
红豆树优树种子和幼苗性状的变异分析及优良家系的初选
2018-06-06 02:49:32李峰卿陈焕伟周志春楚秀丽徐肇友肖纪军
植物资源与环境学报 2018年2期
李峰卿,陈焕伟,周志春,楚秀丽,徐肇友,肖纪军
(1. 中国林业科学研究院亚热带林业研究所 浙江省林木育种技术研究重点实验室,浙江 杭州 311400;2. 中国林业科学研究院亚热带林业实验中心,江西 分宜 336600;3. 浙江省龙泉市林业科学研究院,浙江 龙泉 323700)
红豆树(OrmosiahosieiHemsl. et E. H. Wilson)隶属于豆科(Fabaceae)红豆属(OrmosiaJasks.),为半常绿或落叶乔木,又称花梨木,其木材质地坚硬,是上等的工艺、雕刻、装饰及镶嵌用材。红豆树自然分布于陕西南部、江苏、浙江、江西、福建、湖北、四川、重庆和贵州等地,是红豆属中分布最北、最耐寒的珍贵树种。长期以来,由于红豆树材质优异,天然林资源遭到过度利用和破坏,导致其资源匮乏。已有研究结果表明:较小的红豆树天然居群仍具有较高的遗传多样性[1-2]。由此可见,红豆树具有丰富的遗传变异,这为其遗传改良和高效培育提供了丰富的材料。近年来,在浙江、江西和福建等省,红豆树作为珍贵用材树种越来越被重视,红豆树育苗和造林关键技术取得突破[3-4],并开展了天然林和人工林的优树选择[5-6],亟需进一步开展家系水平的相关研究。
种子形态虽然具有较高的稳定性,但也受到与环境(气候和纬度等)相关的自然选择的影响[7-8],其变异是物种适应环境变化的综合结果[9-10]。植物生长节律,尤其是苗高生长节律关系到林木的存活及种间和种内竞争[11],且在种源、家系及个体等水平上均存在明显差异[12-15]。这些特点在林木遗传育种中具有重要价值[16-17],因此,开展种子形态、幼苗生长性状及苗高生长节律的研究,不仅可以揭示林木主要性状的遗传变异规律,还可以为科学制定林木育种方案并进行遗传改良提供理论依据。
鉴于此,作者在所在课题组前期研究[6]的基础上,从江西、浙江、福建和四川等红豆树主要分布区的天然群体中选择76株优树,对其种子和幼苗性状进行分析,调查了30个代表性家系幼苗的苗高生长节律,并初选出一批优良家系,以期揭示幼苗性状的家系遗传变异,明确不同家系幼苗的苗高生长节律,阐释种子变异对幼苗生长的影响,并为进一步的育种研究提供科学依据。
1 材料和方法
1.1 材料
2015年,采用优势木对比法[18]对供试红豆树进行材积评定并结合形质指标选择优树,所选优树间距在50 m以上。在江西资溪,浙江的龙泉和庆元,福建的泰宁、南平和柘荣及四川内江分别选择红豆树优树5、31、2、2、10、23和3株,分别编号为1~5、6~36、37~38、39~40、41~50、51~73和74~76,共76株,所选优树树龄均大于30 a,树高10~20 m,胸径19.4~95.3 cm。于当年10月至11月,密切观察各优树种子成熟状况,于种子成熟期采用竹竿人工敲打树枝,种子落地后集中收取,分别采集各优树单株自由授粉种子。除1和4号优树种子量较少外,其他优树分别收集育苗用种子0.2 kg。
1.2 方法
1.2.1 种子性状测量 各优树随机选取30粒种子,用电子游标卡尺(精度0.01 mm)分别测量种长(种子平行于种胚方向的长度)、种宽(种子腹面横向的宽度)和种厚(种子腹面与背面间的距离),均重复测量3次。然后计算种子长宽比。种子百粒质量测定采用四分法取样,用百分之一电子天平称量100粒种子的质量(不足100粒的换算为百粒质量)。
1.2.2 幼苗培育及其性状的测量和统计 2016年5月中旬,将采集的优树种子用50 ℃~60 ℃温水浸泡15~24 h并人工破皮后,及时播种于浙江省龙泉市林业科学研究院省级保障性苗圃,地理坐标为北纬28°02′、东经119°05′,海拔245 m,年均温17.6 ℃,年均降水量1 699.4 mm。苗圃地为黄壤土,其表层覆盖混合均匀的黄壤土和泥炭(体积比7∶3),排灌条件良好,土壤肥力中等。采用完全随机区组设计,以点播方式育苗,行距30 cm,株距10 cm,每小区播种3行,每行10株,每小区30株,3次重复。
每个采样点选择1~3个家系,共选择30个代表性家系进行苗高生长节律及叶片性状和根系性状的测量和统计。每小区选取中间一行的10株幼苗进行调查,并挂牌标记。从2016年6月13日(出苗后15 d)开始,用钢卷尺(精度0.1 cm)测量苗高(土壤表面到植株顶端的距离),之后每隔30 d定株测量苗高,直至12月20日。最后一次同时测量所有家系的苗高和地径(植株茎干在土壤表面处的直径)。其中,地径采用电子游标卡尺测量。幼苗生长性状测量完毕后,每小区挖取10株生长正常、平均大小的幼苗,分别测量和统计单株复叶数、单株叶片数、最大叶片的长度和宽度、主根长、最长侧根长以及单株长度大于5 cm的一级侧根数。
1.3 数据处理与分析
1.3.1 方差分析、相关性分析及家系遗传力估算 对每个性状的观测值进行统计,单株复叶数、单株叶片数以及单株长度大于5 cm的一级侧根数经X1/2数据转换,种子长宽比经arcsinX-1/2数据转换,然后采用SPSS 20.0统计分析软件进行方差分析(单样本t检验)和相关性分析。种子性状的方差分析以重复内单粒种子为单元,幼苗性状的方差分析以小区内单株为单元,苗高生长节律参数的方差分析以小区平均值为单元。
家系遗传力(hf2)按照公式“hf2=(v1-v2)/v1”进行估算。式中,v1为家系的方差,v2为家系与小区互作的方差。
1.3.2 幼苗生长模型及苗高生长节律参数计算 根据幼苗生长符合“S”型曲线特点,采用SPSS 20.0统计分析软件进行Logistic模型拟合。拟合方程为y=k/(1+ea-bt)。式中,y为家系幼苗苗高累计生长量,k为拟合苗高的极限值,t为生长时间,a和b为待定系数。参照杨志玲等[19]的方法计算红豆树各家系幼苗苗高生长节律参数,其中,物候期参数包括线性生长始期、线性生长末期和线性生长期,生长参数包括最大线性生长速率、线性生长速率和线性生长总量。
2 结果和分析
2.1 种子和幼苗性状的变异分析
2.1.1 种子性状变异分析 红豆树优树种子性状的方差分析结果见表1。由表1可以看出:红豆树优树间种子性状的差异均达到显著 (P<0.05)或极显著(P<0.01)水平。不同红豆树优树种子性状中变幅最大的为百粒质量,变幅为55.28~158.04 g,最大值为最小值的2.86倍,其表型变异系数达18.73%;种长、种宽、种厚和种子长宽比的变幅分别为11.49~18.03 mm、10.38~15.41 mm、6.06~9.80 mm和1.01~1.48,最大值分别为最小值的1.57、1.48、1.62和1.47倍,4个性状的表型变异系数接近,分别为9.04%、7.79%、9.18%和8.07%。
表1红豆树优树种子性状的方差分析
Table1VarianceanalysisonseedtraitsofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson
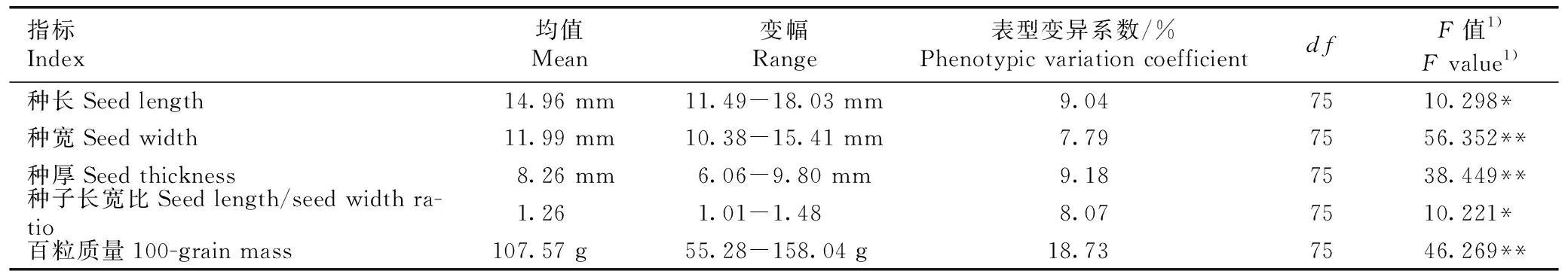
指标Index均值Mean变幅Range表型变异系数/%Phenotypic variation coefficientdfF值1)F value1)种长Seed length14.96 mm11.49-18.03 mm9.047510.298*种宽Seed width11.99 mm10.38-15.41 mm7.797556.352**种厚Seed thickness8.26 mm6.06-9.80 mm9.187538.449**种子长宽比Seed length/seed width ra-tio1.261.01-1.488.077510.221*百粒质量100-grain mass107.57 g55.28-158.04 g18.737546.269**
1)*:P<0.05;** :P<0.01.
2.1.2 幼苗生长性状变异分析 红豆树优树幼苗苗高和地径的方差分析及家系遗传力估算结果见表2。由表2可以看出:红豆树优树幼苗的苗高和地径在家系间的差异均达到极显著水平。不同家系红豆树优树幼苗苗高和地径的变幅分别为23.57~49.77 cm和5.54~8.32 mm,苗高和地径的最大值分别为最小值的2.11和1.50倍,且苗高的遗传变异系数(35.52%)明显大于地径(16.32%)。苗高和地径的家系遗传力均较高,分别为0.854和0.719。
表2红豆树优树幼苗生长性状的方差分析及家系遗传力估算
Table2VarianceanalysisandfamilyheritabilityestimationofseedlinggrowthtraitsofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson
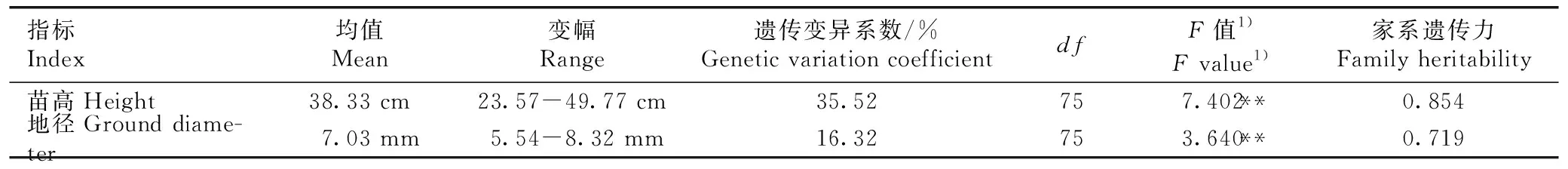
指标Index均值Mean变幅Range遗传变异系数/%Genetic variation coefficientdfF值1)F value1)家系遗传力Family heritability苗高Height38.33 cm23.57-49.77 cm35.52757.402**0.854地径Ground diame-ter7.03 mm5.54-8.32 mm16.32753.640**0.719
1)** :P<0.01.
2.1.3 幼苗苗高生长节律参数分析 对红豆树优树30个代表性家系幼苗的苗高生长节律数据进行Logistic模型拟合,各家系Logistic拟合方程的决定系数为0.967~1.000,均达到极显著水平,表明用Logistic模型拟合的家系幼苗的苗高与实测值的符合程度较高。
红豆树优树30个代表性家系幼苗苗高生长节律参数见表3。由表3可以看出:红豆树优树幼苗苗高生长节律参数在家系间的差异均达到极显著水平。红豆树优树30个代表性家系间线性生长始期差异较大,最多相差21.5 d,其中,浙江龙泉3(8号)家系线性生长始期最早(6月9日),福建柘荣21(71号)家系线性生长始期最晚(7月1日)。家系间线性生长末期也存在较大差异,线性生长末期最早和最晚的家系分别为福建南平9(49号)和福建南平2(42号)家系,二者间相差38.1 d。福建柘荣1(51号)家系线性生长期最短(112.8 d),而福建南平6(46号)家系线性生长期最长(150.4 d),二者间相差37.6 d。多数家系在6月中旬进入线性生长期,历时130 d左右,于10月中下旬线性生长期结束。浙江庆元2(38号)家系的最大线性生长速率、线性生长速率和线性生长总量均最高,而浙江龙泉20(25号)家系这3个指标均最低,且前者这3个指标分别为后者的2.00、1.91和1.90倍。
表3红豆树优树30个代表性家系幼苗苗高生长节律参数1)
Table3Parametersofheightgrowthrhythmofseedlingsof30representativefamiliesofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson1)
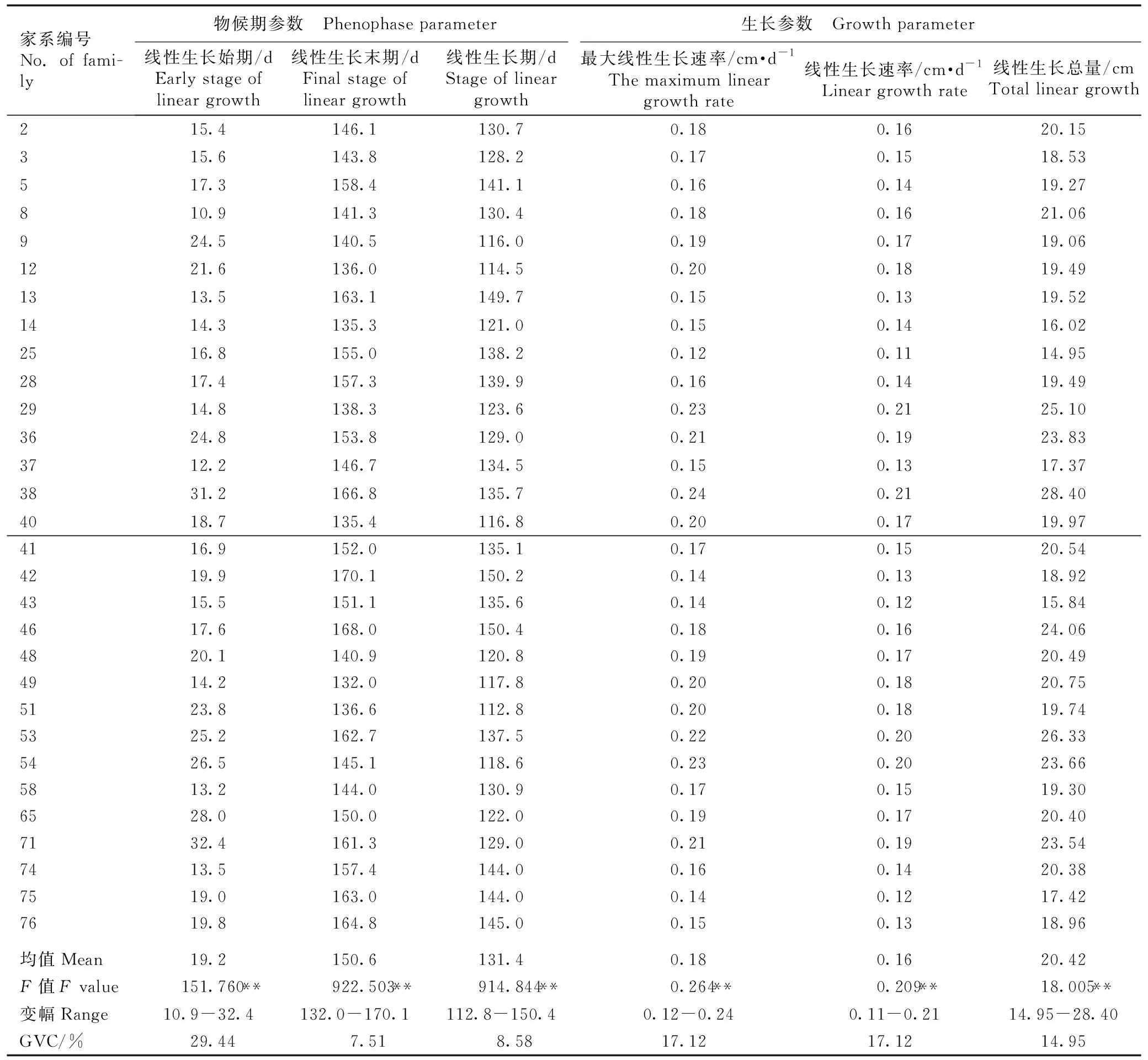
家系编号No. of fami-ly物候期参数 Phenophase parameter生长参数 Growth parameter线性生长始期/dEarly stage of linear growth线性生长末期/dFinal stage of linear growth线性生长期/dStage of linear growth最大线性生长速率/cm·d-1The maximum linear growth rate线性生长速率/cm·d-1Linear growth rate线性生长总量/cmTotal linear growth215.4146.1130.70.180.1620.15315.6143.8128.20.170.1518.53517.3158.4141.10.160.1419.27810.9141.3130.40.180.1621.06924.5140.5116.00.190.1719.061221.6136.0114.50.200.1819.491313.5163.1149.70.150.1319.521414.3135.3121.00.150.1416.022516.8155.0138.20.120.1114.952817.4157.3139.90.160.1419.492914.8138.3123.60.230.2125.103624.8153.8129.00.210.1923.833712.2146.7134.50.150.1317.373831.2166.8135.70.240.2128.404018.7135.4116.80.200.1719.974116.9152.0135.10.170.1520.544219.9170.1150.20.140.1318.924315.5151.1135.60.140.1215.844617.6168.0150.40.180.1624.064820.1140.9120.80.190.1720.494914.2132.0117.80.200.1820.755123.8136.6112.80.200.1819.745325.2162.7137.50.220.2026.335426.5145.1118.60.230.2023.665813.2144.0130.90.170.1519.306528.0150.0122.00.190.1720.407132.4161.3129.00.210.1923.547413.5157.4144.00.160.1420.387519.0163.0144.00.140.1217.427619.8164.8145.00.150.1318.96均值Mean19.2150.6131.40.180.1620.42F值F value151.760**922.503**914.844**0.264**0.209**18.005**变幅Range10.9-32.4132.0-170.1112.8-150.40.12-0.240.11-0.2114.95-28.40GVC/%29.447.518.5817.1217.1214.95
1)2:江西资溪2 Zixi 2 of Jiangxi;3:江西资溪6 Zixi 6 of Jiangxi;5:江西资溪7 Zixi 7 of Jiangxi;8:浙江龙泉3 Longquan 3 of Zhejiang;9:浙江龙泉4 Longquan 4 of Zhejiang;12:浙江龙泉7 Longquan 7 of Zhejiang;13:浙江龙泉8 Longquan 8 of Zhejiang;14:浙江龙泉9 Longquan 9 of Zhejiang;25:浙江龙泉20 Longquan 20 of Zhejiang;28:浙江龙泉23 Longquan 23 of Zhejiang;29:浙江龙泉24 Longquan 24 of Zhejiang;36:浙江龙泉31 Longquan 31 of Zhejiang;37:浙江庆元1 Qingyuan 1 of Zhejiang;38:浙江庆元2 Qingyuan 2 of Zhejiang;40:福建泰宁2 Taining 2 of Fujian;41:福建南平1 Nanping 1 of Fujian;42:福建南平2 Nanping 2 of Fujian;43:福建南平3 Nanping 3 of Fujian;46:福建南平6 Nanping 6 of Fujian;48:福建南平8 Nanping 8 of Fujian;49:福建南平9 Nanping 9 of Fujian;51:福建柘荣1 Zherong 1 of Fujian;53:福建柘荣3 Zherong 3 of Fujian;54:福建柘荣4 Zherong 4 of Fujian;58:福建柘荣8 Zherong 8 of Fujian;65:福建柘荣15 Zherong 15 of Fujian;71:福建柘荣21 Zherong 21 of Fujian;74:四川内江1 Neijiang 1 of Sichuan;75:四川内江2 Neijiang 2 of Sichuan;76:四川内江3 Neijiang 3 of Sichuan. ** :P<0.01. GVC:遗传变异系数Genetic variation coefficient.
从红豆树优树30个代表性家系幼苗苗高生长节律参数的遗传变异系数可以看出:遗传变异系数最大的是线性生长始期,为29.44%;最大线性生长速率(17.12%)、线性生长速率(17.12%)和线性生长总量(14.95%)的遗传变异系数居中;线性生长末期(7.51%)和线性生长期(8.58%)的遗传变异系数较小。
2.1.4 幼苗叶片性状和根系性状变异分析 红豆树优树幼苗叶片性状和根系性状的方差分析及家系遗传力估算见表4。由表4可以看出:红豆树优树幼苗的叶片性状和根系性状在家系间的差异均达到显著或极显著水平。红豆树各家系幼苗单株复叶数、单株叶片数、最大叶片的长度和最大叶片的宽度的最大值分别为最小值的1.93、2.06、1.46和1.50倍。不同家系红豆树幼苗的主根长、最长侧根长和单株长度大于5 cm的一级侧根数的变幅分别为21.73~35.97 cm、10.90~20.61 cm和6.4~16.6,3个性状的最大值分别为最小值的1.66、1.89和2.59倍。
红豆树幼苗叶片性状和根系性状遗传变异系数的变化范围为21.23%~42.58%,其中,最长侧根长的遗传变异系数最大(42.58%),其次为最大叶片的长度(31.77%),单株复叶数的遗传变异系数最小(21.23%)。红豆树幼苗叶片性状和根系性状家系遗传力的变化范围为0.479~0.851,其中,最大叶片的长度、最大叶片的宽度、主根长和最长侧根长的家系遗传力较大,分别为0.851、0.705、0.764和0.704。
表4红豆树优树幼苗叶片性状和根系性状的方差分析及家系遗传力估算
Table4VarianceanalysisandfamilyheritabilityestimationofleaftraitsandroottraitsofseedlingsofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson
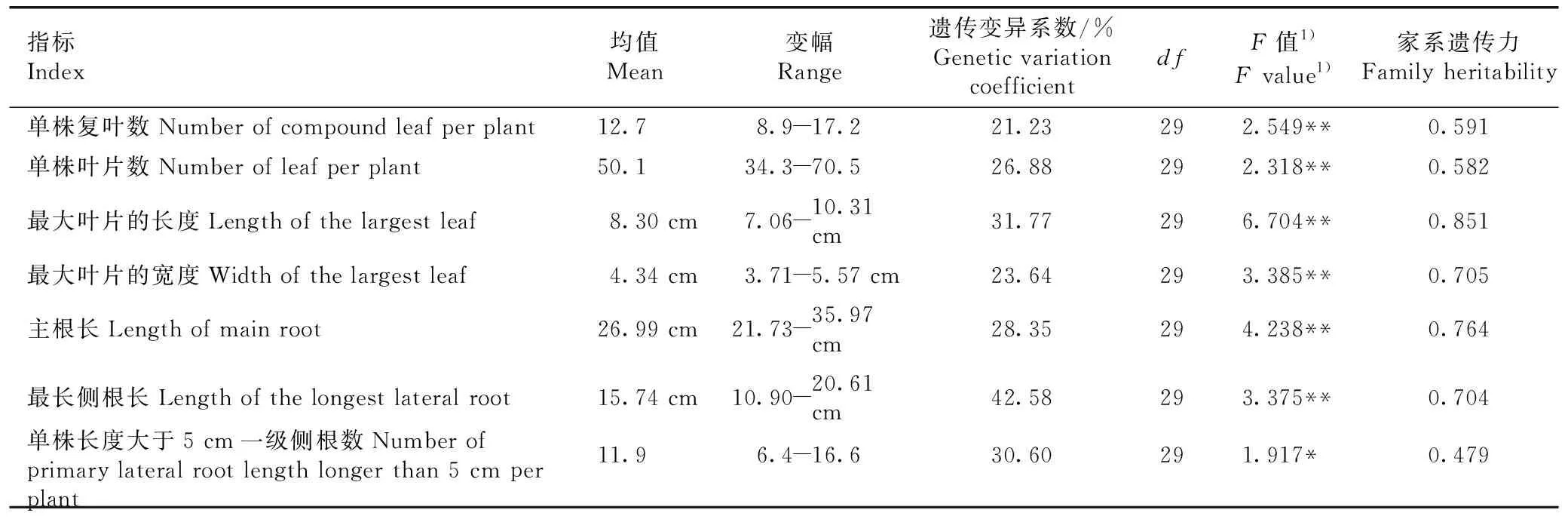
指标Index均值Mean变幅Range遗传变异系数/%Genetic variation coefficientdfF值1)F value1)家系遗传力Family heritability单株复叶数Number of compound leaf per plant12.7 8.9-17.221.23292.549**0.591单株叶片数Number of leaf per plant50.134.3-70.526.88292.318**0.582最大叶片的长度Length of the largest leaf8.30 cm7.06-10.31 cm31.77296.704**0.851最大叶片的宽度Width of the largest leaf4.34 cm3.71-5.57 cm23.64293.385**0.705主根长Length of main root26.99 cm21.73-35.97 cm28.35294.238**0.764最长侧根长Length of the longest lateral root15.74 cm10.90-20.61 cm42.58293.375**0.704单株长度大于5 cm一级侧根数Number of primary lateral root length longer than 5 cm per plant11.96.4-16.630.60291.917*0.479
1)*:P<0.05;** :P<0.01.
2.2 相关性分析
2.2.1 种子性状与幼苗生长性状间的相关性分析红豆树优树种子性状与幼苗生长性状的相关性分析结果见表5。由表5可以看出:红豆树优树的种厚和百粒质量与苗高和地径均呈极显著(P<0.01)正相关,相关系数为0.388~0.599;种宽与地径也呈极显著正相关,相关系数为0.317,但种宽与苗高呈不显著正相关;种长和种子长宽比与苗高和地径均呈不显著正相关。
2.2.2 幼苗生长性状、叶片性状及根系性状间的相关性分析 红豆树优树幼苗生长性状、叶片性状及根系性状间的相关性分析结果见表6。由表6可以看出:在表型相关系数中,红豆树优树幼苗地径与苗高以及叶片和根系性状呈显著(P<0.05)或极显著正相关,表型相关系数为0.343~0.746;苗高与单株长度大于5 cm的一级侧根数呈极显著正相关,表型相关系数为0.644;最大叶片的长度和最大叶片的宽度与主根长、最长侧根长和单株长度大于5 cm的一级侧根数均呈极显著正相关,表型相关系数为0.493~0.639。此外,单株复叶数与单株叶片数呈极显著正相关,表型相关系数为0.953;最大叶片的长度与最大叶片的宽度呈极显著正相关,表型相关系数为0.833;最长侧根长与单株长度大于5 cm的一级侧根数呈显著正相关,表型相关系数为0.406。
表5红豆树优树种子性状与幼苗生长性状的相关系数1)
Table5CorrelationcoefficientbetweenseedtraitsandseedlinggrowthtraitsofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson1)
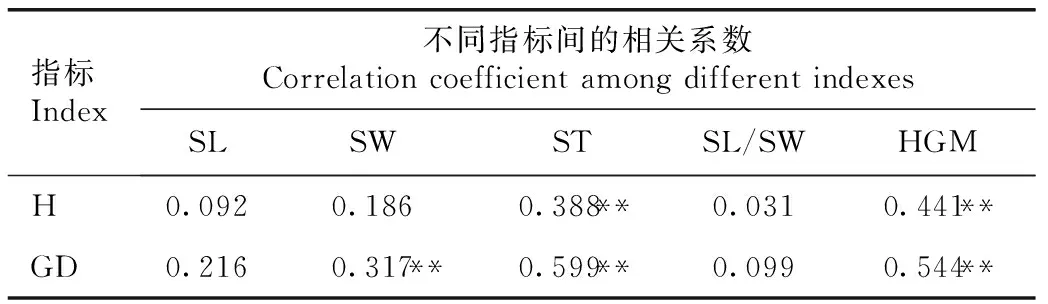
指标Index不同指标间的相关系数Correlation coefficient among different indexesSLSWSTSL/SWHGMH0.0920.1860.388**0.0310.441**GD0.2160.317**0.599**0.0990.544**
1)SL:种长Seed length;SW:种宽Seed width;ST:种厚Seed thickness;SL/SW:种子长宽比Seed length/seed width ratio;HGM:百粒质量100-grain mass;H:苗高Height;GD:地径Ground diameter. ** :P<0.01.
表6红豆树优树幼苗生长性状、叶片性状及根系性状间的相关系数1)
Table6Correlationcoefficientamonggrowthtraits,leaftraits,androottraitsofseedlingsofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson1)
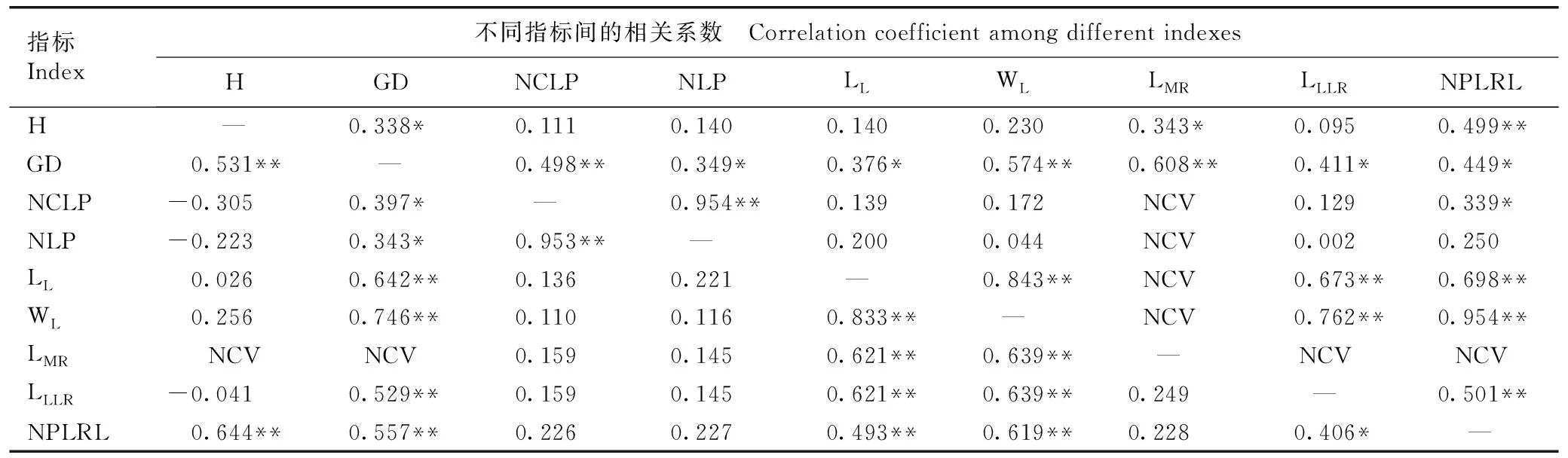
指标Index不同指标间的相关系数 Correlation coefficient among different indexesHGDNCLPNLPLLWLLMRLLLRNPLRLH—0.338*0.1110.1400.1400.2300.343*0.0950.499**GD0.531**—0.498**0.349*0.376*0.574**0.608**0.411*0.449*NCLP-0.3050.397*—0.954**0.1390.172NCV0.1290.339*NLP-0.2230.343*0.953**—0.2000.044NCV0.0020.250LL0.0260.642**0.1360.221—0.843**NCV0.673**0.698**WL0.2560.746**0.1100.1160.833**—NCV0.762**0.954**LMRNCVNCV0.1590.1450.621**0.639**—NCVNCVLLLR-0.0410.529**0.1590.1450.621**0.639**0.249—0.501**NPLRL0.644**0.557**0.2260.2270.493**0.619**0.2280.406*—
1)H:苗高Height;GD:地径Ground diameter;NCLP:单株复叶数Number of compound leaf per plant;NLP:单株叶片数Number of leaf per plant;LL:最大叶片的长度Length of the largest leaf;WL:最大叶片的宽度Width of the largest leaf;LMR:主根长Length of main root;LLLR:最长侧根长Length of the longest lateral root;NPLRL:单株长度大于5 cm一级侧根数Number of primary lateral root length longer than 5 cm per plant. “—”上方和下方的数据分别为遗传相关系数和表型相关系数Data above and below “—” are genetic correlation coefficient and phenotypic correlation coefficient,respectively. NCV:环境方差过大,遗传相关系数不可估计Environmental variance is too large,so genetic correlation coefficient cannot be estimated. *:P<0.05;** :P<0.01.
在遗传相关系数中,除主根长外,红豆树优树幼苗生长性状、叶片性状及根系性状间的相关性分析结果与表型相关性基本一致。
2.2.3 苗高及其生长节律参数间的相关性分析 红豆树优树幼苗苗高及其生长节律参数间的相关性分析结果见表7。由表7可以看出:红豆树优树幼苗苗高与最大线性生长速率、线性生长速率和线性生长总量均呈极显著正相关,表型相关系数为0.877~0.897,遗传相关系数为0.903~0.948,与线性生长期呈不显著负相关。苗高与线性生长始期呈正相关,但仅在遗传相关性分析中达到极显著水平。最大线性生长速率和线性生长速率与线性生长期均呈极显著负相关,表型相关系数分别为-0.528和-0.529,遗传相关系数分别为-0.677和-0.683;与线性生长总量均呈极显著正相关,表型相关系数分别为0.867和0.866,遗传相关系数分别为0.578和0.561;与线性生长始期也均呈极显著正相关,表型相关系数分别为0.578和0.579,遗传相关系数分别为0.469和0.496;与线性生长末期均呈负相关,但仅在遗传相关性分析中达到极显著水平,表型相关系数均为-0.239,遗传相关系数分别为-0.568和-0.555。线性型生长末期与线性生长期以及最大线性生长速率与线性生长速率均呈极显著正相关,表型相关系数分别为0.875和1.000,遗传相关系数分别为0.925和1.000。
表7红豆树优树幼苗苗高及其生长节律参数间的相关系数1)
Table7CorrelationcoefficientamongheightanditsgrowthrhythmparametersofseedlingsofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilson1)
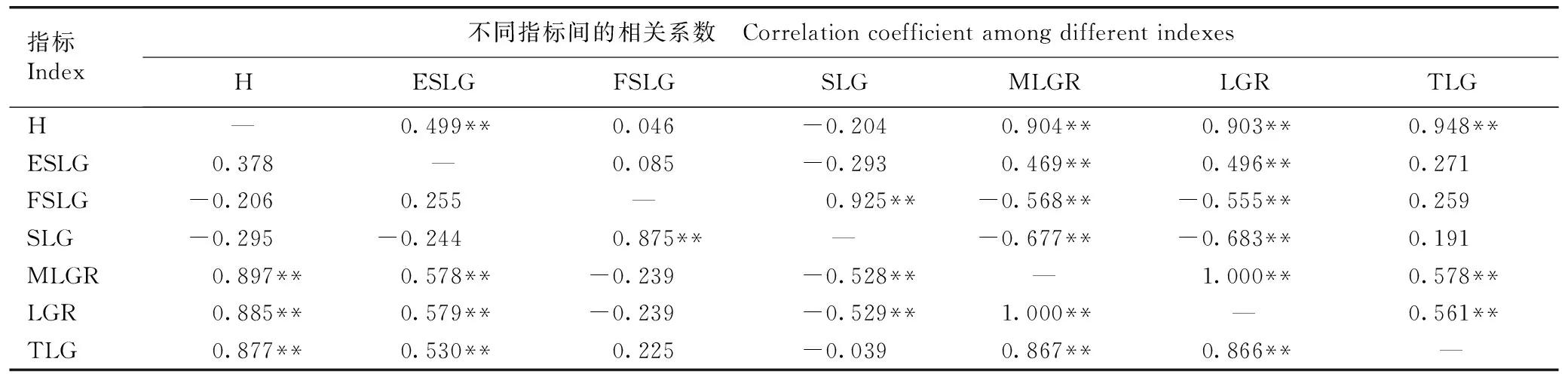
指标Index不同指标间的相关系数 Correlation coefficient among different indexesHESLGFSLGSLGMLGRLGRTLGH—0.499**0.046-0.2040.904**0.903**0.948**ESLG0.378—0.085-0.2930.469**0.496**0.271FSLG-0.2060.255—0.925**-0.568**-0.555**0.259SLG-0.295-0.2440.875**—-0.677**-0.683**0.191MLGR0.897**0.578**-0.239-0.528**—1.000**0.578**LGR0.885**0.579**-0.239-0.529**1.000**—0.561**TLG0.877**0.530**0.225-0.0390.867**0.866**—
1)H:苗高Height;ESLG:线性生长始期Early stage of linear growth;FSLG:线性生长末期Final stage of linear growth;SLG:线性生长期Stage of linear growth;MLGR:最大线性生长速率The maximum linear growth rate;LGR:线性生长速率Linear growth rate;TLG:线性生长总量Total linear growth. “—”上方和下方的数据分别为遗传相关系数和表型相关系数Data above and below “—” are genetic correlation coefficient and phenotypic correlation coefficient,respectively. ** :P<0.01.
2.3 红豆树优树优良家系初选
在不同层次上,对林木生长性状进行一定强度的选择,能获得较高的遗传增益。遗传增益与性状遗传力和选择强度等密切相关。本研究中,由于红豆树幼苗苗高的家系遗传力大于地径,但地径与叶片性状和根系性状均呈显著或极显著正相关,因此,选择红豆树优良家系时以苗高作为主要指标,同时兼顾地径。
以25%的入选率对红豆树优树76个家系进行初选,入选的优良家系及其幼苗生长性状见表8。由表8可以看出:入选的19个优良家系中,63.2%的优良家系来自浙江龙泉,26.3%的优良家系来自福建柘荣,来自福建南平和浙江庆元的优良家系各1个。19个优良家系幼苗苗高平均值为44.62 cm,较76个家系幼苗苗高平均值高16.41%;地径平均值7.37 mm,较76个家系幼苗地径平均值高4.84%。苗高和地径最大的优良家系均为浙江龙泉12(17号)家系,浙江龙泉11(16号)家系的苗高和浙江龙泉19(24号)家系的地径次之。
表8红豆树优树19个优良家系幼苗生长性状及其排序1)
Table8Growthtraitsofseedlingsof19optimumfamiliesofsuperiortreesofOrmosiahosieiHemsl.etE.H.Wilsonandtheirorder1)
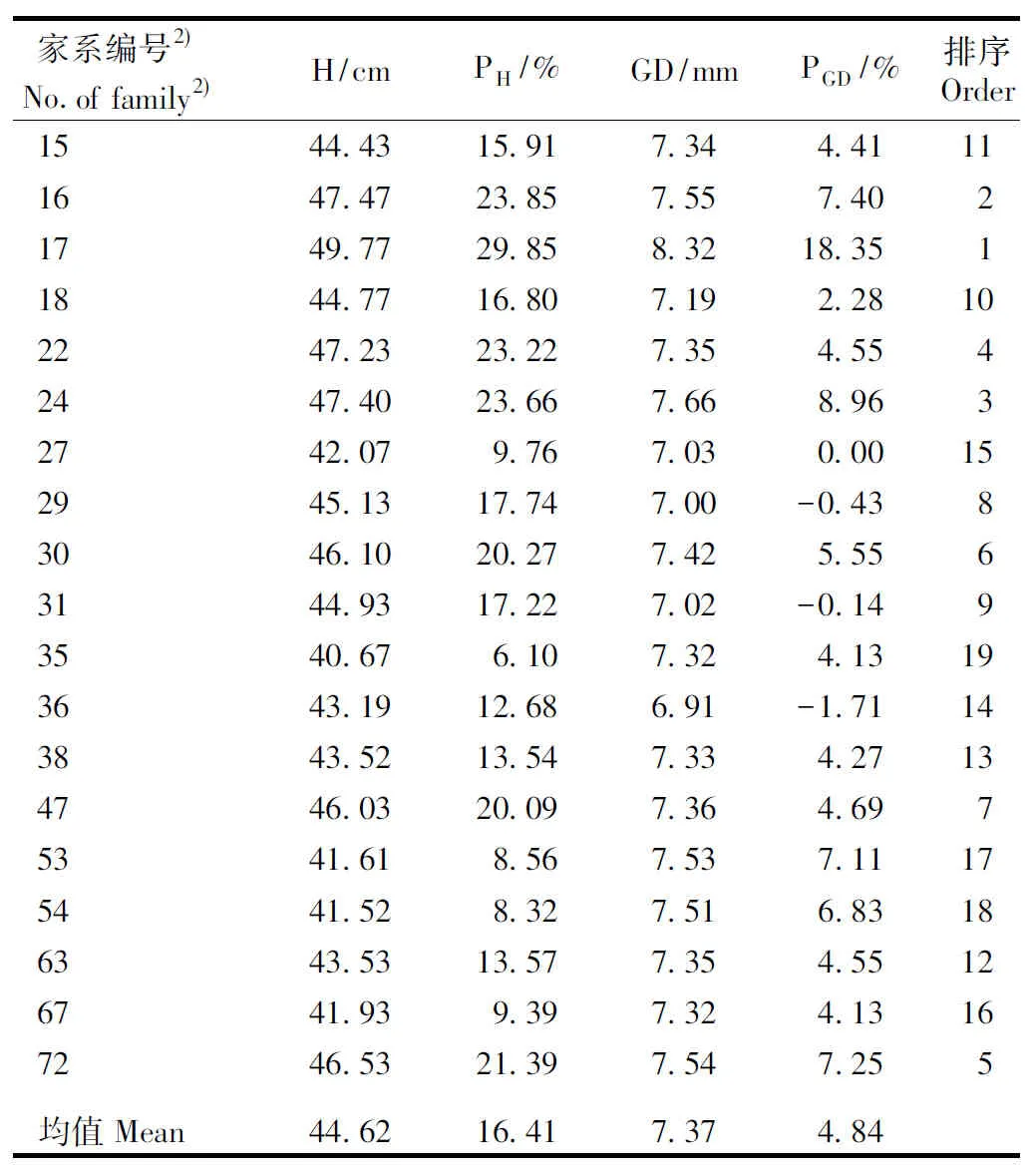
家系编号2)No. of fami-ly2)H/cmPH/%GD/mmPGD/%排序Order1544.4315.917.344.41111647.4723.857.557.4021749.7729.858.3218.3511844.7716.807.192.28102247.2323.227.354.5542447.4023.667.668.9632742.079.767.030.00152945.1317.747.00-0.4383046.1020.277.425.5563144.9317.227.02-0.1493540.676.107.324.13193643.1912.686.91-1.71143843.5213.547.334.27134746.0320.097.364.6975341.618.567.537.11175441.528.327.516.83186343.5313.577.354.55126741.939.397.324.13167246.5321.397.547.255均值Mean44.6216.417.374.84
1)H:苗高Height;PH:优良家系苗高较76个家系苗高平均值变化的百分率Percentage of change of height of optimum family to mean of height of 76 families;GD:地径Ground diameter;PGD:优良家系地径较76个家系地径平均值变化的百分率Percentage of change of ground diameter of optimum family to mean of ground diameter of 76 families.
2)15:浙江龙泉10 Longquan 10 of Zhejiang;16:浙江龙泉11 Longquan 11 of Zhejiang;17:浙江龙泉12 Longquan 12 of Zhejiang;18:浙江龙泉13 Longquan 13 of Zhejiang;22:浙江龙泉17 Longquan 17 of Zhejiang;24:浙江龙泉19 Longquan 19 of Zhejiang;27:浙江龙泉22 Longquan 22 of Zhejiang;29:浙江龙泉24 Longquan 24 of Zhejiang;30:浙江龙泉25 Longquan 25 of Zhejiang;31:浙江龙泉26 Longquan 26 of Zhejiang;35:浙江龙泉30 Longquan 30 of Zhejiang;36:浙江龙泉31 Longquan 31 of Zhejiang;38:浙江庆元2 Qingyuan 2 of Zhejiang;47:福建南平7 Nanping 7 of Fujian;53:福建柘荣3 Zherong 3 of Fujian;54:福建柘荣 4 Zherong 4 of Fujian;63:福建柘荣13 Zherong 13 of Fujian;67:福建柘荣17 Zherong 17 of Fujian;72:福建柘荣22 Zherong 22 of Fujian.
3 讨 论
林木在种内的不同层次和不同性状间均存在丰富的遗传变异[20],这为研究人工驯化和遗传育种提供了基础[21-22]。与木荷(SchimasuperbaGardn. et Champ.)[23]和薄壳山核桃〔Caryaillinoensis(Wangenh.) K. Koch〕[24]等树种的相关研究结果相似,红豆树优树间的种子性状及家系间幼苗的苗高、地径、叶片和根系等相关性状均具有显著(P<0.05)或极显著(P<0.01)差异,说明红豆树不同优树的种子和幼苗性状的分化均较大,有较大的选择潜力,这为其优良家系或单株的选择提供了理论依据和物质基础。生长节律参数在种源间和家系间均存在较大的遗传变异[13,25]。红豆树幼苗苗高生长节律参数在家系间也存在极显著差异。红豆树幼苗苗高和地径的家系遗传力分别为0.854和0.719,均受较强程度的家系遗传控制。木荷[23]和南方红豆杉〔Taxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li〕[13]苗期性状的种源(家系)遗传力较高,可根据苗期性状对优良种源(家系)进行选择。叶片性状受遗传高度控制。红豆树幼苗最大叶片的长度和宽度的家系遗传力分别为0.851和0.705,二者不仅在家系间差异显著,还均与地径呈显著或极显著正相关,因此,可通过红豆树幼苗的叶片大小间接选择优良家系,这一结果与林磊等[23]对木荷的研究结果一致,但还需要进一步开展幼苗的遗传测定和分析进行验证。综上所述,红豆树家系间存在较大的遗传改良潜力。由于本研究以1年生红豆树幼苗为研究对象,而苗期各家系性状尚不稳定,因此,本研究结果只能为红豆树优良家系的早期选育提供参考。此外,表型性状是基因与环境互作的结果,而本研究关于基因和环境对表型性状的贡献大小并未涉及,需在此基础上结合生理指标及DNA分子标记技术进行辅助选择,以获得更准确的优选效果。
种子形态特征对苗木生长影响结果不尽相同。本研究中,红豆树优树的种宽、种厚和百粒质量与幼苗的苗高和地径总体上存在显著正相关,说明根据种子性状可对红豆树优树幼苗生长进行早期选择,与曾祥艳等[26]的研究结果类似,但与多数研究结果不尽相同[20,27],具体原因有待进一步研究。研究红豆树家系苗木性状有助于通过易观测性状间接进行家系优选,并通过管理发挥苗木的生长潜力。根据红豆树各性状间相关性分析结果,较之苗高,地径与苗木生长各指标(叶片和根系)的关系更为密切。红豆树幼苗地径生长受植株光合面积和根系生长的双重制约,而苗高受根系生长的影响更为明显,但也有研究认为苗高亦受植株光合面积和根系生长的双重影响[28]。红豆树幼苗苗高与其生长节律参数的相关性分析结果表明:线性生长量较大的家系具有较高的最大线性生长速率和线性生长速率,而与线性生长期的关系不紧密,因此,在苗木培育过程中,可以采用加强水肥管理等方式提高线性生长速率,而非通过延长线性生长期达到促进苗高生长的目的。这一研究结果与张萍等[28]的研究结论类似,却有别于杨志玲等[19]的研究结果,推测与各树种不同的生物学特性有关。
本文以苗高作为主要筛选指标,同时兼顾地径,从红豆树优树76个家系中选出生长表现优异的19个优良家系,可为红豆树育种及无性繁殖提供材料。入选的红豆树家系中主要包括来自浙江龙泉和福建柘荣的家系,线性生长总量和线性生长速率较大的浙江龙泉24、浙江龙泉31、浙江庆元2、福建柘荣3和福建柘荣4家系均包括在内,说明浙江省和福建省可能是红豆树的中心产区和优良种源区。由于不同产区的气候和水热条件不同,长期生长在不同环境条件下的红豆树形成了不同的基因型,因此,还应加强浙江省和福建省以外的红豆树主要分布区的优树选择。鉴于苗木生长节律存在显著的家系与地点互作效应,还需开展多地点的红豆树优树幼苗测量以充分了解不同家系的生长习性和遗传特性,以期为不同地区选择出适生的优良家系,发挥其最大生长潜力。
参考文献:
[1] ZHANG R,ZHOU Z,DU K. Genetic diversity of natural populations of endangeredOrmosiahosiei,endemic to China[J]. Biochemical Systematics and Ecology,2012,40:13-18.
[2] 李峰卿,周志春,谢耀坚. 3个小流域红豆树天然居群的遗传多样性和遗传分化[J]. 分子植物育种,2017,15(10):4263-4274.
[3] 郑天汉. 红豆树苗木的质量性状分析及其主因子选择[J]. 福建林业科技,2008,35(1):63-66.
[4] 冯建国,季新良,周志春,等. 特种经济高档用材红豆树培育技术[J]. 林业科技开发,2007,21(5):93-95.
[5] 郑天汉,兰思仁. 红豆树天然林优树选择[J]. 福建农林大学学报(自然科学版),2013,42(4):366-370.
[6] 郑天汉,兰思仁. 红豆树人工林优树选择[J]. 福建农林大学学报(自然科学版),2013,42(6):610-615.
[8] 洑香香,刘红娜,周晓东,等. 山茱萸种子形态变异及与环境因子的相关性[J]. 生态学杂志,2013,32(1):27-32.
[9] PIGLIUCCI M,MURREN C J,SCHLICHTING C D. Phenotypic plasticity and evolution by genetic assimilation[J]. The Journal of Experimental Biology,2006,209:2362-2367.
[10] 林 玲,王军辉,罗 建,等. 砂生槐天然群体种实性状的表型多样性[J]. 林业科学,2014,50(4):137-143.
[11] LUKKARINEN A J,RUOTSALAINEN S,PELTOLA H,et al. Annual growth rhythm ofLarixsibiricaandLarixgmeliniiprovenances in a field trial in southern Finland[J]. Scandinavian Journal of Forest Research,2013,28:518-532.
[12] 张亚东,黄国伟,刘少峰,等. ‘美洲黑杨’不同家系苗期生长特征比较[J]. 中国农学通报,2015,31(10):1-6.
[13] 肖 遥,楚秀丽,徐肇友,等. 南方红豆杉2年生容器苗多点试验的生长节律家系变异[J]. 林业科学研究,2016,29(2):238-244.
[14] 贾庆彬,张含国,张 磊,等. 杂种落叶松优良家系选择与生长节律分析[J]. 北京林业大学学报,2016,38(2):52-60.
[16] 祝列克. 新世纪中国林木遗传育种发展战略[J]. 南京林业大学学报(自然科学版),2001,25(1):3-8.
[17] 王明庥. 森林遗传管理的现代基础理论与技术:林木遗传育种学[J]. 南京林业大学学报(自然科学版),2001,25(5):1-5.
[18] 南京林学院树木育种研究室. 树木良种选育方法[M]. 北京:中国林业出版社,1984:68-97.
[19] 杨志玲,杨 旭,谭梓峰,等. 厚朴不同种源苗期生长模型的拟合[J]. 西北农林科技大学学报(自然科学版),2011,39(4):60-68.
[20] 张 鹏,沈海龙. 花曲柳优树种子性状变异及其子代苗期生长表现[J]. 林业科技开发,2013,27(2):18-22.
[21] 曾 杰,郑海水,甘四明,等. 广西西南桦天然居群的表型变异[J]. 林业科学,2005,41(2):59-65.
[22] 姜顺邦,张 怡,韦小丽,等. 不同闽楠优树子代苗期生长及光合生理特性比较[J]. 浙江农林大学学报,2016,33(1):51-59.
[23] 林 磊,周志春,范辉华,等. 木荷优树子代苗期生长遗传和变异研究[J]. 林业科学研究,2009,22(2):155-160.
[24] 李永荣,李晓储,吴文龙,等. 66个薄壳山核桃实生单株果实性状变异选择研究[J]. 林业科学研究,2013,26(4):438-446.
[25] 肖 遥,楚秀丽,尹增芳,等. 不同产地南方红豆杉各家系幼苗生长、光合生理与株高生长节律的差异分析[J]. 植物资源与环境学报,2016,25(1):34-42.
[26] 曾祥艳,陈金艳,李开祥,等. 多穗柯优树种子性状变异及其子代苗期生长差异[J]. 广西林业科学,2015,44(4):358-362.
[27] 栗宏林,张志翔,张 鑫. 小桐子不同产地种子性状及苗期生长差异研究[J]. 干旱区资源与环境,2010,24(2):204-208.
[28] 张 萍,金国庆,周志春,等. 木荷苗木性状的种源变异和地理模式[J]. 林业科学研究,2004,17(2):192-198.
猜你喜欢
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
现代农业科技(2017年24期)2018-01-22 21:35:06
防护林科技(2016年9期)2016-09-27 01:34:21
广西林业科学(2016年2期)2016-03-20 05:53:21
广西林业科学(2016年3期)2016-03-16 05:43:22
广东海洋大学学报(2015年4期)2016-01-13 08:39:30
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
首都医科大学学报(2015年4期)2015-12-16 13:00:08
