
浙江省蜡梅野生种群种实表型性状变异及其与土壤相关因子的关系
2018-06-06 03:15:15杜会聪蒋雅婷段国敏
植物资源与环境学报 2018年2期
杜会聪,蒋雅婷,张 莹,田 敏,段国敏
(中国林业科学研究院亚热带林业研究所,浙江 杭州 311400)
植物的种实普遍存在形态变异,其表型性状主要受遗传因子控制[1],且在长期的生殖隔离、自然选择和人工选择等[2-3]作用下,产生了丰富的变异。种实表型性状变异程度不仅决定植物的扩散能力,对种子萌发、幼苗定居和存活、种群更新和群落演替等[4-7]也有重要的生态学意义。学者对思茅松〔Pinuskesiyavar.langbianensis(A. Chev.) Gaussen〕[8]、秦岭冷杉(AbieschensiensisTiegh.)[2]、夏蜡梅(CalycanthuschinensisCheng et S. Y. Chang)[9-10]和无患子(SapindusmukorossiGaertn.)[11]等植物的种实表型多样性进行了研究,表明利用表型性状研究种群遗传多样性是重要且有效的途径之一[12]。
蜡梅〔Chimonanthuspraecox(Linn.) Link〕隶属于蜡梅科(Calycanthaceae)蜡梅属(ChimonanthusLindl.),为中国特有的传统名花和经济树种[13]。蜡梅冬季开花且芳香宜人,观赏价值极高,现已作为重要的园林绿化树种在全球不同区域广泛栽培,且被用于芳香油的开发生产[14-15]。蜡梅广泛分布于安徽、福建、贵州、河南、湖北、湖南、江苏、江西、陕西、山东、四川、云南和浙江等地,是第四纪孑遗植物。野生的蜡梅是培育蜡梅优良品种的重要基因资源,但近年来由于生境破坏及人为影响使其野生资源遭到严重破坏,因此,有必要对蜡梅野生种群的遗传多样性和变异状况进行深入研究。
浙江省是华东地区蜡梅野生种群最主要的分布区,在蜡梅的区系分布和种质资源等方面占有重要地位[16-17]。作者在全面调查浙江省蜡梅野生种群分布状况的基础上,选择4个主要分布区域,对种群间和种群内蜡梅种实表型性状的变异规律进行分析,并探究其与土壤相关因子的关联度,为蜡梅优良种源的筛选和保护提供基础数据。
1 研究区概况和研究方法
1.1 研究区概况
浙江省蜡梅野生种群主要分布于杭州富阳的龙王坎、碧东山和五尖山以及杭州临安的方山境内。各种群的自然概况见表1。
浙江省蜡梅野生分布区属亚热带季风气候,年均温16.6 ℃,年均降水量1 457.8 mm,年均空气相对湿度82%,年雨日152.3 d;土壤主要为黑色石灰土。富阳龙王坎种群(P1)的伴生植物主要有柏木(CupressusfunebrisEndl.)、化香树(PlatycaryastrobilaceaSieb. et Zucc.)和盐肤木(RhuschinensisMill.)等;种群外貌藤蔓交错,藤蔓植物主要有紫藤〔Wisteriasinensis(Sims) Sweet〕、三叶木通〔Akebiatrifoliata(Thunb.) Koidz.〕和常春油麻藤(MucunasempervirensHemsl.)等。富阳碧东山种群(P2)和临安方山种群(P4)的伴生植物主要有化香树、枇杷〔Eriobotryajaponica(Thunb.) Lindl.〕、浙江楠(PhoebechekiangensisC. B. Shang)、桂花〔Osmanthusfragrans(Thunb.) Lour.〕、紫楠〔Phoebesheareri(Hemsl.) Gamble〕、糙叶树〔Aphanantheaspera(Thunb.) Planch.〕、青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕、枳椇(HoveniaacerbaLindl.)、络石〔Trachelospermumjasminoides(Lindl.) Lem.〕和野菊(ChrysanthemumindicumLinn.)等。富阳五尖山种群(P3)的蜡梅个体数量较多且分布相对集中,该种群以蜡梅为优势种,其他灌木和草本种类均较少,群落盖度极低。
表1浙江省4个蜡梅野生种群的自然概况
Table1NaturalstatusoffourwildpopulationsofChimonanthuspraecox(Linn.)LinkinZhejiangProvince
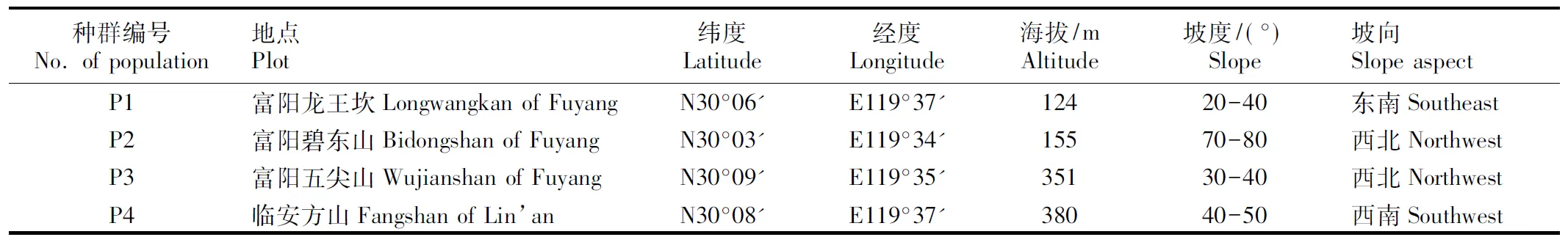
种群编号No. of population地点Plot纬度Latitude经度Longitude海拔/mAltitude坡度/(°)Slope坡向Slope aspectP1富阳龙王坎Longwangkan of Fuy-angN30°06'E119°37'12420-40东南SoutheastP2富阳碧东山Bidongshan of FuyangN30°03'E119°34'15570-80西北NorthwestP3富阳五尖山Wujianshan of FuyangN30°09'E119°35'35130-40西北NorthwestP4临安方山Fangshan of Lin’anN30°08'E119°37'38040-50西南Southwest
1.2 方法
1.2.1 种实采集和性状测定 在2017年7月中旬蜡梅果实成熟期,在各种群内随机选取株龄10~20 a、生长正常、无严重缺陷和病虫害的成年个体10~20株作为样株,株间距保持20 m以上,以最大限度排除亲缘性。选择各样株向阳侧枝条,在枝条中部随机采集果实10~20个,按样株分装后带回实验室测量种子和果实的相关指标。
1.2.2 土壤样品采集及其相关因子测定 在各种群内按照“S”型曲线选取5株样株,在每一样株根系附近挖取距地表0~20 cm的土层样品,混合后自然风干、过筛(100目),备用。分别按照LY/T 1239—1999、LY/T 1228—2015、LY/T 1232—2015、LY/T 1234—2015、LY/T 1270—1999、LY/T 1271—1999、LY/T 1262—1999、LY/T 1261—1999、LY/T 1260—1999、LY/T 1263—1999和LY/T 1246—1999中的方法测定土壤的pH值及N、P、K、Ca、Mg、Fe、Zn、Cu、Mn和Na含量,测定工作由国家林业局经济林产品质量检验检测中心(杭州)完成。
1.3 数据处理和分析
采用EXCEL 2007、SPSS 20.0和SAS 11.5软件进行数据统计和差异显著性分析;采用UPGMA法进行Q型聚类分析;用表型分化系数衡量种群间表型分化程度;用变异系数衡量种群内各表型性状的变异水平;用主成分分析法获得各表型性状对蜡梅种群种实变异的贡献率。
采用线性模型“Yijk=μ+Pi+Ti(j)+e(ij)k”对蜡梅表型性状变异进行巢式方差分析[8]。式中,Yijk为第i个种群第j个个体第k个观测值,μ为总均值,Pi为第i个种群的效应值(固定),Ti(j)为第i个种群第j个个体的效应值(随机),e(ij)k为实验误差。
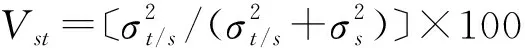

以蜡梅种实表型性状数据组成母数列、土壤元素含量各自的数列为子数列,构建数据矩阵并均值化后进行无量纲化处理,参照文献[19-20]对蜡梅种实表型性状变异与土壤相关因子进行灰色关联度分析。
2 结果和分析
2.1 种实表型性状的比较及其变异分析
分布于浙江省的4个蜡梅野生种群种实11个表型性状的均值及各表型性状的变异系数和表型分化系数均见表2。
文化工作者,是为我们的灵魂服务的。我们肩负了将党和国家的文化惠民政策贯彻落实到基层的使命,因此,我们的责任和义务十分重要。这一观点,我们在调研采访中得到了验证。
2.1.1 种实表型性状的比较 由表2可见:富阳龙王坎种群的种形系数和单果果壳质量均最大;富阳碧东山种群的种子长、种子宽、种子千粒质量、果实长、果实宽、果形系数、果口直径、单果质量和单果种子数均最大,其中,果口直径、单果质量和单果种子数均极显著高于其他种群;富阳五尖山种群的果形系数和种子千粒质量均最小;临安方山种群的种子长、种形系数、果实长、果实宽、单果质量、单果种子数和单果果壳质量均最小,其中,种子长、种形系数、果实宽和单果质量均极显著低于其他种群。11个表型性状中,仅种子千粒质量在种群间无显著差异。
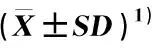
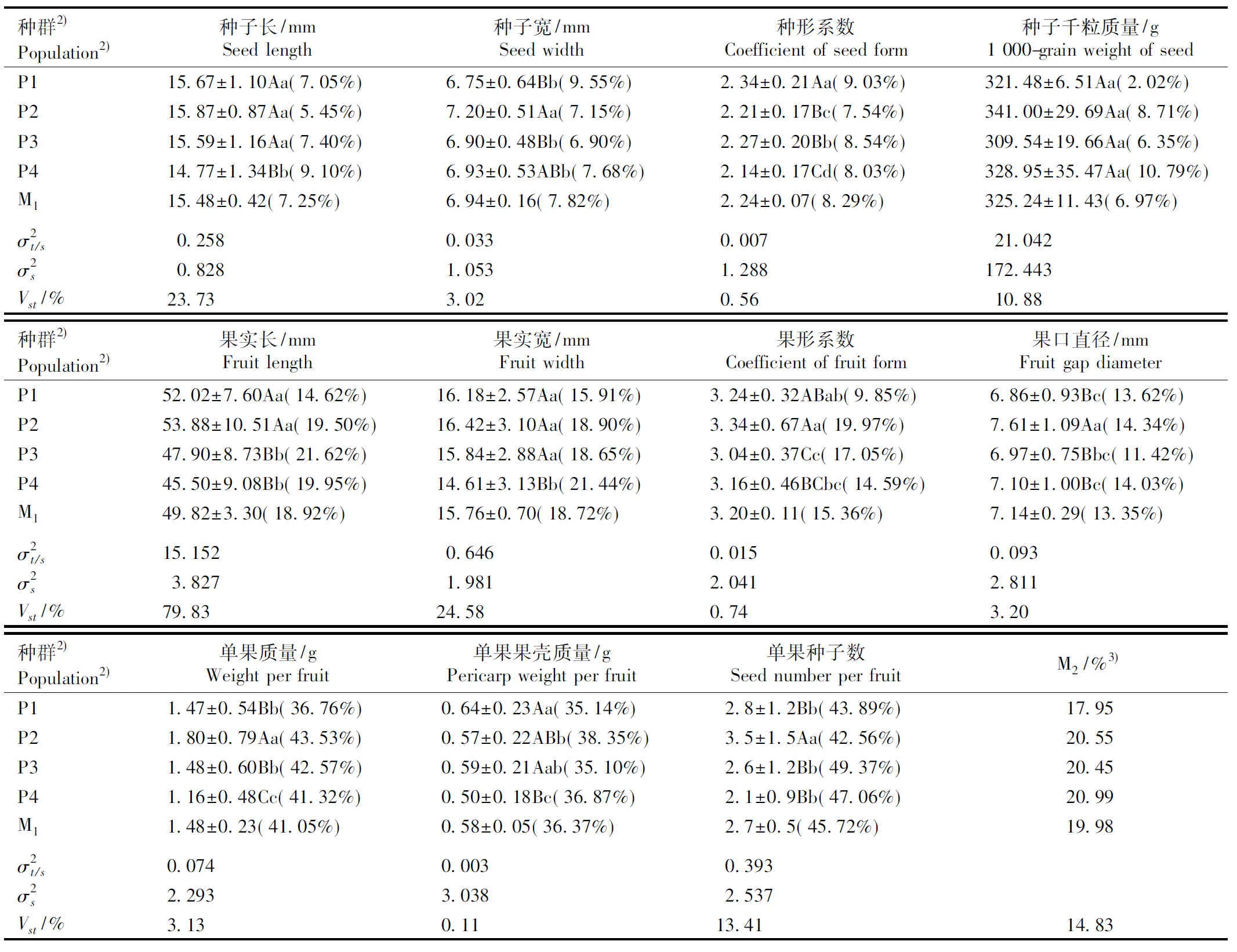
种群2)Popula-tion2)种子长/mmSeed length种子宽/mmSeed width种形系数Coefficient of seed form种子千粒质量/g1 000-grain weight of seedP115.67±1.10Aa(7.05%)6.75±0.64Bb(9.55%)2.34±0.21Aa(9.03%)321.48±6.51Aa(2.02%)P215.87±0.87Aa(5.45%)7.20±0.51Aa(7.15%)2.21±0.17Bc(7.54%)341.00±29.69Aa(8.71%)P315.59±1.16Aa(7.40%)6.90±0.48Bb(6.90%)2.27±0.20Bb(8.54%)309.54±19.66Aa(6.35%)P414.77±1.34Bb(9.10%)6.93±0.53ABb(7.68%)2.14±0.17Cd(8.03%)328.95±35.47Aa(10.79%)M115.48±0.42(7.25%)6.94±0.16(7.82%)2.24±0.07(8.29%)325.24±11.43(6.97%)σ2t/s0.2580.0330.00721.042σ2s0.8281.0531.288172.443Vst/%23.733.020.5610.88种群2)Popula-tion2)果实长/mmFruit length果实宽/mmFruit width果形系数Coefficient of fruit form果口直径/mmFruit gap diameter P152.02±7.60Aa(14.62%)16.18±2.57Aa(15.91%)3.24±0.32ABab(9.85%)6.86±0.93Bc(13.62%)P253.88±10.51Aa(19.50%)16.42±3.10Aa(18.90%)3.34±0.67Aa(19.97%)7.61±1.09Aa(14.34%)P347.90±8.73Bb(21.62%)15.84±2.88Aa(18.65%)3.04±0.37Cc(17.05%)6.97±0.75Bbc(11.42%)P445.50±9.08Bb(19.95%)14.61±3.13Bb(21.44%)3.16±0.46BCbc(14.59%)7.10±1.00Bc(14.03%)M149.82±3.30(18.92%)15.76±0.70(18.72%)3.20±0.11(15.36%)7.14±0.29(13.35%)σ2t/s15.1520.6460.0150.093σ2s3.8271.9812.0412.811Vst/%79.8324.580.743.20种群2)Popula-tion2)单果质量/gWeight per fruit单果果壳质量/gPericarp weight per fruit单果种子数Seed number per fruit M2/%3)P11.47±0.54Bb(36.76%)0.64±0.23Aa(35.14%)2.8±1.2Bb(43.89%)17.95P21.80±0.79Aa(43.53%)0.57±0.22ABb(38.35%)3.5±1.5Aa(42.56%)20.55P31.48±0.60Bb(42.57%)0.59±0.21Aab(35.10%)2.6±1.2Bb(49.37%)20.45P41.16±0.48Cc(41.32%)0.50±0.18Bc(36.87%)2.1±0.9Bb(47.06%)20.99M11.48±0.23(41.05%)0.58±0.05(36.37%)2.7±0.5(45.72%)19.98σ2t/s0.0740.0030.393σ2s2.2933.0382.537Vst/%3.130.1113.4114.83
1)同列中不同的小写和大写字母分别表示不同种群间同一表型性状在0.05和0.01水平上差异显著 Different lowercases and capitals in the same column indicate the significant differences in the same phenotypic trait among different populations at 0.05 and 0.01 levels,respectively. 括号内的百分数为变异系数 The percentages in brackets are coefficients of variation.

3)M2:各种群所有表型性状变异系数的均值 Average of coefficients of all phenotypic trait variations of each population.
2.1.2 种实表型性状的变异分析 由表2还可见:各表型性状的变异程度有明显差异。各表型性状的变异程度由大到小依次为单果种子数、单果质量、单果果壳质量、果实长、果实宽、果形系数、果口直径、种形系数、种子宽、种子长、种子千粒质量。单果种子数、单果质量和单果果壳质量的变异系数明显大于其他表型性状,说明这3个表型性状易受环境影响。其中,单果种子数的变异系数最大,为45.72%,稳定性最差;种子长、种子宽、种形系数和种子千粒质量的变异系数相对较小,说明这4个表型性状较为稳定,其中种子千粒质量变异系数最小(6.97%),稳定性最好。
从11个表型性状的变异系数均值看,富阳碧东山、富阳五尖山和临安方山3个种群的均值相近(约20%),而富阳龙王坎种群的均值仅为17.95%,明显小于前3个种群,表明不同种群间11个表型性状的变异系数均值也存在一定差异。
从巢式方差分析结果看,11个表型性状的表型分化系数为0.11%~79.83%,差异明显。而种群间的表型分化系数均值仅为14.83%,说明种群内的表型分化系数均值达到了85.17%,表明蜡梅野生种群种实表型性状变异以种群内变异为主。
2.2 种实表型性状的相关性和主成分分析
2.2.1 相关性分析 对供试4个蜡梅野生种群种实11个表型性状间的相关性进行分析,结果见表3。结果显示:蜡梅种子和果实性状间存在不同程度的相关性。除种形系数外,果实长与其他表型性状均呈显著(P<0.05)或极显著(P<0.01)正相关;种形系数仅与种子宽呈极显著负相关,相关系数为-0.546,与其他表型性状均无显著相关性;种子千粒质量仅与果实长呈显著正相关,相关系数为0.507,与其他表型性状均无显著相关性。总体上看,11个表型性状中,种形系数和种子千粒质量与其他表型性状的相关性均较小;果实长、果实宽、单果质量、单果种子数、果口直径和单果果壳质量6个果实表型性状间均存在极显著相关性。
表3浙江省蜡梅野生种群种实表型性状的相关系数1)
Table3CorrelationcoefficientamongphenotypictraitsofseedandfruitofwildpopulationsofChimonanthuspraecox(Linn.)LinkinZhejiangProvince1)

表型性状Phenotypic trait不同表型性状间的相关系数 Correlation coefficient among different phenotypic traitsSLSWCSFGWSFLFWCFFFGDWFPWFSNFSL1.000SW0.367**1.000CSF-0.013-0.546**1.000GWS0.2470.423-0.1741.000FL0.287**0.170**-0.0120.507*1.000FW0.308**0.137*-0.0110.2150.722**1.000CFF-0.0500.040-0.0060.2390.363**-0.365**1.000FGD0.116*0.178**-0.0600.2470.463**0.604**-0.196**1.000WF0.379**0.229**-0.0740.3560.759**0.840**-0.1180.564**1.000PWF0.314**0.137*-0.0550.3570.698**0.818**-0.167**0.533**0.882**1.000SNF0.230**0.173**-0.0700.1360.686**0.736**-0.0620.482**0.895**0.705**1.000
1)SL:种子长Seed length;SW:种子宽Seed width;CSF:种形系数Coefficient of seed form;GWS:种子千粒质量1 000-grain weight of seed;FL:果实长Fruit length;FW:果实宽Fruit width;CFF:果形系数Coefficient of fruit form;FGD:果口直径 Fruit gap diameter;WF:单果质量Weight per fruit;PWF:单果果壳质量 Pericarp weight per fruit;SNF:单果种子数Seed number per fruit. *:P<0.05;** :P<0.01.
2.2.2 主成分分析 对供试4个蜡梅野生种群种实11个表型性状进行主成分分析,结果见表4。结果显示:第1、第2、第3和第4主成分的贡献率分别为40.011%、19.992%、16.486%和10.167%,累计贡献率达到86.656%,表明这4个主成分可以代表11个表型性状的绝大部分信息。
在第1主成分中,主要作用因子是单果果壳质量、果实宽、单果质量和果实长,特征向量分别为0.957、0.863、0.828和0.824;在第2主成分中,主要作用因子是种子长,特征向量为0.838;在第3主成分中,主要作用因子是种形系数,特征向量为0.807;在第4主成分中,主要作用因子是种子千粒质量,特征向量为0.738。由此可见,单果果壳质量、果实宽、单果质量、果实长、种子长、种形系数和种子千粒质量对蜡梅野生种群种实表型性状变异有主要作用。
2.3 种实表型性状与土壤相关因子的灰色关联度分析
2.3.1 土壤相关因子的比较 供试4个蜡梅野生种群土壤相关因子的测定结果见表5。结果显示:土壤大量元素中,富阳碧东山种群土壤中N和P含量明显高于其他种群,而富阳五尖山种群土壤中N和K含量明显小于其他种群。土壤微量元素中,Zn和Fe含量在4个种群间有明显差异,其中,土壤Zn含量以临安方山种群最高,富阳龙王坎种群最低,前者是后者的5.70倍;土壤Fe含量以富阳碧东山种群最高,富阳五尖山种群最低,前者是后者的4.28倍。另外,4个种群的土壤均呈弱碱性,土壤pH值为pH 7.56~pH 8.06。
表4浙江省蜡梅野生种群种实表型性状的主成分分析结果1)
Table4ResultofprincipalcomponentanalysisonphenotypictraitsofseedandfruitofwildpopulationsofChimonanthuspraecox(Linn.)LinkinZhejiangProvince1)

主成分Principal component特征向量 EigenvectorSLSWCSFGWSFLFWCFFFGDWFPWFSNFECR/%CCR/%1-0.2610.137-0.375 0.4080.8240.863-0.195 0.7890.8280.9570.5684.40140.01140.01120.8380.6110.2530.2430.362-0.2720.797-0.1530.1860.0090.3192.19919.99260.00330.251-0.6260.807-0.082-0.0050.193-0.313-0.4340.2670.0770.5481.81316.48676.4894-0.071-0.3850.3050.7380.1790.0360.2220.175-0.3070.025-0.3461.11810.16786.656
1)SL:种子长Seed length;SW:种子宽Seed width;CSF:种形系数Coefficient of seed form;GWS:种子千粒质量1 000-grain weight of seed;FL:果实长Fruit length;FW:果实宽Fruit width;CFF:果形系数Coefficient of fruit form;FGD:果口直径 Fruit gap diameter;WF:单果质量Weight per fruit;PWF:单果果壳质量 Pericarp weight per fruit;SNF:单果种子数Seed number per fruit;E:特征值Eigenvalue;CR:贡献率Contribution rate;CCR:累计贡献率Cumulative contribution rate.
表5浙江省蜡梅野生种群土壤相关因子的比较
Table5ComparisononsoilrelatedfactorsofwildpopulationsofChimonanthuspraecox(Linn.)LinkinZhejiangProvince

种群Population土壤元素含量/mg·kg-1 Soil element contentNPKCaMgFeZnCuMnNa土壤pH值Soil pH value富阳龙王坎Longwangkan of Fuyang2133.394.73 5041 2141684.22.771814.97.79富阳碧东山Bidongshan of Fuyang36911.799.44 9955662536.23.954915.57.56富阳五尖山Wujianshan of Fuyang923.653.75 452505594.31.831812.48.00临安方山Fangshan of Lin’an2946.1109.62 4446819523.71.839518.88.06
2.3.2 灰色关联度分析 对4个种群种实11个表型性状与土壤相关因子进行灰色关联度分析,结果见表6。结果显示:各表型性状与同一土壤因子的关联度系数差异较小,即不同表型性状与单一土壤因子无明显相关性。不同土壤因子与同一表型性状的关联度系数有明显差异,即同一个表型性状需要多个土壤因子的综合作用。与表型性状的关联度系数均值从大到小依次为pH值、Na含量、K含量、Cu含量、Mn含量、Ca含量、Mg含量、N含量、Fe含量、P含量、Zn含量,表明蜡梅种实的表型性状变异与土壤pH值的关联性最大,与Zn含量的关联性最小。
表6浙江省蜡梅野生种群种实表型性状与土壤相关因子的灰色关联度分析结果
Table6ResultofgreycorrelationdegreeanalysisonphenotypictraitsofseedandfruitandsoilrelatedfactorsofwildpopulationsofChimonanthuspraecox(Linn.)LinkinZhejiangProvince

土壤因子Soil factor与不同表型性状的关联度系数1) Correlation degree coefficient with different phenotypic traits1)SLSWCSFGWSFLFWCFFFGDWFPWFSNF均值AverageN含量 N content0.6970.7070.6800.7080.6940.6920.6980.7170.7140.6600.7370.700P含量 P content0.6790.6830.6670.6800.6730.6710.6850.6930.6690.6440.6870.676K含量 K content0.8190.8090.8120.8240.8400.8220.8250.8200.7890.7910.7940.813Ca含量 Ca content0.7630.7540.7440.7420.7660.7670.7390.7620.8500.7540.8470.772Mg含量 Mg content0.7480.7190.7510.7200.7570.7550.7300.7200.7020.7610.7090.734Fe含量 Fe content0.6650.6390.6630.6430.6890.6750.6530.6440.7260.7060.7640.679Zn含量 Zn content0.5440.5380.5420.5370.5380.5410.5360.5400.5340.5410.5430.539Cu含量 Cu content0.7660.7380.7640.7470.7960.7770.7580.7460.8340.7810.8800.781Mn含量 Mn content0.7710.7500.7590.7570.8060.7840.7640.7640.8060.7980.8050.779Na含量 Na content0.8750.8850.8590.8860.8490.8610.8740.8830.8110.8310.8150.857pH值 pH value0.9630.9590.9600.9510.9080.9440.9370.9400.8820.9200.8660.930
1)SL:种子长Seed length;SW:种子宽Seed width;CSF:种形系数Coefficient of seed form;GWS:种子千粒质量1 000-grain weight of seed;FL:果实长Fruit length;FW:果实宽Fruit width;CFF:果形系数Coefficient of fruit form;FGD:果口直径 Fruit gap diameter;WF:单果质量Weight per fruit;PWF:单果果壳质量 Pericarp weight per fruit;SNF:单果种子数Seed number per fruit.
2.4 基于种实表型性状的供试种群的聚类分析
基于种实11个表型性状,采用UPGMA法对4个蜡梅野生种群进行聚类分析。结果(图1)显示:在欧氏距离为1处,4个种群分为3组。其中,临安方山和富阳龙王坎2个种群归为一组,距离最小且表型性状特征相似;富阳五尖山和富阳碧东山种群分别单独归为一组。将欧氏距离与地理距离进行相关性分析发现,二者间不存在显著相关性,说明供试的4个蜡梅野生种群种实表型性状变异没有随地理距离呈连续变化的趋势。
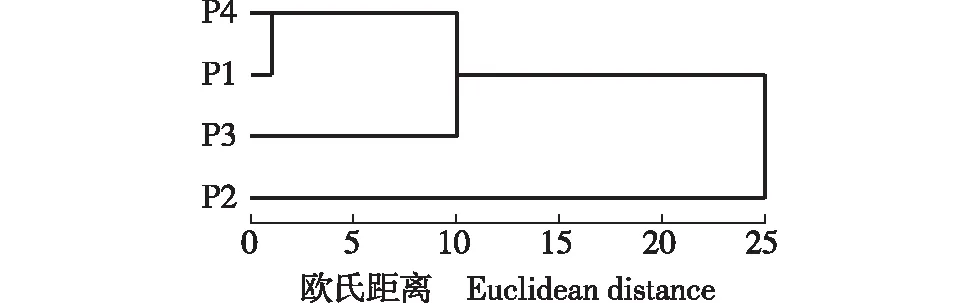
P1:富阳龙王坎Longwangkan of Fuyang;P2:富阳碧东山Bidongshan of Fuyang;P3:富阳五尖山Wujianshan of Fuyang;P4:临安方山Fangshan of Lin’an.图1 基于种实表型性状的浙江省4个蜡梅野生种群的聚类图Fig. 1 Cluster diagram of four wild populations of Chimonanthus praecox (Linn.) Link in Zhejiang Province based on phenotypic traits of seed and fruit
3 讨论和结论
表型性状既有变异性又有稳定性,是植物基因型和所处生境的综合体现。植物种群保持较高变异性有利于种群发展,变异系数越大,物种适应的环境条件越广。本研究结果表明:供试4个蜡梅野生种群的种子长、种子宽、种子千粒质量、果实长和果实宽的均值分别为15.48 mm、6.94 mm、325.24 g、49.82 mm和15.76 mm,大于其他蜡梅野生种群相应表型性状的最大均值(13.4 mm、5.88 mm、229.8 g、35.7 mm和15.7 mm)[21-22];另外,不同表型性状的变异系数差异较大(6.97%~45.72%),均值为19.98%,高于夏蜡梅种群表型性状的变异系数均值(12.15%)[23],表明浙江省蜡梅野生种群种实表型性状间存在比较丰富的遗传变异,因而,供试的浙江省4个蜡梅野生种群可作为优良的种质资源。由本研究结果还可见:蜡梅果实表型性状的变异系数明显大于种子表型性状,由此可知蜡梅的果实长、果实宽和果实质量等果实表型性状主导蜡梅种实表型性状的变异方向。这一现象可能与种子位于果实内部,受外界环境选择的压力小,种子表型性状较为稳定有关,在思茅松[8]和夏蜡梅[23]等种类中也存在类似的现象。供试4个蜡梅野生种群间表型分化系数均值为14.83%,低于张彩霞等[21]对蜡梅群体8个表型性状研究获得的表型分化系数(63.16%),这与研究中选取的表型性状不同有关。作者在实地调查中发现,受开花物候和海拔等生境因子的影响,野生的蜡梅个体传粉昆虫较少,种群可能存在较高的自交率,这也是其表型分化系数较低、种群内遗传变异占主导地位的原因之一。
Khurana等[7]认为,与生活在空旷、易受干扰生境的物种相比,生活在郁闭度高、稳定生境中的物种的种子更大。在供试的4个蜡梅野生种群中,富阳龙王坎和富阳碧东山2个种群种实的多数表型性状显著或极显著高于另2个种群,说明这2个种群种实表型性状更佳,这可能与不同种群的生境差异有关。富阳龙王坎和富阳碧东山2个种群的生境相似,海拔均约100 m,多沿陡峭的溪谷呈狭带状分布,且主要分布在沟谷中下部、溪边及山脊两侧,受人为影响较少;而富阳五尖山种群所处的群落无乔木层,其他灌木和草本植物也较少,群落内光照充足且相对空旷;临安方山种群所处的群落类型为落叶阔叶林,海拔较高、环境潮湿,且受周围村落的人为干扰较大。邓洪平等[24]认为,生长在土层厚、土壤肥沃等土壤条件较好的植物,其种子的长度、宽度、厚度、总质量等形态指标均明显大于生长于不良土壤条件的植物,且土壤类型对种子形态也有一定的影响。富阳碧东山土层厚且松软肥沃,而富阳五尖山和临安方山的土层薄、土壤贫瘠且石灰岩较多,因而,蜡梅野生种群种实表型性状差异可能是对不同生境适应的表现。
灰色关联度分析结果表明:土壤pH值与蜡梅野生种群种实表型性状的关联度系数较高。土壤pH值是土壤化学性质的综合反映,土壤中N、P和K元素的转化和释放,Ca、Fe和Mg等微量元素的有效性,有机质的合成和分解及微生物的活动等[25]都与土壤pH值相关,因而,与单一土壤因子的影响效应相比,其综合性更强,这可能是导致其关联度系数较高的原因。N、P、K和Ca等元素是植物体内重要物质(如核酸、蛋白质和叶绿素等)的组成成分,能直接或间接影响植物的生长发育和代谢;单一土壤因子不能单独对植物的某些表型性状起主要作用,植物表型性状的形成需要多个土壤因子甚至所有环境因子的综合作用。依据种实表型性状对供试蜡梅野生种群进行聚类分析,其结果与种群间的地理距离无关,说明在相对较小的分布范围内,遗传是蜡梅野生种群种实表型性状变异的主导因子。
综上所述,供试蜡梅野生种群种实表型性状具有丰富的变异,这与环境因子密切相关,但本研究只对部分土壤因子与种实表型性状进行了灰色关联度分析,而植物种实表型性状变异还可能与植物体大小、物候期、系统发育、基因控制和控制程度等因子有关,这些有待深入研究。
参考文献:
[1] 李 伟,林富荣,郑勇奇,等. 皂荚南方天然种群种实表型多样性[J]. 植物生态学报,2013,37(1):61-69.
[2] 孙玉玲,李庆梅,杨敬元,等. 秦岭冷杉球果与种子的形态变异[J]. 生态学报,2005,25(1):176-181.
[3] GARCIA R,SIEPIELSKI A M,BENKMAN C W. Cone and seed trait variation in whitebark pine (Pinusalbicaulis;Pinaceae) and the potential for phenotypic selection[J]. American Journal of Botany,2009,96(5):1050-1054.
[4] 柯文山,钟章成,席红安,等. 四川大头茶地理种群种子大小变异对萌发、幼苗特征的影响[J]. 生态学报,2000,20(4):697-701.
[5] ERIKKSON O. Seed size variation and its effects on germination and seedling performance in the clonal herbConvallariamajalis[J]. Acta Oecologica,1999,20(1):61-66.
[6] KHURANA E,SINGH J S. Influence of seed size on seedling growth ofAlbiziaproceraunder different soil water levels[J]. Annals of Botany,2000,86(6):1185-1192.
[7] KHURANA E,SAGAR R,SINGH J S. Seed size:a key trait determining species distribution and diversity of dry tropical forest in northern India[J]. Acta Oecologica,2006,29(2):196-204.
[8] 李帅锋,苏建荣,刘万德,等. 思茅松天然群体种实表型变异[J]. 植物生态学报,2013,37(11):998-1009.
[9] 张文标,金则新. 夏蜡梅果实和种子形态变异及其与环境因子相关性[J]. 浙江大学学报(理学版),2007,34(6):689-695.
[10] 蔡琰琳,金则新. 濒危植物夏蜡梅果实、种子形态变异研究[J]. 西北林学院学报,2008,23(3):44-49.
[11] 邵文豪,刁松峰,董汝湘,等. 无患子种实形态及经济性状的地理变异[J]. 林业科学研究,2013,26(5):603-608.
[12] 李全健,王彩霞,田 敏,等. 浙江扇脉杓兰野生居群的表型性状变异及其与地理-土壤养分因子的相关性[J]. 植物资源与环境学报,2012,21(2):45-52.
[13] 赵 冰,张启翔. 蜡梅种质资源表型多样性[J]. 东北林业大学学报,2007,35(5):10-13,35.
[14] 明 军,明刘斌. 蜡梅科植物种质资源研究进展[J]. 北京林业大学学报,2004,26(增刊):128-135.
[15] 明 军,廖卉荣. 蜡梅科植物种植资源研究文献分析[J]. 北京林业大学学报(社会科学版),2004,3(3):60-66.
[16] 李根有,金水虎,楼炉焕,等. 浙江省野生蜡梅数量及群落学研究[J]. 北京林业大学学报,2003,25(6):30-33.
[17] 李根有,楼炉焕,金水虎,等. 浙江省野生蜡梅群落及其区系[J]. 浙江林学院学报,2002,19(2):127-132.
[18] 葛 颂,王明庥,陈岳武. 用同工酶研究马尾松群体的遗传结构[J]. 林业科学,1988,24(4):399-409.
[19] 刘玉成,杜道林,岳 泉. 缙云山森林次生演替中优势种群的特性与生态因子的关联度分析[J]. 植物生态学报,1994,18(3):283-289.
[20] 肖宜安,何 平,邓洪平. 井冈山长柄双花木种群形态分化的数量分析[J]. 武汉植物学研究,2002,20(5):365-370.
[21] 张彩霞,左丹丹,赵海涛,等. 蜡梅天然群体表型多样性分析[J]. 西南林业大学学报,2011,31(5):19-23.
[22] 赵 冰,张启翔. 蜡梅种质资源表型多样性[J]. 东北林业大学学报,2007,35(5):10-13,35.
[23] 陈香波,叶文国,田 旗,等. 夏蜡梅天然群体表型变异及分布特征[J]. 北京林业大学学报,2010,32(2):133-140.
[24] 邓洪平,何 平,钟章成. 不同地理种源及演替群落的四川大头茶居群种子形态分化研究[J]. 西南师范大学学报(自然科学版),1999,24(2):207-213.
[25] 孙 凡,钟章成. 四川大头茶繁殖分配及其环境适应性的关联度研究[J]. 植物生态学报,1997,21(1):44-52.
猜你喜欢
种子世界(2024年10期)2024-12-31 00:00:00
作文小学中年级(2024年2期)2024-03-02 02:23:12
广西植物(2023年7期)2023-08-20 06:03:03
故事作文·低年级(2023年6期)2023-07-28 16:50:16
新农业(2022年7期)2022-05-03 10:05:58
科学大众(2021年6期)2021-07-20 07:42:50
凯里学院学报(2021年3期)2021-07-16 06:29:42
军工文化(2021年2期)2021-03-30 08:35:34
中国自行车(2018年11期)2018-12-03 08:20:34
中国自行车(2017年11期)2017-04-04 02:51:36
