
蝉花居群形态变异式样
2018-05-31 08:14:09李建平曾文波
中国食用菌 2018年3期
李建平,张 铁,曾文波*
(1.文山学院环境与资源学院,云南 文山 663099;2.文山学院文山州生物资源开发研究中心,云南 文山 663099)
虫草类真菌Cordyceps s.l.感染寄主后形成虫菌复合体,包括大多数虫草、菌菌复合体(如大团囊虫草、头状虫草和分枝虫草等)[1],以及重寄生(多头霉等可寄生于其他虫草菌上)[2],虫草类真菌形态独特,昆虫等被真菌感染后僵死,真菌利用寄主体内营养长成子座或孢梗束突出于寄主表面,完整的子实体包括虫体部分(菌核)和一段似草的部分子座(或孢梗束),因此人们形象地将之称为虫草。
“蝉花”(Isaria cicadae)是我国名贵传统中药材,最早关于“蝉花”的记载是成书于公元五世纪的《雷公炮炙论》[3],现代研究表明,蝉花为真菌Isaria cicadae Miquel感染蝉若虫后形成的虫菌复合体[4],分布广泛,包括中国、韩国、日本、澳大利亚、新西兰、巴西、南亚、欧洲、北美等国家和地区[4-5],具有多种药理作用,在我国的药用历史超过1 500年,可作为冬虫夏草的代用品[3-4],蝉花是具有巨大开发价值的野生资源之一。
目前,关于虫草类真菌的形态特征研究多是关于虫草子座、菌核、子囊结构[6-12],以及菌丝体形态[4]的描述,关于虫草居群形态学的报道很少,本文以居群为单位,研究蝉花的孢梗束、菌核形态在居群间和居群内及个体间形态变异式样。从形态角度探讨蝉花状变异式样与生态适应特征,揭示其分化特点,为蝉花资源的开发利用提供基础数据。
1 材料和测定方法
1.1 材料
用于观察的材料为新鲜虫草,包括蝉花10个居群共100个个体,冬虫夏草(Ophiocordyceps sinensis)5个居群共50个个体,详见表1。
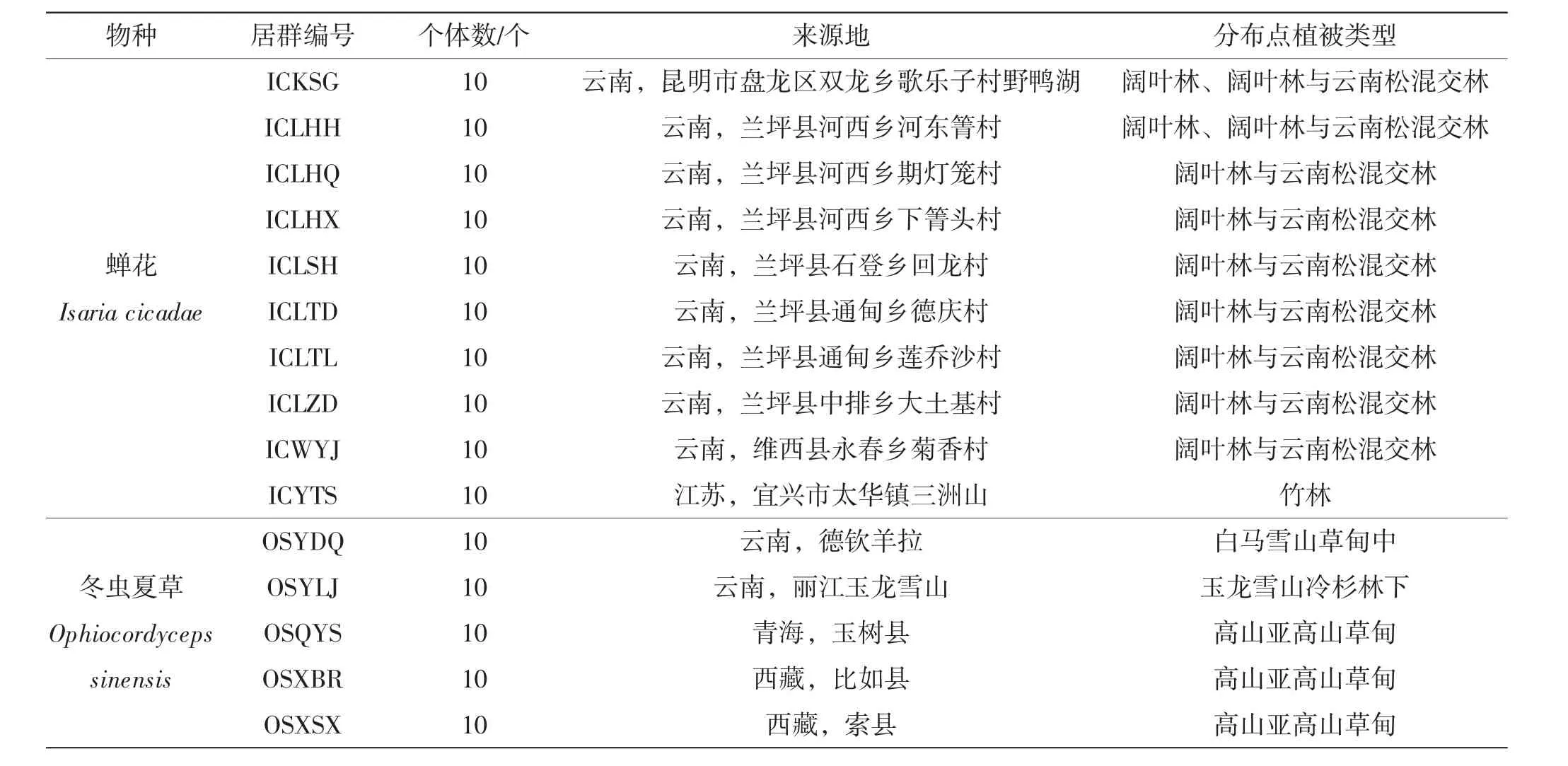
表1 10个居群蝉花和5个居群冬虫夏草来源Tab.1 Locality of 10 populations of Isaria cicadae and 5 populations of Ophiocordyceps sinensis
1.2 形态性状测定方法
采用生物统计方法[13],每个居群随机选取新鲜样品10个个体,按虫草不同部位(菌核部分,孢梗束或子座部分),分别测量其重量、周长、长、分枝数等性状,其中,重量为样品干燥后测定值,每个性状测量3次,取平均值,所测性状及其对应的编号见表2,其中,由于蝉花孢梗束分枝较多,选取一级分枝最长的3枝测量,对于达不到3枝的个体,缺失分枝长BC和分枝周长BZ两个性状数值记为“0”。

表2 蝉花和冬虫夏草观察性状、编号Tab.2 Number of morphological traits of Isaria cicadae and Ophiocordyceps sinensis
1.3 数据分析方法
1.3.1 形态性状基本统计量
使用SAS 8.1软件计算蝉花和冬虫夏草形态性状的平均值、标准差、最大值、最小值和变异系数。标准差StdDev表示形态性状值在平均值上下波动大小,标准差越大则反映形态性状值离平均值波动越远;变异系数CV(标准差除以平均值)表示表型性状值离散性程度,变异系数越大则表型性状值离散程度越大,反映形态性状值在居群中的整齐性程度[14]。
1.3.2 形态性状巢式方差分析
形态性状巢式等级方差分析[15-18]的模型为:

式中:Yijkl为第i个居群第j个单株第k次重复第l次的观测值;μ为总体平均值;Si为第i个居群的效应(固定);T(i)j为第i个居群第j个单株的效应(随机);R(ij)k为第i个居群第j个单株第k次重复的效应(固定);ε(ijk)l=ε(ijk)=试验误差。
为进一步了解蝉花形态性状在居群间和个体间的分配情况,利用SAS 8.1进行巢式方差分析,使用的SAS程序[18]如下:
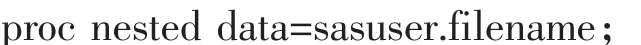
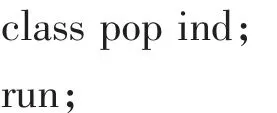
1.3.3 形态性状Q-聚类分析
采用AVE聚类法,对10个居群蝉花样品形态性状的平均值进行居群间聚类分析,使用的SAS程序[18]如下:

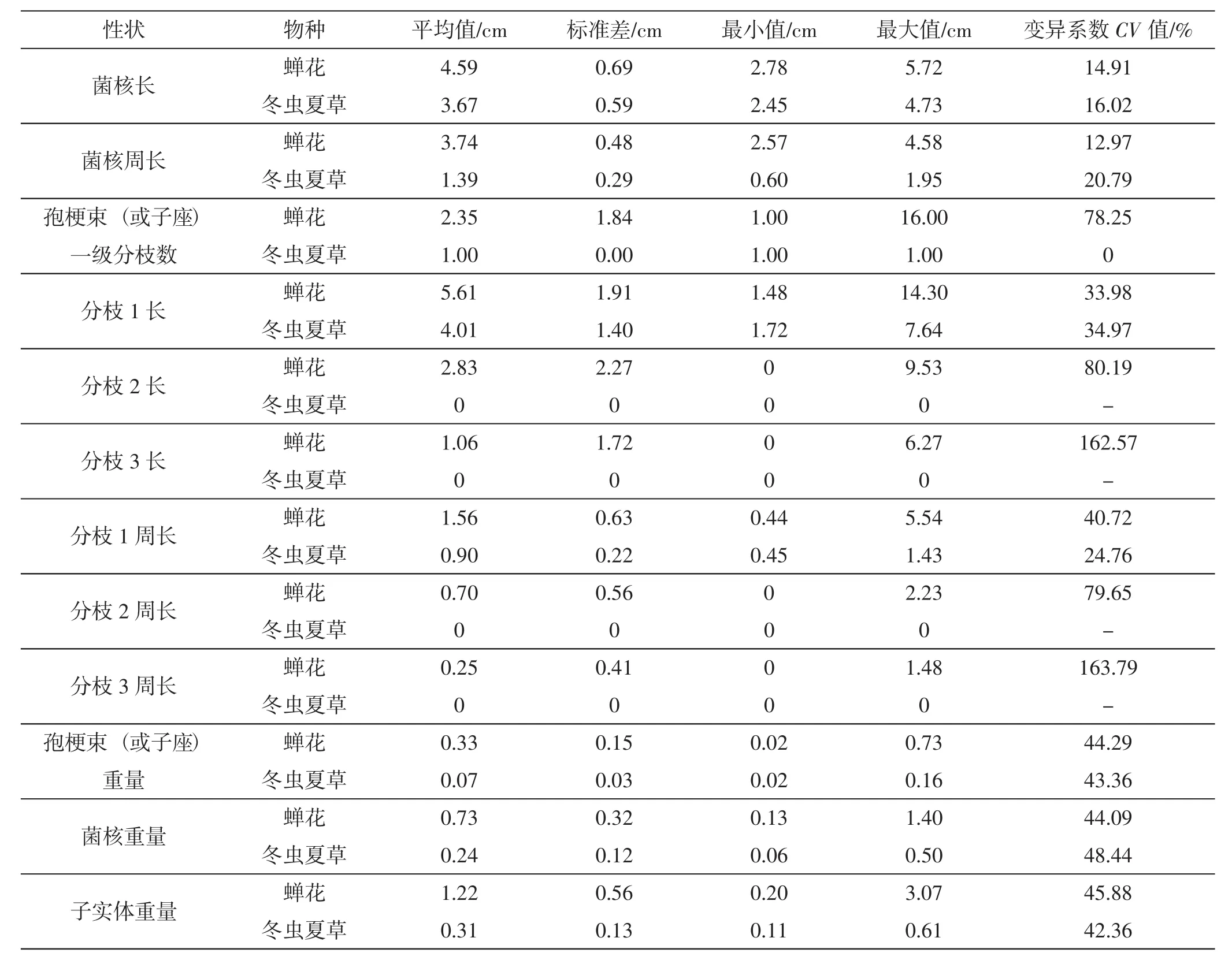
表3 蝉花与冬虫夏草物种水平形态性状分析结果Tab.3 The analysis results of morphologic traits of Isaria cicadae and Ophiocordyceps sinensis at species level
2 结果与分析
2.1 形态特征基本统计量
变异系数可表示表型性状值离散性的程度,变异系数越大,则性状值离散程度越大,反之则小。蝉花和冬虫夏草物种水平形态性状统计结果见表3。
10个居群的蝉花,其菌核长(JC)平均值在3.18 cm~5.15 cm 之间,居群水平 CV 值为 5.93%~10.56%,物种水平 CV值为 14.91%;菌核周长(ZC)平均值在2.98 cm~4.16 cm之间,居群水平CV值为 6.07%~9.97%,物种水平 CV 值为 12.97%,Pop ICKSG的菌核长和周长最小,Pop ICYTS的菌核长和周长最大,Pop ICYTS的寄主为竹蝉,Pop ICKSG的寄主是鸣鸣蝉,Pop ICWYJ的寄主是云南黑蝉,表明蝉花的菌核长和周长主要是与寄主直接相关的,菌核长和菌核周长这两个形态性状的CV值较小,说明这两个性状稳定,变异不大。蝉花菌核重量(JZ)平均值在 0.20 g~0.95 g 之间,CV 值为 17.35%~36.63%;孢梗束重量 (ZZ)平均值在 0.09 g~0.44 g之间,CV值为13.40%~45.79%,表明不同产地的蝉花的菌核和孢梗束重量有较大的差异。蝉花孢梗束一级分枝数(BR)平均值在1.7枝~3.9枝之间,CV值为38.83%~114.32%,表明蝉花孢梗束多分枝,孢梗束的分枝数变异大。分枝1长(BC-1)平均值在3.05 cm~7.19 cm 之间,分枝 1 周长 (BZ-1)平均值在 1.26 cm~2.31 cm 之间;分枝 2 长 (BC-2)平均值在 1.91 cm~5.36 cm 之间,分枝 2 周长 (BZ-2)平均值在 0.38 cm~0.90 cm 之间;分枝 3 长 (BC-3)平均值在 0.50 cm~2.19 cm 之间,分枝 3周长 (BZ-3)平均值在0.06 cm~0.38 cm之间,蝉花12个形态性状的变异系数(物种水平),分枝3周长(BZ-3)最大,CV值为163.79%,其次为分枝3长度(BC-3),CV值为162.57%,由于个体间分枝数目不一致,使得分枝相关的形态性状变异最大。
除了与孢梗束分枝数相关性状以外,蝉花孢梗束重量的变异系数最大(45.88%);其次是子实体重量QZ(44.29%);不同居群内每一表型性状的变异系数均有一定差异,说明蝉花居群间由于遗传和环境异质性导致居群表型变异存在差异。所研究10个蝉花居群的12个形态性状中,与孢梗束分枝数相关的形态性状变异系数值均偏大,说明这些性状在居群内个体间分化较大,多态性丰富,这可能与其适应生长环境有关。
冬虫夏草5个居群8个形态性状的变异系数(物种水平)在16.02%~48.44%之间。这些形态性状中菌核重量的变异系数最大(48.44%),不同产地冬虫夏草的大小和重量等形态性状,主要与冬虫夏草寄主的种类和被感染年龄有关,子座一级分枝数(BR)的变异系数为0,表明该性状较其他性状稳定,冬虫夏草子座多单生。
2.2 巢式方差分析
巢式方差分析可以揭示居群间和居群内的变异,而且可以划分居群间和居群内变异组分。蝉花形态性状的表型分化系数(Vst)见表4。
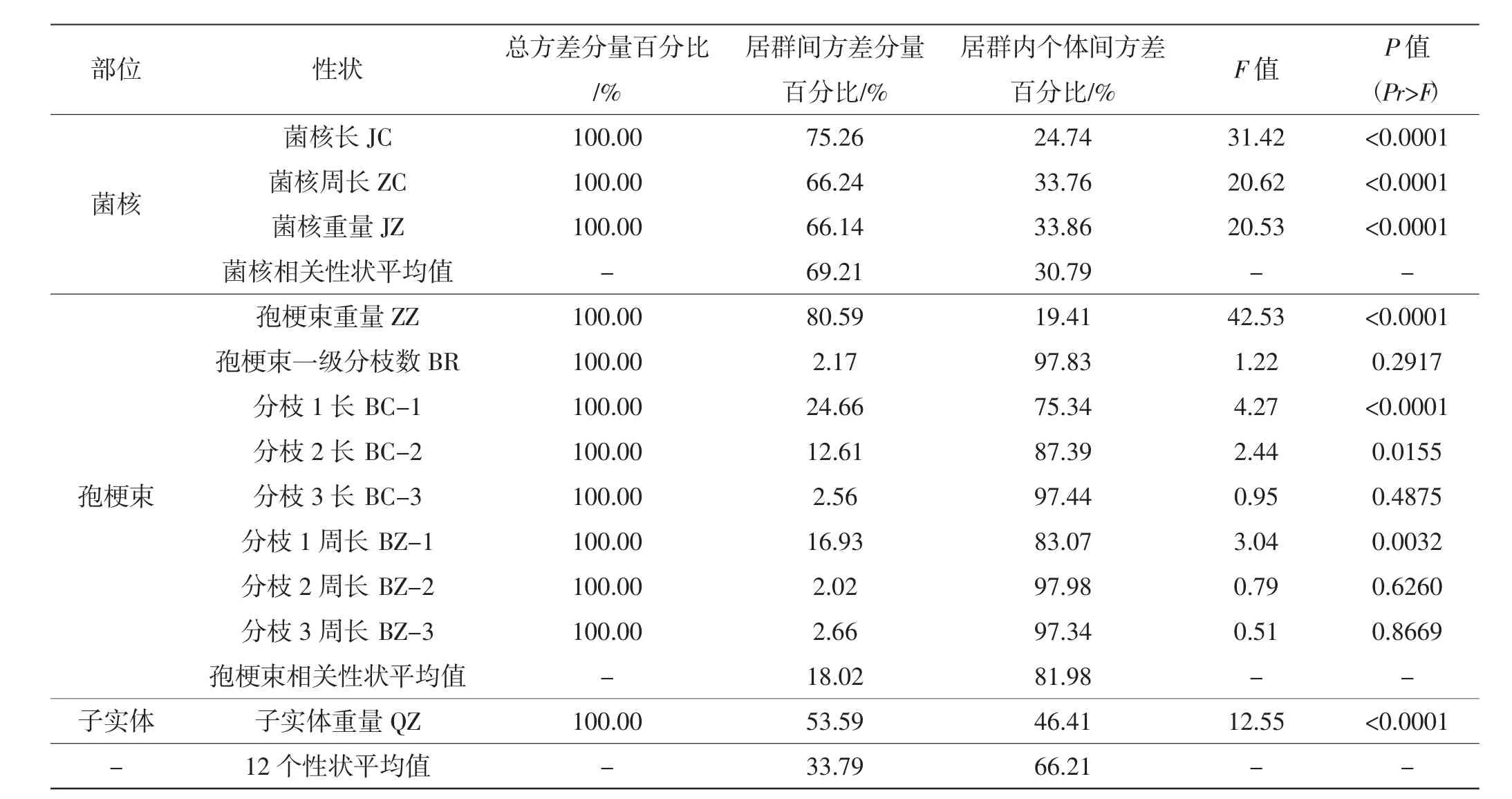
表4 蝉花10个居群12个性状巢式方差分析Tab.4 Nested variation analysis of 12 morphological traits among 10 populations of Isaria cicadae
从表4可知,蝉花菌核部分12个形态性状在居群间的平均方差分量占33.79%,居群内个体间的平均方差分量占总变异的66.21%,居群内个体是形态性状变异的主要来源。其中,菌核部分3个形态性状在居群间的平均方差分量占69.21%,居群内个体间的平均方差分量占总变异的30.79%,居群间的多态性大于居群内的多态性;蝉花孢梗束部分7个形态性状在居群间的平均方差分量占18.02%,居群内个体间的平均方差分量占总变异的81.98%,居群内的多态性远大于居群间的多态性,与孢梗束分枝等相关的形态性状的差异则主要来自于个体间,同一居群不同个体的孢梗束形态性状差异巨大,孢梗束的生长除受到蝉花本身的遗传因素影响,在生长过程中环境因素的影响较大,这使得居群内个体间的孢梗束形态变异较菌核相关形态性状丰富。
冬虫夏草12个性状中,由于测量个体的分枝数均为1枝,有效的形态性状数据有7个,详见表5。

表5 冬虫夏草5个居群7个性状巢式方差分析Tab.5 Nested variation analysis of 7 morphological traits among 5 populations of Ophiocordyceps sinensis
从表5可以看出,这七个性状均表现为居群间方差分量大于个体间方差分量,表明居群是这些形态性状的主要来源,居群间形态有较大的分化。冬虫夏草形态性状在居群间与居群内的分布式样与蝉花不一样,冬虫夏草形态性状在居群间的分化大于蝉花居群间的分化。
2.3 Q-聚类分析
10个蝉花居群17个形态性状AVE聚类法的谱系聚类图见图1。

图1 10个蝉花居群17个形态性状AVE聚类法的谱系聚类图Fig.1 Q-Cluster of 17 morphological traits assayed in 10 populations of Isaria cicadae with average linkage method
由图1可知,10个居群的蝉花聚成CladeA和CladeB两支,在CladeB中包括9个居群的蝉花,CladeA中包括1个居群(Pop ICKSG)的蝉花。在CladeB中,产自兰坪县的7个居群(Pop ICLHH、I CLHQ、ICLZD、ICLSH、 ICLTL、 ICLHX、ICLTD)先聚成一支,再依次与Pop ICWYJ和Pop ICYTS聚在一起,聚类图在一定程度上与地理呈相关性,表明不同居群的蝉花的形态性状具有地域性特征。
3 讨论
虫草菌核的大小和重量等性状主要是受到寄主大小的影响,蝉花的寄主为同翅目的昆虫蝉,现已报道关于蝉花的寄主蝉共有8种[19],而冬虫夏草的寄主为蝙蝠蛾,已报道的种类有57种[20],冬虫夏草寄主的种类远远多于蝉花的寄主,这可能是冬虫夏草形态性状居群间分化大于蝉花的原因之一。另外一个可能的原因是:人们在巨大的经济利益驱动下,对冬虫夏草进行破坏性的采集,导致冬虫夏草居群分布点的范围不断缩小,这阻碍了冬虫夏草居群间的基因交流,使得冬虫夏草居群间分化进一步加大。由于寄主的种类直接决定蝉花和冬虫夏草的菌核大小,与菌核大小直接相关的性状,如菌核长(JC)、菌核周长(ZC)、菌核重量(JZ)等性状的变异要小(CV值较小),但受到不同产地的寄主种类和分化的影响,使得菌核大小直接相关的性状在居群间形成分化。与孢梗束相关的形态性状,如孢梗束一级分枝数(BR)、分枝1长(BC-1)、分枝 2长 (BC-2)、分枝3长(BC-3)、分枝1周长(BZ-1)、分枝3周长(BZ-3)、分枝3周长(BZ-3)则变异较大。同一居群内不同个体的孢梗束差异较大,可能是由于不同的个体在遗传上的差异导致的,在培养蝉棒束孢菌丝体的过程中还发现,不同的菌株在相同的培养条件下,其产孢子、产菌丝、产孢梗束的能力差异明显,表明其在遗传结构上可能具有较大的差异,有必要开展蝉花遗传结构的研究;另外,蝉花孢梗束的形成还受到其生长环境的影响,如菌核距地表的距离、地表的枯枝落叶、地表植被等。总之,遗传上的差异和生境的多样性是蝉花孢梗束形态变异丰富的来源。丰富的变异一定程度上也反映了蝉花具有较强的适应环境能力,这与蝉花分布广、适应性强[4]的实际情况是一致的。
参考文献:
[1]梁宗琦.中国真菌志(第三十二卷虫草属)[M].北京:科学出版社,2007.
[2]王元兵.昆明地区下垂虫草生物学研究[D].昆明:云南大学,2012.
[3]李冰岚.蝉花的本草学考证[J].现代应用药学,1993,10(2):21-22.
[4]曾文波,常衬心,李建平,等.蝉棒束孢显微形态变异样[J].微生物学报,2017,57(3):350-362.
[5]曾文波,王元兵,虞泓,等.蝉棒束孢居群分布及生境调查[J].中国食用菌,2015,34(4):50-56.
[6]刘爱英,梁宗琦.中国虫草属一新记录种[J].真菌学报,1993,12 (3):248-250.
[7]梁宗琦,刘爱英,刘作易.冬虫夏草子囊孢子的发育[J].真菌学报,1995,14(2):148-152.
[8]梁宗琦,刘爱英,黄建忠,等.宽阔水自然保护区的虫草及其相关真菌I[J].真菌学报,1996,15(4):264-271.
[9]梁宗琦,刘爱英,黄建忠,等.宽阔水自然保护区的虫草及其相关真菌II[J].菌物系统,1997,16(1):61-67.
[10]刘作易.虫草属及其无性型关系研究[D].武汉:华中农业大学,1999.
[11]臧穆.我国热带真菌两新种[J].云南植物研究,2001,23(3):295-297.
[12]杨钟林,虞泓,陈自宏,等.滇中蛹虫草居群生物学及其生态学习性研究[J].中国食用菌,2011,30(5):43-47.
[13]杜荣骞.生物统计学[M].北京:高等教育出版社,1990.
[14]高惠璇.实用统计方法与SAS系统[M].北京:北京大学出版社,2001.
[15]葛颂,洪德元,钱迎倩.遗传多样性及其检测方法,生物多样性研究的原理与方法 [M].北京:中国科学技术出版社,1994.
[16]虞泓,黄瑞复.云南松居群核型变异及其分化研究[J].植物分类学报,1998,36(3):222-231.
[17]虞泓,杨彩云,徐正尧.云南松居群花粉形态多态性[J].云南大学学报:自然科学版,1999,21(2):86-89.
[18]何显静,王兵益,虞泓.SAS系统在川百合形态变异式样研究中的运用[J].云南大学学报:自然科学版,2003,25(增刊):75-77.
[19]陈祝安,李增智,陈以平.金蝉花[M].北京:中医古籍出版社,2014.
[20]Wang XL,Yao YJ.Host insect species of Ophiocordyceps sinensis:a review[J].ZooKeys,2011(127):43-59.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
贵州科学(2021年3期)2021-07-09 12:19:28
西北农业学报(2021年5期)2021-06-28 07:42:02
中国食用菌(2021年1期)2021-03-12 06:33:30
军事文摘(2020年22期)2021-01-04 02:16:52
草地学报(2018年5期)2018-11-07 02:25:00
山西农业科学(2017年3期)2017-04-14 07:37:52
小学生必读(高年级版)(2016年9期)2016-12-12 06:03:44
汉语世界(The World of Chinese)(2013年5期)2013-04-29 00:44:03
