
具饱和发生率的被修正HIV传染病模型的全局稳定性*
2018-06-07 03:02杨俊仙王雷宏
中山大学学报(自然科学版)(中英文) 2018年3期
杨俊仙,王雷宏
(1.安徽农业大学理学院,安徽合肥230036;2.安徽农业大学林学与园林学院,安徽合肥230036)
艾滋病的病原体是人类免疫缺陷病毒(Human Immunodeficiency Virus,简称 HIV),主要感染人体免疫系统细胞CD4+T,可引起细胞计数大幅度下降,导致人体免疫缺陷,严重影响患者抵御感染的能力[1]。
目前,数学建模已经成为分析和控制传染病传播的重要工具。近年来,对艾滋病在数学建模方面的研究已取得很大进展[2-3]。对模型的改进是为了更好地分析和解释病情,并预测和控制发病率。最初的HIV/AIDS模型是由Nowak和Perelson等[4-5]提出的,该模型被广泛应用于艾滋病感染动力学:

其中T(t),I(t),V(t)分别表示t时刻未感染 CD4+T细胞个数、已感染CD4+T细胞个数和病毒载量(HIV)。参数A表示未感染细胞的固有生成率,β表示病毒感染率,d1,d2,d3分别表示未感染细胞、已感染细胞和病毒的死亡率,k表示病毒复制率。A,β,d1,d2,d3,k均为正数。
在20世纪90年代,有一个关于HIV RNA转录成DNA的讨论:当HIV病毒进入CD4+T细胞后,HIV病毒可能并未完全反转录成DNA。在病毒基因组被整合到淋巴细胞基因组之前,一部分处于潜伏期的感染细胞可以恢复到未被感染的状态[6]。最近,已有一些数学模型建立在这样假设(部分感染细胞可恢复到未被感染状态)的基础上[7-8]。其中 Srivastava和 Chandra讨论了如下模型[7]:

其中变量T(t),I(t),V(t)和参数A,β,d1,d2,d3,k与模型(1)具有相同的意义,这里参数p(p>0)表示处于潜伏期感染细胞的恢复率。
在模型(1)和(2)中,感染率 βT(t)V(t)被假设为在未感染细胞个数T(t)和病毒载量V(t)之间是双线性的。然而,实际发生率可能不是完全双线性的。Sun等[9]提出了如下模型:

此处用的是非线性发生率即发生率不再是未感染细胞个数T(t)和病毒载量V(t)的双线性关系。
在本文,我们提出了具有饱和发生率的被修正HIV传染病模型:

假设参数α>0且d1≤d2。
由系统(4)的前2个方程得,

再由系统(4)的第3个方程得

于是

因此系统(4)的正向不变集为

1 平衡点的局部稳定性
系统(4)总存在无病平衡点E0(T0,0,0),其中。定义基本再生数

易证:当R0>1时,系统(4)存在唯一的正平衡点E*(T*,I*,V*),其中

且正平衡点E*(T*,I*,V*)在Ω的内部:


因此,只需要在Ω0上考虑E*(T*,I*,V*)的稳定性。
1.1 无病平衡点E0的局部稳定性
定理1当R0<1时,系统(4)的无病平衡点E0(T0,0,0)是局部渐近稳定的;当R0>1时,E0(T0,0,0)是不稳定的。
证明系统(4)在无病平衡点E0(T0,0,0)处的线性化系统的特征方程为

显然,方程(9)总有负实根λ=-d1,其余的根取决于方程

整理方程(10)得

当,方程(11)所有的根均有负实部,因此无病平衡点E0是局部渐近稳定的。当R0>1时,方程(11)有一个正实根,因此无病平衡点E0是不稳定的。
1.2 正平衡点E*(T*,I*,V*)的局部渐近稳定性
定理2当R0>1时,系统(4)的正平衡点E*(T*,I*,V*)是局部渐近稳定的。
证明系统(4)在正平衡点E*(T*,I*,V*)的线性化系统的特征方程为

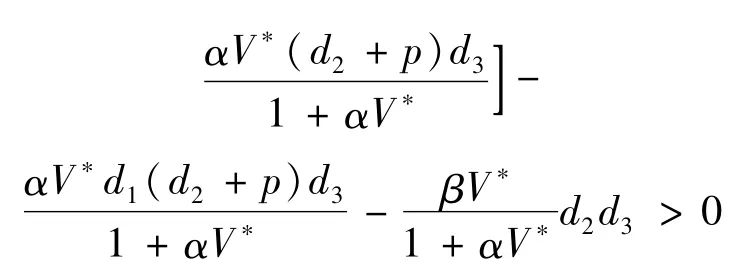
由Routh-Hurwitz判别准则知,正平衡点E*(T*,I*,V*)是局部渐近稳定的。
2 平衡点的全局稳定性
2.1 无病平衡点E0的全局渐近稳定性
定理3当R0<1时,系统(4)的无病平衡点E0(T0,0,0)在Ω内是全局渐近稳定的。
证明令(T(t),I(t),V(t))是系统(4)的任一正解,定义Lyapunov函数:

计算V1(t)沿系统(4)的全导数:

当R<1时,当且仅当V(t)=0时。因此,在 Ω内的最大正向不变集为

于是有极限方程:

定义Lyapunov函数:

其中T0=。计算V2(t)沿系统(13)的全导数:

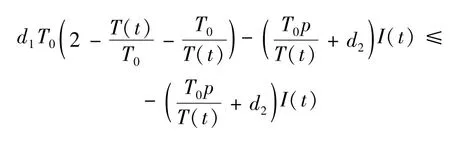
其中当且仅当T(t)=T0,I(t)因此的正向不变集为:

由 LaSalle不变集原理知,E0(T0,0,0)是全局渐近稳定的。
2.2 正平衡点E*的全局渐近稳定性
设开集是C1函数。考虑微分方程

设x(t,x0)代表方程(15)满足条件x(0)=x0的解。令是系统(15)的一个平衡点,Li等[10]作了下面两个基本假设:
(H1)方程(15)在Γ内存在一个紧吸引子集K⊂Γ;
(H2)方程(15)在Γ内有唯一平衡点∈Γ。
给出了如下结论:
引理1[10]若下列条件成立:
(i)假设(H1)和(H2)成立;
(ii)方程(15)满足 Poincare′-Bendixson性质;
(iii)对系统(15)具有p(0)∈D的每一个周期解x=p(t),系统(15)关于p(t)的二阶复合矩阵

是渐近稳定的,其中是f的Jacobian矩阵的第二加性复合矩阵;

则系统(15)的唯一平衡点在Γ内是全局渐近稳定的。
定理4当R0>1时,系统(4)的正平衡点E*(T*,I*,V*)在Ω0内是全局渐近稳定的。
证明根据引理,逐条验证四个条件:
(i)条件(H1)等价于系统(4)的一致持久性[11]。由式(8)知,Ω0是有界的,所以对于系统(4)的一致持久性的充分必要条件等价于平衡点E0是不稳定的 。定理1已证:当R0>1时,E0是不稳定的,因此系统(4)一致持久的,从而(H1)成立。
同时,由于E*(T*,I*,V*)是系统(4)在Ω0内的唯一平衡点,因此(H2)成立。
(ii)系统(4)的Jacobian矩阵为

取H=diag(1,-1,1),则

显然,HJH的非对角线元素非正,系统(4)满足Poincare′-Bendixson性质,因此引理1的条件(ii)成立。
(iii)令p(t)=(T(t),I(t),V(t))是系 统(4)在Ω0内的任意一个周期解,则系统(4)的Jacobian矩阵的第二加性复合矩阵为

沿系统(4)任一周期解(T(t),I(t),V(t))的二阶复合系统为

考虑Lyapunov函数:
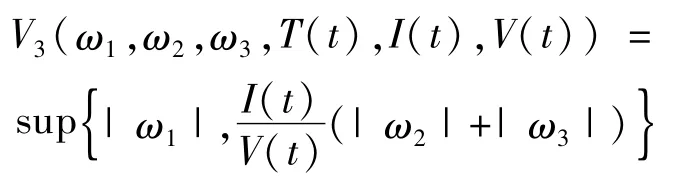
由一致持久性知,周期解p(t)=(T(t),I(t),V(t))与边界∂Ω有一定的距离,故存在常数μ>0,使得T(t)>μ,I(t)>μ,V(t)>μ。并结合式(8)中的得

对全部(ω1,ω2,ω3)∈ R3及(T(t),I(t),V(t)),计算V3的右导数。注意到


于是由式(22),式(25)和Gronwall不等式得

由上式知故由式(19)知,当t即二阶复合系统(18)是渐近稳定的。这样就验证了引理1中的条件(iii)。
(iv)由式(13)可得,

由于J(E*)是3×3矩阵,即n=3,于是

验证了引理1的条件(iv)。
因此,当R0>1时,由引理得,系统(4)唯一的正平衡点E*(T*,I*,V*)在Ω0内是全局渐近稳定的。
3 数值模拟
在系统(4)中,令参数A=5,k=10,β=0.000 2,α=0.5,p=0.5,d1=0.1,d2=0.5,d3=0.3,显然R0=<1,此时系统存在一个无病平衡点E0(50,0,0)。由定理3可知,E0是全局渐近稳定的,数值模拟结果验证了上述结论(见图1)。

图1 R0 <1时,E0(T0,0,0)的稳定性Fig.1 When R0 <1,the stability of E0(T0,0,0)
若令参数此时系统(4)有唯一的正平衡点E*(41.92,1.62,53.89)。根据定理4可知,E*是全局渐近稳定的,数值模拟的结果与文中结论一致(见图2)。

图2 R0>1时,E*(T*,I*,V*)的稳定性Fig.2 When R0 >1,the stability of E*(T*,I*,V*)
[1]WANGL C,LI M Y.Mathematical analysis of the global dynamics of a model for HIV infection of CD4+T cells[J].Mathematical Biosciences,2006,200(1):44-57.
[2]杨俊仙,吴元翠,闫萍.一类具时滞与饱和发生率的HIV-1传染病模型的全局稳定性[J].中山大学学报(自然科学版),2016,55(4):26-29,38.YANG J X,WU Y C,YAN P.Global stability of a HIV-1 epidemic model with time delay and saturation incidence rate[J].Acta Scientirum Naturalium Universitatis Sunyatseni,2016,55(4):26-29,38.
[3]冯依虎,陈贤峰,莫嘉琪.一类免疫缺陷病毒传播的非线性动力学系统[J].中山大学学报(自然科学版),2017,56(5):60-63.FENG Y H,CHEN X F,MOJQ.A class of nonlinear dynamic system of human groups for HIV transmission[J].Acta Scientirum Naturalium Universitatis Sunyatseni,2017,56(5):60-63.
[4]NOWAK M A,BANGHAM CR M.Population dynamics of immune responses to persistent viruses[J].Science,1996,272(5258):74-79.
[5]PERELSON A S,NELSON P W.Mathematical models of HIV dynamics in vivo[J].SIAM Review,1999,41(1):3-44.
[6]ESSUNGER P,PERELSON A S.Modeling HIV infection of CD4+T cell subpopulations[J].Journal of Theoretical Biology,1994,170(4):367-391.
[7]SRIVASTAVA PK,CHANDRA P.Modeling the dynamics of HIV and CD4+T cells during primary infection[J].Nonlinear Analysis: Real World Applications,2010,11(2):612-618.
[8]BUONOMO B,VARGAS-DE-LEON C.Global stability for an HIV-1 infection model including an eclipse stage of infected cells[J].Journal of Mathematical Analysis and Applications,2012,385(2):709-720.
[9]SUN Q L,MIN L Q.Dynamics analysis and simulation of a modified HIV infection model with a saturated infection rate[J].Computational and Mathematical Methods in Medicine,2014,2014(5258):145162.
[10]MICHAEL Y L,WANG L C.Global stability in some SEIR epidemic models[J].Mathematical Approaches for Emerging and Reemerging Infectious Diseases:Models,Methods,and Theory,2002,126:295-311.
[11]WALTMAN P.A brief survey of persistence[C]∥Proceedings of a Conference in Honor of Kenneth Cooke,1991:31-40.
[12]LI M Y,GRAEF JR,WANG L,et al.Global dynamics of a SEIR model with varying total population size[J].Mathematical Biosciences,1999,160(2):191-213.
猜你喜欢
北京航空航天大学学报(2022年8期)2022-08-31
中等数学(2022年6期)2022-08-29
吉首大学学报(自然科学版)(2022年1期)2022-08-10
中学生数理化(高中版.高考数学)(2021年11期)2021-12-21
闽南师范大学学报(自然科学版)(2021年3期)2021-10-19
校园英语·上旬(2019年6期)2019-10-09
金桥(2018年4期)2018-09-26
北京航空航天大学学报(2016年9期)2016-11-16
IT时代周刊(2015年7期)2015-11-11
