
托里桉在我国南部的潜在地理适生区预测
2018-05-07 06:48欧阳林男陈少雄张维耀刘学锋何沙娥朱林生
桉树科技 2018年1期
欧阳林男,陈少雄,张维耀,刘学锋,何沙娥,朱林生
托里桉在我国南部的潜在地理适生区预测
欧阳林男,陈少雄*,张维耀,刘学锋,何沙娥,朱林生
(国家林业局桉树研究开发中心,广东 湛江 524022)
本文通过收集托里桉的实地栽培点地理数据,运用最大熵软件(Maxent)和地理信息系统技术(ArcGIS),结合12个气候因子数据,预测划定了托里桉在我国南部的潜在地理适生分布区。结果表明:Maxent模拟托里桉的潜在地理分布准确性较高,模型预测训练子集和验证子集AUC值均大于0.87。托里桉最适宜分布区集中在广东和海南沿海、广西沿海和中部及福建南部沿海,面积依次为广东(8.63 × 104km2)、海南(2.65 × 104km2)、广西(2.22 × 104km2)、福建(1.18 × 104km2);适宜区集中在广西中南部、广东中北部、福建南部、江西中南部,面积依次为广西(10.17 × 104km2)、广东(6.15 × 104km2)、福建(2.52 × 104km2)、江西(1.43 × 104km2)。Maxent刀切法(Jackknife)分析结果表明:影响托里桉适生区分布的主导气候因子为年平均气温、≥10℃积温、极端低温、最冷月最低温、最冷月均温、≥10℃的天数。
托里桉;适生区分布;主导气候因子;最大熵(Maxent)模型
托里桉()为常绿乔木,树皮绿色光滑,树冠密集,幼枝嫩叶有红色绒毛,枝叶婆娑[1];其叶有疏风解热、消炎抗菌和杀虫作用[2]。一些低纬度国家已成功引种了托里桉,如托里桉在斯里兰卡主要用做用材林树种[3];在尼日利亚,托里桉叶片浸出物用于治疗消化溃疡疾病和其他肠胃疾病[4];在刚果,托里桉用于森林建设、抗土壤侵蚀,其叶片用于产油[5];在美国佛罗里达州,将其用做能源植物,用于开发林化产品、生物燃料等能源产品[6];在印度南部,托里桉用做纸浆材树种。我国云南最早引种托里桉,随后1974年引入广西东门林场,1979年引入广东,华南农业大学开始育苗造林,至今在南亚热带以南地区用作四旁绿化、庭院布置、低丘造林优良树种[1]。
国内外有关托里桉的研究主要集中在植物叶片提取物抗菌能力[7]、叶片香精油化学成分[8]、根系菌根诱导[9]、生根能力和扦插活力测试[10]、木材形稳性改良[11]、微繁殖细胞分裂素对植物生长和根部结构的影响[12]、体外组织培养和微繁殖技术及其对生物量增产的作用[13-14]、杂交种遗传优势[15]、干旱胁迫下的植物生理响应[16]等,而对托里桉在我国的潜在地理适生区分布暂无报道。本文利用MaxEnt生态软件,结合托里桉在我国的已知栽培点和气象数据划分其在我国南方十省(区)的潜在适生区,为托里桉在我国的培育和推广提供科学依据。
1 材料与方法
1.1 模型构建
1.1.1 数据收集
检索国内外与托里桉有关的文献报道,收集托里桉在我国南方的引种地点共28个。28个样点中,大部分经纬度信息准确,可直接使用;少数样点仅有地名,利用Google earth获取经纬度。
1.1.2 环境因子
1981—2010年的广东、四川、江西、重庆、湖南、云南、广西、贵州、海南、福建十个省(区)中各县气象站点的现实地面气候标准值数据集由我国气象数据网提供,共记768个气象站(图1),气象数据包括5类指标:
反映平均温度的指标:年平均气温、最冷月平均气温、最热月平均气温。
反映极端温度的指标:最冷月平均最低温、最热月平均最高温、极端低温。
反映水分条件的指标:年降水、年均相对湿度。
反映热量供应的指标:≥10℃积温、≥10℃的天数、太阳辐射。
反映海拔高度的指标:高程。
图1 我国南方十省(区)气象观测站分布
1.2 模型预测
1.2.1 模型精度
首先,将托里桉地理分布数据和提取的环境因子变量数据导入MaxEnt模型(3.4.1)中,由于托里桉实际引种地点较少,为保证模型预测的准确性,随机选取引种点数据的90%作为训练子集(training data)用以训练模型,剩余10%的数据作为评估子集(testing data)用以验证模型。其次,通过模型中的AUC评价指标对模型效果进行检测,验证模型准确度。AUC值是模型ROC曲线与横坐标围成的面积(the area under the ROC curve),不同值代表不同预测精度:AUC 0.5 ~ 0.6,预测结果不可接受;AUC 0.6 ~ 0.7,预测结果较差;AUC 0.7 ~ 0.8,预测结果一般;AUC 0.8 ~ 0.9,预测结果良好;AUC 0.9 ~ 1.0,预测结果优秀。
1.2.2 适生性分析
以上述分析为基础,将MaxEnt模型输出结果导入ArcGIS 10.2软件进行适生性分析。进行重分类操作,依照适生性指数将托里桉分布区划分成4个等级:不适宜区(<0.05)、次适宜区(0.05≤<0.33)、适宜区(0.33≤<0.66)、最适宜区(≥0.66)。
1.2.3 适生区面积计算
使用ArcGIS 10.2中的Raster Reclassify功能,分别计算出各省(区)4个等级适宜区面积。
1.2.4 关键环境因子筛选
使用MaxEnt中的刀切法(Jackknife)模块对气候因子的重要度进行计算,得出各环境变量对模型预测的贡献值,评价各环境因子权重,筛选出主导托里桉潜在地理适生区分布的关键环境因子。
2 结果与分析
2.1 模型模拟结果的准确性
基于MaxEnt软件模拟托里桉在我国南方十省(区)的潜在分布,以ROC曲线为模型预测准确性衡量指标(图2)。结果表明,训练子集和验证子集AUC值均大于0.87(表1),说明模型预测准确性较好,可用于托里桉适生区分布与气候因子关系的研究。

表1 模型AUC预测精度
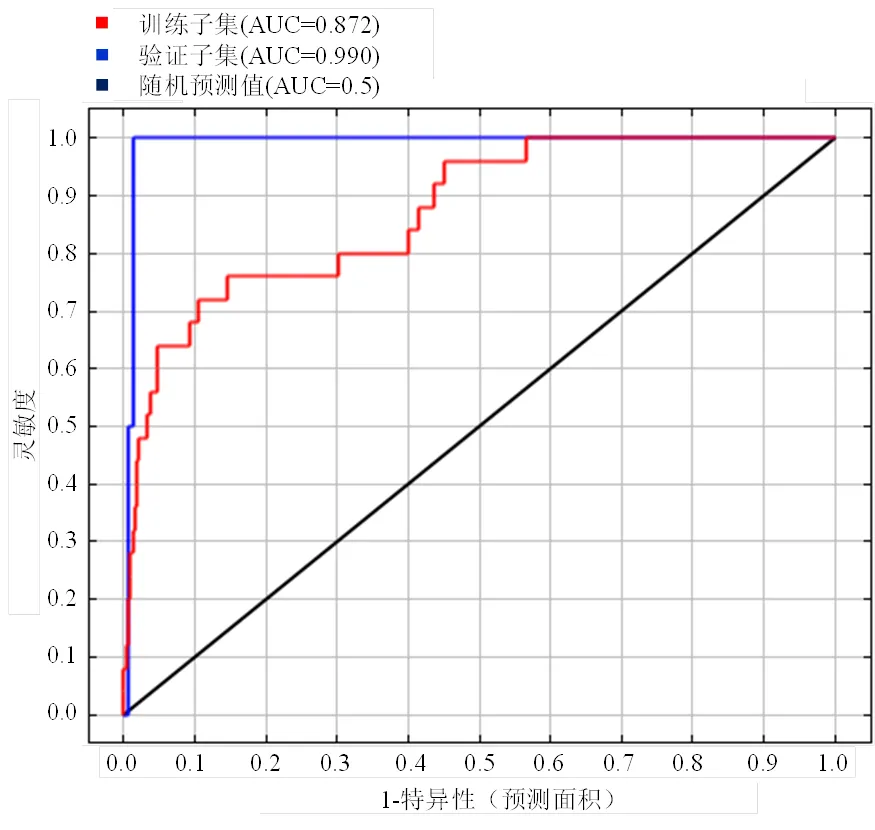
图2 托里桉分布预测的ROC曲线
2.2 托里桉在我国南部的潜在适生区
表2为托里桉在我国南方十省(区)的适生区统计适生面积。使用ArcGIS计算,托里桉在我国南方十省(区)的最适宜区总面积14.77 × 104km2,适宜区总面积22.23 × 104km2,次适宜区总面积105.06 × 104km2,与油松()最适宜区总面积2.6 × 104km2[17]在我国的潜在地理分布相比,托里桉在我国的发展潜力更大。托里桉适生区主要集中在南部沿海地区,其中最适宜区面积最大的为广东省(8.63 × 104km2),最适宜区占各省区面积比例最大的为海南省(82.05%),适宜区面积最大的为广西(10.17 × 104km2)。
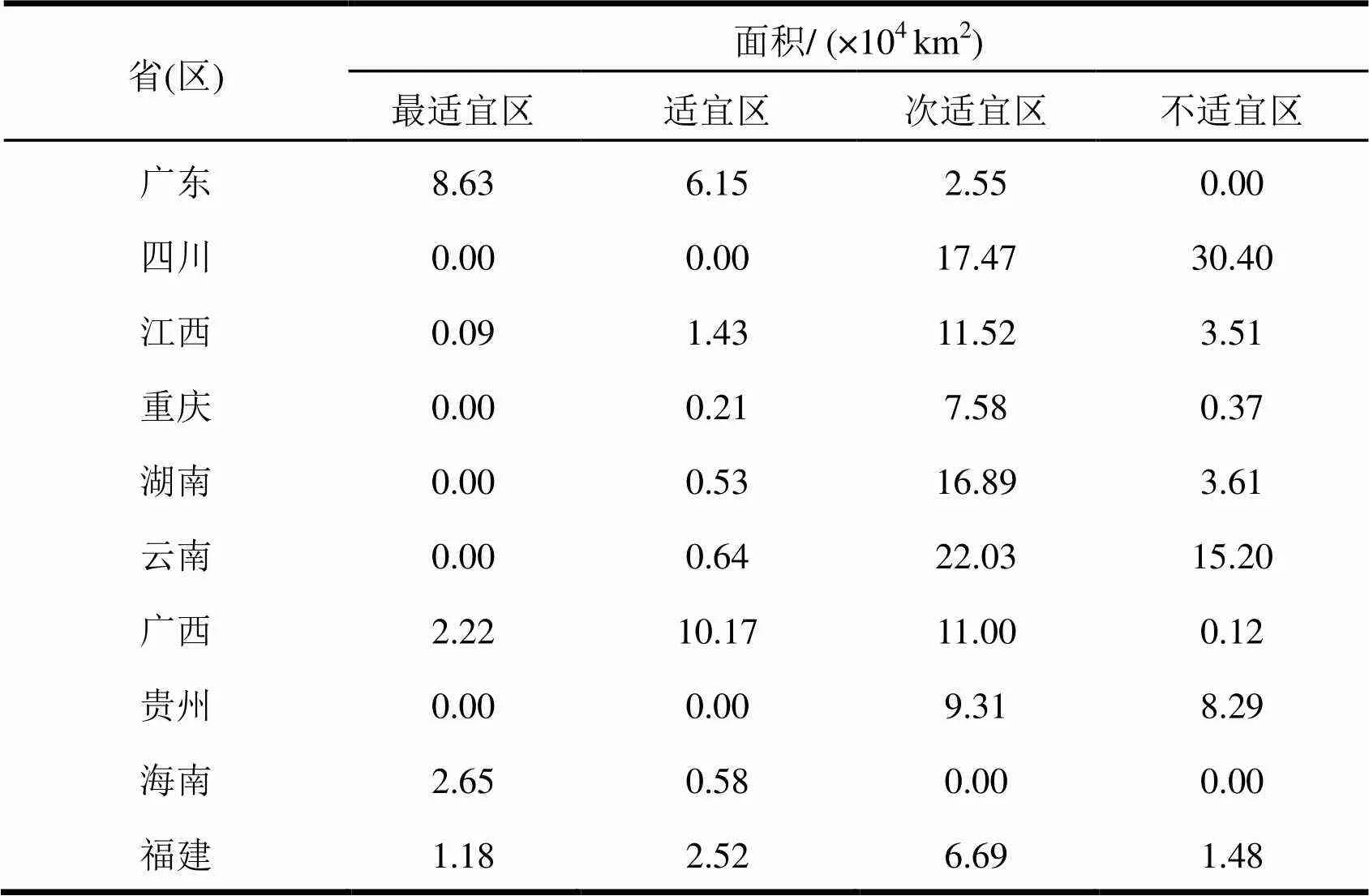
表2 托里桉在我国南方十省(区)的适生区分布面积
图3和图4展示了托里桉在我国南方十省(区)的适生区分布。最适宜分布区集中在广东和海南沿海、广西沿海和中部,福建南部沿海;适宜区集中在广西中南部、广东中北部、福建南部、江西中南部。海南省和广东省的托里桉最适宜区面积分别是其省总面积的82.05%和49.79%。
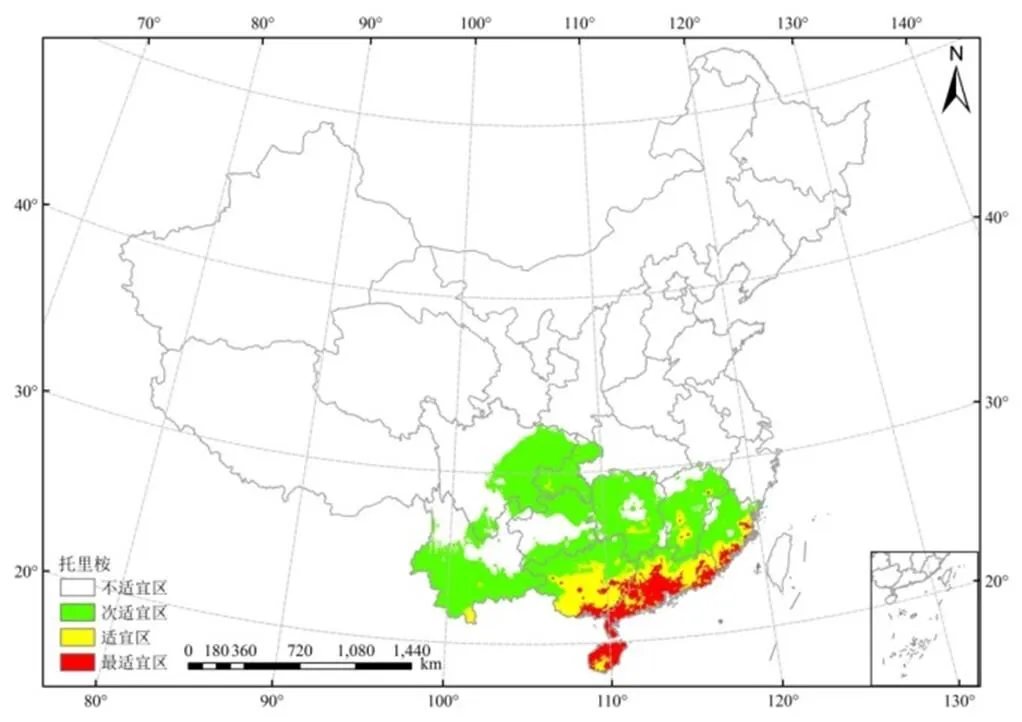
图3 基于Maxent模型的托里桉潜在适生区
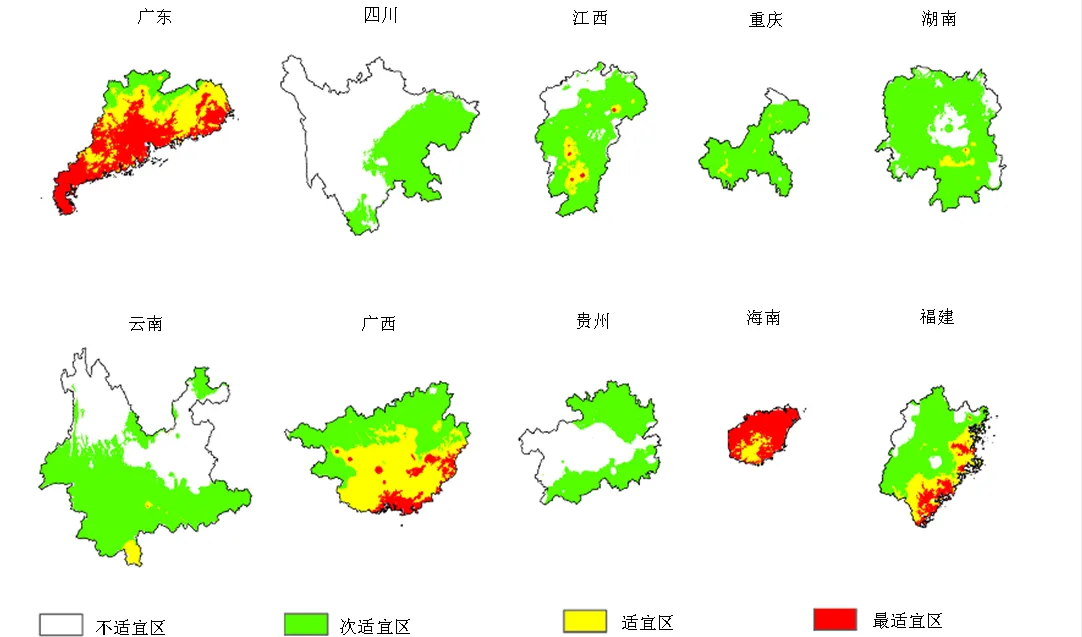
图4 托里桉在我国南方十省(区)的适生区分布
2.3 影响托里桉分布的不同气候因子评价
图5显示了各气候因子对托里桉适生区分布的贡献情况。影响托里桉适生区分布的环境气候因子按其贡献度由高到低排名依次为年平均气温、≥10℃积温、极端低温、最冷月最低温、最冷月均温、≥10℃的天数、高程、年降水、最热月均温、太阳辐射、最热月平均最高温、年平均相对湿度。综合来看,年平均气温、≥10℃积温是影响托里桉分布的最主要气候因子,其次为极端低温、最冷月最低温、最冷月均温,≥10℃的天数、高程对托里桉分布也有一定影响,但其影响程度比年平均气温、≥10℃积温小。
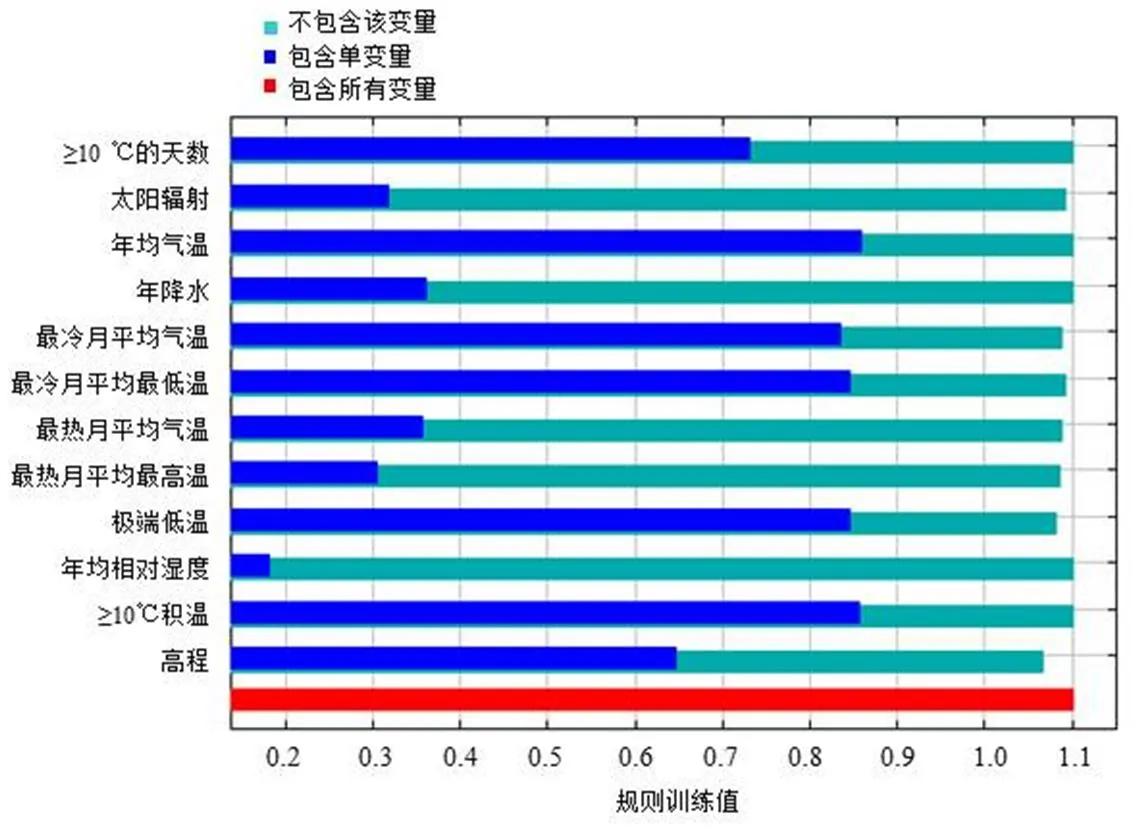
图5 基于Jackknife的气候因子对托里桉在我国南方十省(区)分布的贡献
3 讨论
本文利用MaxEnt生态软件与地理信息系统相结合的方法预测了原产于澳洲的托里桉潜在我国南部的分布,获得了托里桉在我国南方十省(区)的潜在分布图。由于托里桉在国内成功引种的范例较少,本文只收集到28个引种栽培地点。经检验,MaxEnt模型的受试者工作特征曲线AUC值均接近于1,预测结果较好。
桉树品种的地理分布主要取决于气候差异[3]。本研究发现,托里桉在我国南部十省(区)的适生地主要集中在沿海地区,最适宜分布区集中在广东和海南沿海、广西沿海和中部,福建南部沿海,这与南部沿海地区温暖湿润的气候特征相关。广东和海南为托里桉最适宜区分布面积最大的省份,这主要与广东和海南的年均气温和年降水排列前二相关。托里桉为四季常绿植物种,其蒸腾速率快,较其他品种需要更强的光照和水分条件,因而为维持其正常的新陈代谢,需要在气温和年降水均较大的地区生长[18]。
本研究结果表明,托里桉适生区主要集中在南部沿海地区,其中最适宜区面积最大的为广东省(8.63 × 104km2),最适宜区占各省区面积比例最大的为海南省(82.05%),适宜区面积最大的为广西(10.17 × 104km2),这同样与广东、海南、广西的温暖湿润气候条件相关。据报道,阴冷、低温的天气对托里桉花蕾发育非常不利,严重时受冻害死亡,结果极少[19],因而常年温暖、年间低温天气较少的地区有利于托里桉花蕾顺利孕育、来年结实,从而促进托里桉人工林生长,有助于增强其形成的森林生态系统稳定性。托里桉适宜区面积以广西、广东的为最大,适宜区在内陆的分布以福建南部和江西中南部面积最大,这与托里桉早期成功引种于福建的结果相一致[1],为托里桉在我国内陆的栽植提供了方向。学者还通过叶片解剖结构分析证明托里桉具有一定抗旱、抗寒能力,观察到福建引种托里桉的叶片并不全为等面叶,与原产地澳大利亚桉树叶片结构有所不同,推测从原产地引种至我国可能会通过改变叶片结构表达从而导致其适应性的变化[20]。
本研究发现,在影响托里桉适生区分布的12个环境因子中,年平均气温是最主要的影响因子,其次为≥10℃积温、极端低温、最冷月最低温、最冷月均温,表明平均温度、热量供应是影响托里桉适生区分布的关键气候因子类型。有研究指出,托里桉的生长及蒸腾速率主要受环境因子中的温度与光照所影响,这主要与托里桉四季常青、无明显换叶季节的生理生长特性相关,其高蒸腾量需要更充足的温度与光照,因而对温度与光照气候因子更为敏感[18]。在温度与光照强度出现高峰值的夏季,托里桉的蒸腾失水量为最大,在此时期保持土壤适度含水量成为维持植体水分动态平衡,进而提高托里桉人工林生长状况的关键。除气候因子外,其他非生物因素对托里桉的生长发育和分布也很重要,如场地土壤气孔的控制水分流失能力在干旱季节水分胁迫时期能保证植物的正常生长和发育[16],树木在空气中二氧化碳浓度增大时可通过减小叶片气孔导度和降低蒸腾作用强度增大自身的光合作用和对土壤水分的利用效率,提高生长适应性[21]。
4 结论
在我国南部的十省(区)中,托里桉在广东、海南、广西、福建、江西的适生区面积较大,在十省(区)最适宜区面积共14.77 × 104km2,适宜区总面积22.23 × 104km2,其在我国的发展潜力较油松等木本植物大。托里桉的适生区集中在南部沿海地带,最适宜分布区集中在广东和海南沿海、广西沿海和中部,福建南部沿海;适宜区集中在广西中南部、广东中北部、福建南部、江西中南部。
在影响树种适生区分布的12个环境因子中,年平均气温、≥10℃积温、极端低温、最冷月最低温、最冷月均温对托里桉的生长分布影响最大。
MaxEnt最大熵模型对托里桉预测的ROC受试者工作特征曲线AUC值均接近于1,说明模型对托里桉适生区预测的结果较好。在我国南方引种推广托里桉,应首先考虑在本研究结果中的最适宜区和适宜区进行引种栽培,而在次适宜区和不适宜区栽培需谨慎考虑。
[1] 窦志浩,温茂元,吴光儒,等.托里桉引种试验初报[J].热带作物研究,1989(4):60‒65.
[2] Adeniyiand B A, Ayepola O O.The phytochemical screening and antimicrobial activity of leaf extracts ofand(Myrtaceae)[J].Research Journal of Medicinal Plant, 2008, 2(1):34‒38.
[3] Sivananthawerl T, Mitlohner R, Günter S, et al.and other importantspecies:a case study from Sri Lanka[J].Tropical Forestry,2011,8: 463‒472.
[4] Lawal T O, Adeniyi B A, Moody J O, et al. Combination studies ofF.Muell. leaf extracts and clarithromycin on Helicobacter pylori[J].Phytotherapy Research,2012,26(9):1393‒1398.
[5] Silou T, Loumouamou A N, Makany A R, et al. Multivariate statisdcal analysis of the variability of essential oils from the leaves ofacclimatised in Congo‒Brazzaville[J].Journal of Essential Oil Bearing Plants,2010,13(4): 503‒514.
[6] Rockwood D L, Rudie A W, Ralph S A, et al.Energy product options forspecies grown as short rotation woody crops[J].International Journal of Molecular Sciences,2008, 9(8):1361‒1378.
[7] Silou T, Loumouamou A N, Loukakou E, et al.Intra and interspecific variations of yield and chemical composition of essential oils from fivespecies growing in the Congo‒Brazzaville. Corymbia Subgenus[J].Journal of Essential Oil Research, 2009,21(3):203‒211.
[8] 陈婷婷,周晓农,朱丹,等.托里桉叶挥发油化学成分的气相色谱-质谱联用分析[J].今日药学,2011,21(10):620‒623.
[9] Chan W K, Griftiths D A. The induction of mycorrhiza inandgrown in Hong Kong[J].Forest Ecology and Management,1991, 43(1/2): 15‒24.
[10] Wendling I, Brooks P R, Trueman S J. Topophysis in×seedlings: adventitious rooting capacity, stem anatomy, and auxin and abscisic acid concentrations[J]. New Forests, 2015, 46(1): 107‒120.
[11] Paes J B, Loiola P L, Oliveira J T D S, et al. Effect of salt solutions in dimensional stability ofandwoods[J].Scientia Forestalis/ forest Sciences, 2015, 43(106): 271‒281.
[12] Hung C D, Trueman S J. Cytokinin concentrations for optimal micropropagation of×[J].Australian Forestry,2012,75(4): 233‒237.
[13] Khuspe S S, Gupta P K, Kulkarni D K, et al. Increased biomass production by tissue culture of[J].Canadian Journal of Forest Research, 1987, 17(11): 1361‒1363.
[14] Arya I D, Sharma S, Arya S. Micropropagation of superior eucalyptus hybrids FRI‒5 (Dehn ×Sm) and FRI‒14(F.V. Muell ×Hook): A commercial multiplication and field evaluation[J].African Journal of Biotechnology,2009, 8(21):5718‒5726.
[15] Paramathma M, Surendran C, Rai R S V. Studies on heterosis in sixspecies[J].Journal of Tropical Forest Science, 1997, 9(3): 283‒293.
[16] Paula L W D, Paul J, Sophia R, et al. Stomatal responses ofspecies to elevated CO2concentration and drought stress[J].Scientia Agricola,2003,60(2):231‒238.
[17] Li G Q, Xu G H, Guo K, et al. Geographical boundary and climatic analysis ofin China: Insights on its afforestation[J].Ecological Engineering, 2016,86: 75‒84.
[18] 谭绍满.托里桉蒸腾量的研究[J].桉树科技,1982(3):19‒25.
[19] 谭绍满.托里桉[J].热带林业科技,1986(2):50‒52.
[20] 安平,洪长福,蔡振坤.福建引种的13种桉树叶片解剖分析[J].桉树科技,1992(1):57‒61.
[21] Medlyn B E, Barton C V M, Broadmeadow M S J, et al. Stomatal conductance of forest species after long‐term exposure to elevated CO2concentration:a synthesis[J]. New phytologist, 2001,149(2): 247‒264.
Predicting Potential Geographic Distribution ofin Southern China
OUYANG Lin-nan, CHEN Shao-xiong, ZHANG Wei-yao, LIU Xue-feng, HE Sha-e, ZHU Lin-sheng
()
The areas climatically suitable forplantations in southern China were analyzed using maximum entropy modeling (Maxent) and ArcGIS spatial analysis technology in this study, by combining data on the current distribution of.and related climatic factors in China. The results showed that the Maxent model is able to predict the potential distribution ofwith a high accuracy – the correspondence between the training data and test data was over 0.87. The majority of area highest suitable for this species is concentrated in the coastal regions of Guangdong (8.63 × 104km2), Hainan (2.65 × 104km2), the coastal and central regions of Guangxi (2.22 × 104km2) and the southern coastal regions Fujian (1.18 × 104km2). The area highly suitable for the species is concentrated in the central southern regions of Guangxi (10.17 × 104km2), central northern regions of Guangdong (6.15 × 104km2), southern areas of Fujian (2.52 × 104km2) and central southern regions of Jiangxi (1.43 × 104km2). Jackknife analysis of Maxent showed that the dominant climatic factors influencing the potential geographic distribution ofare annual mean temperatures, accumulated temperature of 10oC or more, extreme lowest temperature, lowest temperature of the coldest month, average temperature of the coldest month and the number of days with temperature above 10oC.
; potential geographical distribution; dominant climatic factor; maximum entropy (MaxEnt) model
S724
A
国家重点研发计划课题(2016YFD0600502);广东省林业科技创新项目(2016KJCX005)。
欧阳林男(1990— ),女,博士,助理研究员,主要从事桉树人工林培育研究,E-mail:1332356525@qq.com.
陈少雄(1965— ),男,博士,研究员,主要从事桉树定向培育研究,E-mail:sxchen01@163.com.
猜你喜欢
中国中医药信息杂志(2022年7期)2022-07-17
中国野生植物资源(2022年3期)2022-04-19
云南农业大学学报(自然科学)(2022年2期)2022-04-07
福建农林大学学报(自然科学版)(2022年1期)2022-01-18
——浅析美国短篇小说《宰恩的召唤》中的爱情观
长江丛刊(2020年34期)2020-11-18
孩子(2020年8期)2020-08-13
孩子(2020年12期)2020-01-06
南方农业·下旬(2016年9期)2017-04-27
江苏农业科学(2017年5期)2017-04-15
科技与创新(2016年18期)2016-11-04
