
Cd胁迫对不同砧穗组合葡萄植株光合作用、膜脂过氧化和抗氧化酶活性的影响
2018-05-17 02:42:57李小红李辉信任俊鹏李运景吴亚蓓马晓青陶建敏
河南农业科学 2018年3期
李小红,李辉信,任俊鹏,李运景,吴亚蓓,马晓青,陶建敏
(1.河南省农业科学院 农业经济与信息研究所,河南 郑州 450002; 2.南京农业大学 资源与环境科学学院,江苏 南京 210095; 3.江苏丘陵地区镇江农业科学研究所,江苏 句容 212400;4.南京农业大学 园艺学院,江苏 南京 210095)
目前,由于工业和农业的快速发展,人类的各种活动比如采矿、车辆运载、污水灌溉、化肥施用等引起的重金属污染问题日益严重,研究表明,我国耕地重金属污染面积占耕地总面积的16.7%左右,Cd污染概率为25.2%,远超过Ni、Hg、As和Pb等其他几种重金属[1-3]。Cd不是植物生长发育的必需元素,但是易于被植物吸收并累积,对植物的生长发育产生一系列的影响,比如抑制植物生长、降低植物光合作用、干扰植物矿质营养吸收,导致作物产量降低和品质下降[4-13]。Cd积累不仅对植物生长造成影响,并且能够沿食物链富集进入人体,引起身体机能失调和一系列疾病,比如痛痛病、癌症和中毒性肾损伤[14-15]。Cd作为毒害较强且污染严重的重金属,引起了国内外学者的关注和研究。
果农为了提高果品质量和产量,长期使用农药、化肥,加之不当的农业生产方式,造成许多果园土壤质量下降,微生物数量减少,甚至有的果园土壤还出现了重金属累积[16-18]。果园土壤被Cd污染后,轻者阻碍果树的生长和发育,影响果品的产量和质量,严重者导致果品富集Cd,并通过食物链进入人体,给人类健康带来潜在的风险。
葡萄生产中,通过嫁接可以调整栽培品种的生长势、成熟期、产量和果实品质[19-21]。大量研究发现,嫁接还能够提高果树和蔬菜等园艺作物对逆境胁迫如水分胁迫、盐分胁迫、有机物和重金属污染[22-25]的适应能力。但是,关于嫁接对重金属胁迫下果树生长、膜脂过氧化产物的累积及抗性机制方面的影响研究较少,而关于Cd胁迫条件下嫁接对葡萄植株影响的研究还未见报道。因此,以不同的葡萄砧木嫁接生产上的优良早熟葡萄品种矢富罗莎,研究不同砧穗组合在Cd胁迫下的生长和生理响应差异,以期为葡萄抗性砧木的选择提供初步理论支撑,并为葡萄安全、优质生产提供参考依据。
1 材料和方法
1.1 供试土壤和葡萄植株材料
试验在南京农业大学沧波门葡萄实验基地进行。选取7个葡萄砧木品种(巨峰、5BB、110R、99R、华佳8号、8B、SO4)作为试验材料,于3月中旬将生长相对一致的7个品种砧木苗和矢富罗莎扦插苗定植于塑料盆(35 cm×30 cm)中,待砧木苗生长至3~4片叶时,采用绿枝嫁接矢富罗莎,分别记作Y/巨峰、Y/5BB、Y/110R、Y/99R、Y/华佳8号、Y/8B和Y/SO4,以矢富罗莎扦插苗为对照。
盆土所用土壤来自南京农业大学沧波门葡萄试验基地,土壤类型为黄棕壤,pH值6.8,有机碳含量为6.86 g/kg,阳离子交换量为10.79 cmol/kg,速效磷含量为16.2 mg/kg,Cd的背景值为0.56 mg/kg。
1.2 试验处理与设计
1.2.1 污染浓度的确定 预备试验设置4个Cd水平,分别为2.5、5、15、25 mg/kg,以未添加Cd土壤为对照,共5个处理。试验结果表明:25 mg/kg Cd处理水平下,葡萄植株生长迟缓、叶片变小、新梢节间变短,葡萄果粒小,不能正常着色,有僵果,严重影响树体生长、果实正常发育和成熟;2.5、5 mg/kg 2个Cd处理水平下,葡萄植株生长状况良好,与对照差异不明显;15 mg/kg Cd处理水平下,葡萄植株生长状况与对照相比,明显受到影响,但未严重阻碍植株生长,果实能够正常发育和成熟,由于试验材料有限,最终确定Cd 15 mg/kg作为试验处理。
1.2.2 试验处理 在风干过筛后的土壤中加入CdCl2溶液,以模拟成15 mg/kg的Cd污染土壤,在常温下熟化平衡2个月,然后装入塑料盆,将3年生矢富罗莎葡萄苗栽入塑料盆中,每个处理重复4次。采用避雨栽培,常规土肥水管理和病虫害防治。
1.3 测定指标与方法
1.3.1 光合参数的测定 采用Li-6400(美国LI-COR公司生产)光合测定仪,于5月31日(晴天)9:00—10:00进行测定,每个处理均选择3株生长基本一致、无病虫害的植株,取新梢部位第3~5节位的功能叶片进行净光合速率(Pn)、蒸腾速率(Tr)和胞间CO2浓度(Ci)的测定。
1.3.2 叶绿素和丙二醛(MDA)含量的测定 叶绿素含量的测定采用丙酮提取法[26]:称取0.1 g葡萄叶片,加入5 mL 80% 的丙酮溶液在黑暗条件下浸提,时间为48 h。MDA含量的测定采用硫代巴比妥酸法[27]。
1.3.3 酶活性的测定 超氧化物歧化酶(SOD)的活性采用氮蓝四唑(NBT)光还原法测定,过氧化物酶(POD)的活性采用愈创木酚法测定,过氧化氢酶(CAT)的活性通过单位时间内H2O2含量的变化来表示。
1.4 统计分析
试验数据采用SPSS for Windows 13.0软件和EXCEL进行统计分析,显著性差异水平为P<0.05。
2 结果与分析
2.1 不同砧木嫁接对Cd胁迫下矢富罗莎葡萄叶片光合参数的影响
表1显示,Cd胁迫下,矢富罗莎不同砧穗组合葡萄叶片的光合参数存在差异。嫁接苗葡萄叶片的Pn均高于自根苗,Y/巨峰和Y/华佳8号组合略高于对照自根苗,与对照差异不显著,以Y/99R和Y/8B组合较高,分别为9.62、7.09 μmol/(m2·s),分别是对照自根苗的4.74、3.49倍;矢富罗莎不同砧穗组合葡萄叶片的Tr存在显著差异,以Y/99R和Y/110R组合较高,分别为2.84、1.47 mmol/(m2·s);除Y/SO4组合外,所有嫁接组合葡萄叶片的Ci均显著高于对照,其中以Y/99R和Y/巨峰组合最高,分别为对照的2.96、2.73倍。嫁接减轻了Cd胁迫对葡萄植株的光合作用和蒸腾作用的抑制程度,细胞间隙CO2供应也更为充足。

表1 不同砧木嫁接对Cd胁迫下葡萄叶片光合参数的影响
注:同列不同小写字母表示差异显著(P<0.05),下同。
2.2 不同砧木嫁接对Cd胁迫下矢富罗莎葡萄叶片叶绿素和MDA含量的影响
Cd处理下,矢富罗莎不同砧穗组合葡萄的叶绿素含量存在差异(图1)。以Y/巨峰、Y/华佳8号和Y/5BB嫁接组合的叶绿素含量较高,比对照自根苗叶片叶绿素含量分别提高了17.9%、14.9%和14.2%,Y/110R、Y/8B和 Y/SO4组合居中,比对照自根苗叶片叶绿素含量分别提高了9.7%、9.0%和7.5%,Y/99R略高于对照,与对照差异不显著。这说明嫁接有效减轻了Cd胁迫对葡萄叶片光合色素的破坏。此外,与对照自根苗相比,嫁接减轻了Cd胁迫对葡萄植株叶片的膜脂过氧化损伤程度,降低了葡萄植株叶片的MDA 含量,所有嫁接组合叶片的MDA 含量均显著低于对照,为对照葡萄叶片含量的35.0%~80.8%,其中以 Y/110R的下降幅度最大,为对照植株葡萄叶片含量的35.0%(图2)。
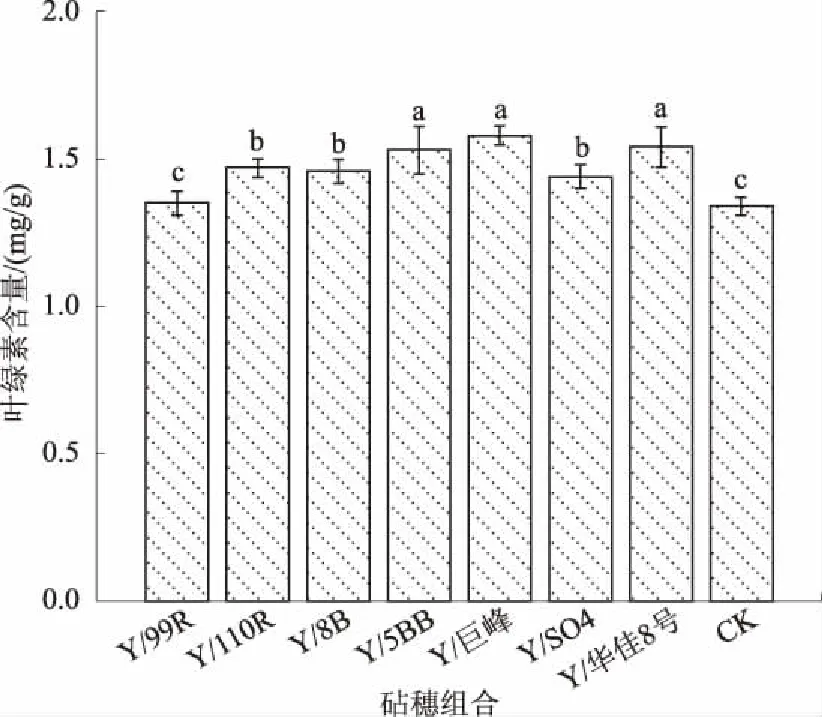
不同小写字母表示差异显著(P<0.05),图2同

图2 不同砧木嫁接对Cd胁迫下葡萄叶片MDA含量的影响
2.3 不同砧木嫁接对Cd胁迫下矢富罗莎葡萄叶片抗氧化酶活性的影响
由表2可以看出,Cd胁迫下矢富罗莎不同砧穗组合葡萄叶片的抗氧化酶活性不同,Y/99R、Y/5BB组合葡萄叶片的SOD和CAT活性分别为9.35 U/g和 13.27 U/(g·min)、8.42 U/g和 9.92 U/(g·min),均明显高于对照和其他组合,POD活性以Y/110R、Y/99R和 Y/8B较高,Y/华佳8号和Y/5BB居中,Y/巨峰和Y/SO4 最低。除Y/110R、Y/8B、Y/巨峰和Y/华佳8号组合的SOD活性低于对照或与对照差异不显著外,其他嫁接组合的抗氧化酶(SOD、POD和CAT)活性均显著高于对照自根苗。

表2 嫁接对Cd胁迫下葡萄叶片抗氧化酶活性的影响
3 结论与讨论
光合作用作为植物体内极为重要的生理过程,也是植物对环境变化最为敏感的代谢过程,其强弱与植物生长发育、产量和抗逆性有着十分密切的关系,因而可用作判断植物生长状况和抗逆性强弱的指标[28]。有学者研究发现,嫁接作为提高植物抗逆性的有效措施之一,能够提高甜瓜幼苗对铜胁迫的耐性,减轻对甜瓜幼苗叶片光合色素的毒害,从而提高光合作用[29]。本研究表明,嫁接减弱了Cd胁迫对葡萄植株叶片光合色素的损伤程度,除Y/99R与对照自根苗差异不显著以外,其他嫁接组合葡萄植株叶片的叶绿素含量均显著高于自根苗。同时本研究结果显示,葡萄嫁接苗在Cd胁迫下表现出较高的净光合速率和蒸腾速率,细胞间隙CO2浓度也明显高于自根苗,不同砧穗组合之间存在差异,其中以Y/99R、Y/110R和Y/5BB组合的较高。所以,嫁接减轻了Cd胁迫对葡萄叶片叶绿素含量、光合作用的损伤和抑制程度,从而减轻了对葡萄植株的毒害作用。
Cd胁迫能够引起植物体内活性氧自由基的过量产生,对植物细胞产生毒害作用,从而导致细胞膜脂过氧化作用,MDA是膜脂过氧化的主要产物之一,可与膜上蛋白质结合,导致蛋白质分子间和分子内产生交联,严重损失和破坏细胞膜系统,MDA含量高低反映了膜脂的损伤程度[30-31]。研究表明,嫁接能够减弱铜胁迫对甜瓜幼苗的膜脂过氧化损伤,减少MDA含量的累积[32]。本试验的研究结果与此一致,在Cd胁迫条件下,嫁接葡萄苗叶片的MDA含量明显低于自根苗,这表明嫁接葡萄苗膜脂过氧化的程度比自根苗轻,嫁接能够减弱Cd胁迫对葡萄植株的膜脂过氧化损伤。
正常情况下植物体细胞内自由基的产生和清除处于动态平衡状态,但是当植物受到Cd胁迫时,超氧阴离子产生速率升高、过氧化氢含量增加,引起植物细胞活性氧爆发,平衡被打破,最终使植物受到了氧化损伤[33-34]。SOD、POD和CAT等抗氧化酶类作为植物细胞中内源活性氧的清除剂,能够在逆境胁迫中清除过量的活性氧自由基,维持细胞内的活性氧的代谢平衡,减轻对膜结构的破坏,从而在一定程度上减缓了逆境胁迫对植物的伤害[35]。本试验结果显示,在Cd胁迫条件下,所有嫁接组合的POD和CAT活性都显著高于自根苗,这表明嫁接增强了葡萄植株对活性氧的清除能力,减轻了对质膜的氧化损伤,从而提高了葡萄植株对Cd胁迫的耐受性。这与谭明明[29]在甜瓜上的研究结果一致。此外,本试验中,Y/8B、Y/巨峰和Y/华佳8号葡萄植株的SOD活性低于自根葡萄植株,这可能是由于不同组合葡萄植株的抗氧化机制存在差异,Y/8B和Y/华佳8号表现出较高的POD活性。
研究发现,甜瓜不同砧木嫁接苗在铜胁迫下抗氧化酶活性大小存在差异[29],本试验也得出了一致的结果,在Cd胁迫条件下,不同组合葡萄植株体内的SOD、POD和CAT活性均存在差异。研究表明,作物对Cd的耐性具有明显的种间和种内差异[36],Cd耐性植物种间和种内差异与体内抗氧化能力有关,本试验结果表明,在Cd胁迫条件下,Y/99R 和Y/5BB 组合的SOD 和 CAT活性显著高于其他组合,这与之前文献报道的Y/99R 和Y/5BB 组合Cd积累浓度较低相一致[37]。
参考文献:
[1] Yousaf B,Liu G,Wang R,etal.Bioavailability evaluation,uptake of heavy metals and potential health risks via dietary exposure in urban-industrial areas[J].Environmental Science and Pollution Research,2016,23:22443-22453.
[2] 罗小玲,郭庆荣,谢志宜,等.珠江三角洲地区典型农村土壤重金属污染现状分析[J].生态环境学报,2014,23(3):485-489.
[3] 宋伟,陈百明,刘琳.中国耕地土壤重金属污染概况[J].水土保持研究,2013,20(2):293-298.
[4] Gharaibeh M A,Marschner B,Heinze S.Metal uptake of tomato and alfalfa plants as affected by water source,salinity,and Cd and Zn levels under greenhouse conditions[J].Environmental Science and Pollution Research,2015,22:18894-18905.
[5] Kuma V,Chopra A K,Srivastava S.Assessment of heavy metals in spinach (SpinaciaoleraceaL.) grown in sewage sludgeeamended soil[J].Communications in Soil Science and Plant Analysis,2016,47:221-236.
[6] Bertoli A C,Gabriel C M,Carvalho R,etal.Lycopersicon esculentum submitted to Cd-stressful conditions in nutrition solution:Nutrient contents and translocation[J].Ecotoxicology and Environmental Safety,2012,86:176-181.
[7] 李萌,解静芳,吕佳莉,等.污水与镉复合胁迫对玉米幼苗抗氧化酶活性的影响[J].山西农业科学,2017,45(2):172-177.
[8] 刘强,呼丽萍,鱼潮水,等.樱桃种植区土壤及樱桃重金属富集状况研究[J].河南农业科学,2017,46(10):60-65.
[9] 牟明辉,石杨程,张晓晴,等.恩施富硒茶园土壤重金属和氟含量及风险评价[J].河南农业科学,2016,45(5):61-65.
[10] 张晶,王姣爱,党建友,等.Cd,Ni,Pb对小麦灌浆期叶绿素荧光参数的影响[J].山西农业科学,2016,44(10):1455-1458.
[11] 张红岩,马丽,张伯阳,等.Fe2+、Cd2+互作对小麦幼苗生长发育的影响[J].河南农业科学,2014,43(8):20-24.
[12] 蔡卓,卢登峰,何晓良,等.重金属镉在芦荟体内的积累及对芦荟抗氧化酶活性的影响[J].南方农业学报,2013,44(2):210-213.
[13] 孙清斌,尹春芹,邓金锋,等.矿区农田土壤小麦体系中重金属污染特征研究[J].河南农业科学,2013,42(4):80-84.
[14] Chaney R L.How does contamination of rice soils with Cd and Zn cause high incidence of human Cd disease in subsistence rice farmers[J].Current Pollution Reports,2015,1:13-22.
[15] Khan A,Khan S,Alam M,etal.Toxic metal interactions affect the bioaccumulation and dietary intake of macro- and micro-nutrients[J].Chemosphere,2016,146:121-128.
[16] 李晓彤,岳田利,胡仲秋,等.陕西省猕猴桃园土壤重金属含量及污染风险评价[J].西北农林科技大学学报(自然科学版),2015,43(2):173-178.
[17] 张连忠,路克国,王宏伟,等.重金属和生物有机肥对苹果根区土壤微生物的影响[J].水土保持学报,2005,19(2):92-95.
[18] Li J T,Qiu J W,Wang X W,etal.Cadmium contamination in orchard soils and fruit trees and its potential health risk in Guangzhou,China[J].Environmental Pollution,2006,143:159-165.
[19] 李小红,周凯,谢周,等.不同葡萄砧木对矢富罗莎葡萄嫁接苗光合作用的影响[J].果树学报,2009,26(1):90-93.
[20] 蒋爱丽,李世诚,杨天仪,等.不同砧木对藤稔葡萄生长与果实品质的影响[J].上海农业学报,2005,21(3):73-75.
[21] 程建徽,梅军霞,郑婷,等.不同砧木对欧亚种葡萄红亚历山大产量和品质的影响[J].核农学报,2015,29(8):1607-1616.
[22] Cohen S,Naor A.The effect of three rootstocks on water use,canopy conductance and hydraulic parameters of apple trees and predicting canopy from hydraulic conductance[J].Plant Cell Environment,2002,25(1):17-28.
[23] Edelstein M,Ben-Hur M,Cohen R,etal.Born and salinity effects on grafted and non-grafted melon plants [J].Plant Soil,2005,269:273-284.
[24] Otani T,Seike N.Comparative effects of rootstock and scion on dieldrin and endrin uptake by grafted cucumber(Cucumissativus)[J].Journal of Pesticide Science,2006,31:316-321.
[25] Zhang Z K,Li H,He H J,etal.Grafting raises the Cu tolerance of cucumber through protecting roots against oxidative stress induced by Cu stress[J].Journal of Integrative Agriculture,2013,12(5):815-824.
[26] Hegedüs A,Erdei S,Horváth G.Comparative studies of H2O2detoxifying enzymes in green and greening barely seedlings under cadmium stress[J].Plant Science,2001,160:1085-1093.
[27] Liu P C,Wang H,Cheng J Q,etal.Regulation of nitric oxide on drought-induced membrane lipid peroxidation in wheat leaves [J].Acta Botanica Boreali-occidentalia Sinica,2004,24:141-145.
[28] 张永平,范红伟,杨少军.外源水杨酸对Cd胁迫下甜瓜幼苗生长、光合作用和活性氧代谢的缓解效应[J].植物生理学报,2014,50(10):1555-1562.
[29] 谭明明.嫁接提高甜瓜幼苗对铜胁迫耐受性的生理机制研究[D].雅安:四川农业大学,2014.
[30] 郭天荣,陈丽萍,冯其芳,等.Al、Cd胁迫对空心菜生长及抗氧化特性的影响[J].核农学报,2015,29(3):571-576.
[31] Shamsi I H,Zhang G,Hu H,etal.Assessment of the hazardous effects of Cd on physiological and biochemical characteristics of soybean genotypes[J].International Journal of Agriculture & Biology,2014,16(1):41-48.
[32] Zhang Z K,Li H,Zhang Y,etal.Grafting enhances copper tolerance of cucumber through regulating nutrient uptake and antioxidative system[J].Agricultural Sciences in China,2010,9(12):1758-1770.
[33] 孙胜,张智,卢敏敏,等.Cd2+胁迫对西瓜幼苗光合生理及活性氧代谢的影响[J].核农学报,2010,24(2):389-393.
[34] 李慧,丛郁,王宏伟,等.Cd对草莓幼苗根尖氧化系统和基因组DNA 损伤的影响[J].园艺学报,2010,37(5):721-730.
[35] Liang Y C,Chen Q,Liu Q.Exogenous silicon(Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (HordeumvulgareL.) [J].Journal of Plant Physiology,2003,160:1157-1164.
[36] 杨居荣,贺建群,黄翌,等.农作物Cd耐性的种内和种间差异[J].应用生态学报,1994,5(2):192-196.
[37] 李小红,陶建敏,陈剑东,等.Cd胁迫对不同砧穗组合葡萄植株Cd吸收规律和果实品质的影响[J].生态环境学报,2010,19(5):1082-1086.
猜你喜欢
中学生数理化·八年级物理人教版(2024年2期)2024-05-18 09:05:50
南方林业科学(2022年2期)2022-06-22 06:49:30
落叶果树(2021年6期)2021-02-12 01:29:12
现代农村科技(2019年3期)2019-03-20 09:19:52
新疆农垦科技(2018年3期)2018-04-24 08:17:28
现代园艺(2017年21期)2018-01-03 06:41:45
湖南农业(2017年1期)2017-03-20 14:04:46
工会信息(2016年4期)2016-04-16 02:39:06
微型小说选刊(2015年2期)2015-11-17 15:03:15
校园英语·中旬(2015年7期)2015-08-10 06:52:17
