布格局研究[J]. 西北林学院学报, 32(2): 67-72.
刘蔚秋, 雷纯义, 戴小华. 2007. 广东黑石顶森林苔藓植物群落特征初探[J]. 热带亚热带植物学报, 15(6): 538-544.
马钦彦. 2009. 负二项式分布及种群格局检验分析[J]. 北京林业大学学报, 31(3): 1-5.
梅军林, 庄枫红, 马姜明, 等. 2017. 桂林喀斯特地区克隆生长红背山麻杆种群的点格局分析[J]. 生态学报, 37(9): 3164-3171.
田维莉, 孙守琴. 2011. 苔藓植物生态功能研究新进展[J]. 生态学杂志,30(6): 1265-1269.
王继和, 靳虎甲, 马全林, 等. 2010. 干旱区油蒿种群结构和分布格局分析[J]. 中国沙漠, 30(3): 534-538.
王利松, 贾渝, 张宪春, 等. 2015. 中国高等植物多样性[J]. 生物多样性,23(2): 217-224.
王中生, 安树青, 方炎明. 2003. 苔藓植物生殖生态学研究[J]. 生态学报, 23(11): 2444-2452.
吴德邻. 2013. 广东苔藓志[M]. 广州: 广东科技出版社.
许宁, 韦博良, 胡金涛, 等. 2017. 宝天曼自然保护区树附生苔藓空间格局分析[J]. 河南农业大学学报, 51(2): 230-236.
张金屯. 2005. 芦芽山亚高山草甸优势种群和群落的二维格局分析[J].生态学报, 25(6): 1264-1268.
张金屯. 2011. 数量生态学[M]. 第2版. 北京: 科学出版社.
张金屯, 孟东平. 2006. 芦芽山油松-辽东栎林优势树种空间分布格局研究[J]. 西北植物学报, 26(8): 1682-1685.
张瑾, 陈文业, 张继强, 等. 2013. 甘肃敦煌西湖荒漠-湿地生态系统优势植物种群分布格局及种间关联性[J]. 中国沙漠, 33(2): 349-357.
张维, 李海燕, 赖晓辉, 等. 2016. 新疆天山峡谷不同坡向野核桃种群分布格局[J]. 应用生态学报, 27(10): 3105-3113.
张旭, 李家湘, 喻勋林, 等. 2015. 湖南大围山杜鹃灌丛木本植物种群空间格局[J]. 生态学杂志, 34(11): 3034-3039.
张元明, 曹同, 潘伯荣. 2002. 干旱与半干旱地区苔藓植物生态学研究综述[J]. 生态学报, 22(7): 1129-1134.
赵峰侠, 尹林克. 2007. 荒漠内陆河岸胡杨和多枝柽柳幼苗种群空间分布格局及种间关联性[J]. 生态学杂志, 26(7): 972-977.
猜你喜欢
中国卫生统计(2022年4期)2022-10-12
今日农业(2022年15期)2022-09-20
——平衡不完全区组设计定量资料一元方差分析
四川精神卫生(2022年2期)2022-05-09
天津中医药大学学报(2020年1期)2020-03-03
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
红土地(2018年7期)2018-09-26
小说月刊(2015年12期)2015-04-23
科技视界(2014年5期)2014-12-27
当代畜禽养殖业(2014年10期)2014-02-27
