
秦岭地区野生山丹遗传多样性表型及ISSR分析
2018-05-10 09:21丁芳兵孙伟博原雅玲寻路路
江苏农业科学 2018年7期
丁芳兵, 孙伟博, 原雅玲, 寻路路, 张 蕾
(1.陕西省西安植物园/陕西省植物研究所,陕西西安 710061; 2.南京林业大学,江苏南京 210037)
秦岭山区及周边地区百合资源丰富。近年来,有许多关于秦岭地区川百合、卷丹、野百合等百合群体形态学水平上遗传多样性的研究报道,这些种在群体内或群体间具有丰富的多态性,主要表现在生长、形态存在显著差异[1-5]。遗传多样性是指种内不同种群之间或同一种群不同个体间的遗传变异总和,除去个体水平上的形态发育差异,还包括细胞水平上的染色体差异及分子水平上的核酸、蛋白差异等。育种需要足够性状遗传多样性的种质资源,而分子生物学的发展为学者研究种质资源遗传多样性提供了更便捷、更精确的方法,目前,分析百合群体遗传多样性与亲缘关系的主要技术有即随机扩增多态性DNA(random amplified polymorphic DNA,简称RAPD)、内部简单序列重复(inter-simple sequence repeat,简称ISSR)、扩增片段长度多态性(amplified fragment length polymorphism,简称AFLP)等[6-8],其中ISSR标记重复性比RAPD好,操作简单、快速高效,且无需测序,是较适合百合研究的一项分子标记技术。
山丹(Liliumpumilum)作为药食两用百合,具有抗寒、抗旱、抗盐碱等多种优良性状,在秦岭地区广有分布,是百合杂交育种的优良亲本。目前,对秦岭地区山丹遗传多样性的研究非常有限,而利用表型性状和ISSR技术研究山丹不同居群的遗传多样性,不仅可以为分析山丹进化演变提供重要参考,而且可筛选出山丹不同居群的优良变异性状,为秦岭地区山丹资源的保护和利用提供依据。
1 材料与方法
1.1 材料
试验材料来自秦岭山周边的营盘镇、佛坪县、小峪、大峪口、太白县等地,共计9个居群,分别于2013、2014年5—9月在秦岭山区及周边地区采集共计30余次,每个居群采样个体数30个,个体间避免为同一无性系。各采样点详细信息见表1。

表1不同居群山丹采样点信息
1.2 试验方法
1.2.1 表型性状调查 采用游标卡尺测量山丹的花柱长度、花丝长度、茎粗、鳞茎最宽处直径,使用米尺测量株高。
1.2.2 基因组DNA的提取 采用天根生化科技(北京)有限公司生产的植物基因组DNA提取试剂盒,提取植株鳞茎最外部鳞片的DNA;提取的DNA通过1%琼脂糖凝胶电泳进行检测,DNA样品于-20 ℃冰箱内保存,备用。
1.2.3 PCR扩增和电泳 初筛选出25条引物,由上海Invitrogen(英骏)生物技术有限公司进行合成;每个居群随机选择2个模板进行扩增,最终选择P1:5′-CTCTCTCTCTCTCT TGA-3′、P2:5′-CTCTCTCTCTCTCTTAG-3′、P3:5′-CTCTC TCTCTCTCTTGT-3′这3条特征带清晰的引物进行PCR扩增。反应体系为20 μL:20~25 ng模板DNA 1 μL,Taq酶 0.5 μL,10 μmol/L ISSR引物 1 μL,2.5 mmol/L dNTP 2 μL,10×buffer 4 μL,加ddH2O至20 μL。 PCR反应条件为94 ℃预变性5 min;94 ℃变性60 s,55 ℃退火60 s,72 ℃延伸2 min,35个循环;72 ℃延伸10 min。扩增产物用聚丙烯酰胺凝胶进行电泳,染色后进行条带分析。
1.3 数据处理与统计
表型性状数据采用SPSS 19软件进行分析比对。ISSR结果在100~2 000 bp范围内选取重复性良好且清晰的条带进行记录统计,相同分子量片段对应1个多态性位点,有特异性条带的记为1,没有的记为0。
2 结果与分析
2.1 居群间表型性状变异分析
由表2可见,5个表型性状中,山丹居群SD3的植株相对最高、花柱相对最短、鳞茎最宽直径相对最大,居群SD1的植株相对最矮、鳞茎最宽直径相对最小,居群SD8的花丝相对最短,与其他居群差异显著(P<0.05);在茎粗指标上,9个居群差异不显著,表明其遗传稳定性相对较高;居群SD3、SD8的花柱长度、株高、鳞茎最宽直径与其他居群相比差异显著(P<0.05)。相关性分析结果表明,5个表型性状仅在某些居群中存在相关性,如居群SD1、SD2、SD3、SD5、SD7的株高与茎粗呈正相关,居群SD1、SD3、SD8的花柱、花丝长度与鳞茎最宽直径呈负相关,居群SD4、SD5的花柱、花丝长度与鳞茎最宽直径呈正相关,表明5个表型性状在9个居群间的相关性不高,居群特异性明显。方差分析结果表明,居群内各指标的差异不显著。

表2山丹9个居群的表型性状cm
注:同列数据后标有不同小写字母表示差异显著(P<0.05)。
2.2 表型性状与地理环境因子的相关性
由表3可见,仅花丝长度与生长环境呈负相关,其他4个表型性状与生长环境呈正相关;花柱、花丝长度与纬度及海拔呈负相关,而株高、茎粗、鳞茎最宽直径与海拔呈正相关,茎粗、鳞茎最宽直径与纬度呈负相关,株高与纬度呈微弱的正相关,大部分相关性未达显著性,仅花丝长度与经度呈显著正相关(P<0.05),表明地理因素对大部分表型性状无决定性影响。
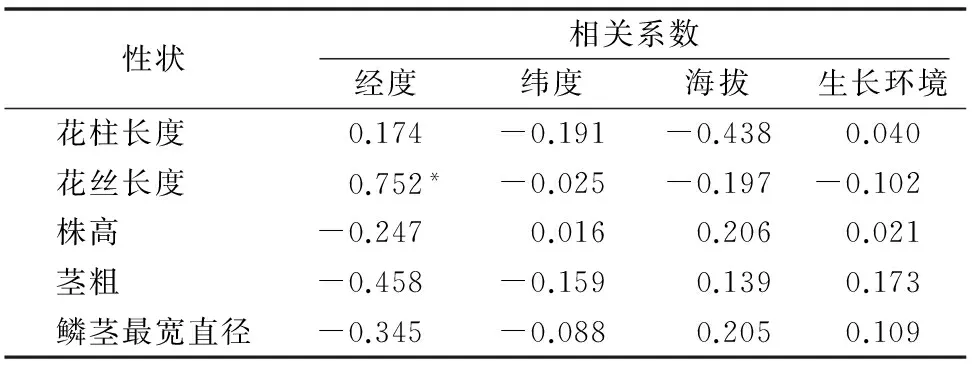
表3表型性状与地理环境相关性分析
注:“*”表示显著相关。
2.3 表型聚类分析
由图1可见,当遗传进化距离L=2时,野生山丹9个居群中除SD1、SD3、SD8为3个独立居群外,可以将其他居群分为1个大类,其中居群SD3、SD8生长环境不同,但海拔差异相对较小;在分为1大类的6个居群中,居群SD5与其他5个居群生长环境有明显差别,这可能是由于山丹在秦岭地区的迁徙经历海拔的变化,而迁徙线路环境的变化又造成山丹分布的生态多样性,同时,在迁徙过程中,也造成相对独立生态居群的产生。

2.4 ISSR多态性分析
由图2可见,从25条引物中筛选出的3条引物用于PCR扩增,条带数多且清晰。由表4可见,3条引物扩增片段的多态性比例平均值分别为44.1%、40.3%、49.0%,引物扩增多态性相对较低;不同引物扩增的多态性存在差异,说明山丹材料遗传背景具有一定的复杂性;所有样品共检测到522个位点,其中多态性位点235个,多态性比例为45.0%;3条引物居群内多态性位点比例在36.5%~55.2%之间,9个居群中SD1遗传多样性水平相对最高,SD4遗传多样性水平相对最低。

2.5 ISSR分子标记聚类分析
由图3可见,9个山丹居群根据ISSR标记结果可聚分为2个大类,居群SD1、SD3、SD6、SD8表现为相对独立的居群,居群SD6与SD4、SD5的亲缘关系相对更近,SD8与SD2、SD7、SD9的亲缘关系相对更近。结合地理因子分析发现,山丹多样性变化的主要原因是海拔的改变,其次是随经纬度的改变而不同,但表型性状与地理因子的相关性不显著。
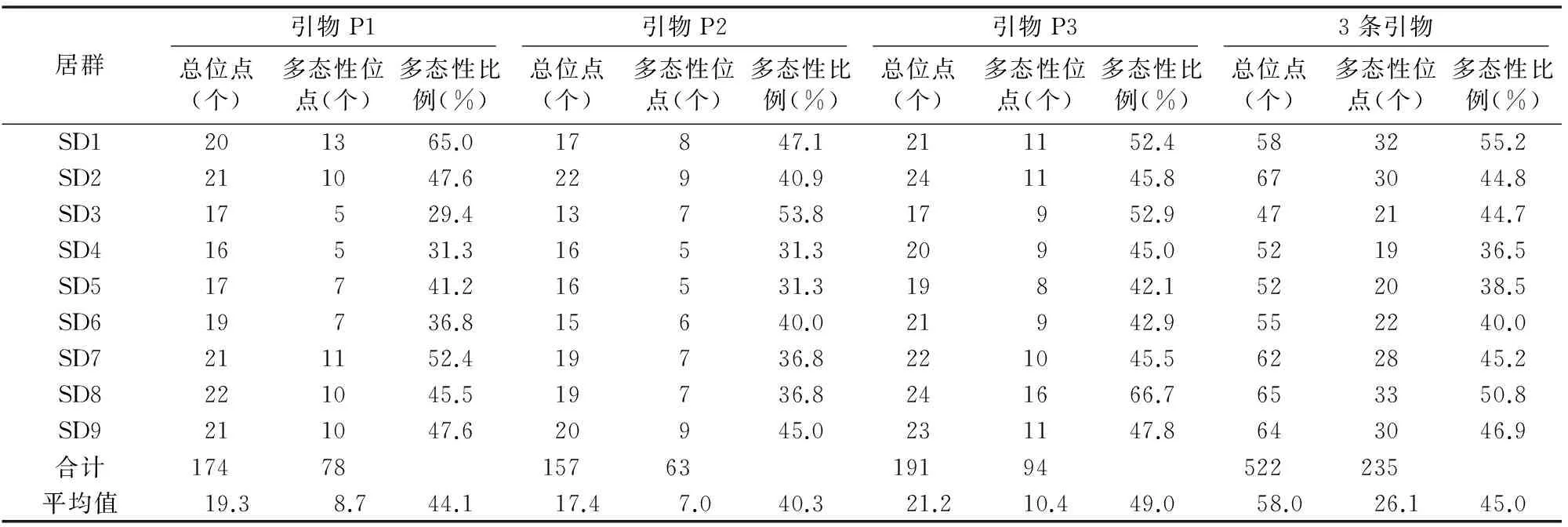
表43条引物的ISSR扩增结果

3 讨论
秦岭地区山丹居群大多数材料能够依据地理来源相对集中地聚在一类,如分别来自佛坪县(SD2)、户县东涝峪(SD6)的山丹居群海拔相近,分别来自长安区大峪(SD4)、太白县(SD5)、秦岭祥峪(SD7)、商洛县西柿沟(SD9)的4个居群纬度相近,且均分布在高海拔地区。同一类中居群有地理因子相差较大的,如SD4、SD9这2个居群的经度与SD5居群相差相对较远,这可能是起源相同的群体生长环境不同造成的。ISSR聚类分析结果表明,山丹居群聚类与地理分布的关系不很明显,这与葛新新等的研究结果[9]较为一致,这种相关性的不显著可能是由于采集地基本在秦岭附近,采样区域经纬度变化幅度有限,生态环境差异对遗传变异影响相对较小,有限的范围可能造成较多的基因交流,另外,对小生境的记录欠缺及利用鳞茎最宽直径进行ISSR标记可能技术不完善,都会造成这种结果。不过,山丹表型性状与地理因子有一定的相关性,特别是花丝长度与经度呈显著正相关(P<0.05),说明按经度分布可能是山丹的一条迁徙路线。ISSR聚类结果表明,居群SD7与SD9亲缘关系相对较近,这两者在地理环境因素上比较相似,结合地理因子可以推断,秦岭地区山丹变异的首要因素可能是海拔的变化。
秦岭地区山丹资源丰富,能为我国山丹的育种栽培、种质改良创新提供大量原始材料,对这些野生山丹资源进行调查、收集和保存,能为将来秦巴山区山丹百合资源的分类提供依据,对山丹的引种保护及驯化栽培起到指导作用。当前,有不少百合研究工作者对秦岭地区山丹资源进行了相关调查与收集,但对遗传等问题未做深入研究,有的仅单独运用传统形态学归类法或ISSR分子标记技术,缺少对2种方法的综合运用。本研究综合利用形态学归类法和ISSR分子标记这2种方法,能更完善地解释秦岭地区山丹资源的遗传特性,可为将来秦巴山区的百合资源调查研究及开发利用提供重要依据。
参考文献:
[1]赵祥云,陈新露,王树栋,等. 秦巴山区野生百合资源研究初报[J]. 西北农林科技大学学报(自然科学版),1990(4):80-84.
[2]向地英,张延龙,牛立新. 秦巴山区及毗邻地区野生百合的形态多样性研究[J]. 武汉植物学研究,2005,23(4):385-388.
[3]车 飞,牛立新,张延龙,等. 秦巴山区野生百合资源及其生境土壤特性的调查[J]. 安徽农业科学,2008,36(23):9955-9957.
[4]郭宇龙,张延龙,司国臣,等. 秦巴山区5种野生百合鳞片繁殖特性研究[J]. 西北林学院学报,2013,28(1):90-93,99.
[5]梁振旭,张延龙,牛立新,等. 秦巴山区野百合表型多样性[J]. 广西植物,2014(6):727-733.
[6]邓明文,吴祝华,席梦利,等. 岷江百合ISSR-PCR反应体系的优化[J]. 林业科技开发,2007,21(6):19-22.
[7]钟海丰,张延龙,牛立新. 秦巴山区野生山丹百合DNA提取与RAPD反应体系建立[J]. 西北农业学报,2008,17(3):285-289.
[8]丁信誉,邱 帅,席梦利,等. 东方百合ISSR-PCR反应体系的正交优化[J]. 南京林业大学学报(自然科学版),2012,36(5):42-46.
[9]葛新新,张文革,郭太君,等. 长白山区3种百合种内ISSR基因标记研究[J]. 吉林农业大学学报,2012,34(3):270-275.
猜你喜欢
甘肃畜牧兽医(2022年7期)2023-01-04
河北科技师范学院学报(2022年2期)2022-08-26
老年博览·上半月(2021年12期)2021-12-21
浙江中医药大学学报(2021年6期)2021-07-12
林业勘查设计(2020年1期)2021-01-18
中国粮油学报(2020年12期)2021-01-09
学生天地(2020年19期)2020-06-01
草地学报(2018年5期)2018-11-07
西南农业学报(2016年6期)2016-04-16
广西林业科学(2016年4期)2016-03-16
