
大鼠血清外泌体对MCF-7细胞自噬的影响
2018-05-02 03:46毛建文
中西医结合心血管病杂志(电子版) 2018年7期
陈 奇,徐 彬,毛建文*
(1.广东药科大学基础学院,广东 广州 510006;2.广东药科大学生命科学与生物制药学院,广东 广州 510006)
1963年,比利时的细胞学家克里斯汀德迪夫率先提出了一个术语“自噬”,即一种由细胞自身调控的毁灭机制,用来降解无用或功能失调的组份[1]。大隅良典于90年代阐明了细胞自噬的机制,研究者们直至此时才发现自噬在人体中的重要作用[2]。外泌体是一种细胞来源的囊泡,存在于许多乃至所有真核生物的体液中,包括血液、尿液以及细胞培养的培养液中[3-4]。几乎所有类型的细胞都通过自噬克服饥饿、回收养分以及将多余的或有害的成分转运至细胞外。同时外泌体能够在细胞间传递,因此在某些情况下这一转运过程是通过外泌体进行的[5]。目前关于血清外泌体的研究大多集中于作为新的生物标志物用于活体检验方面,对于血清外泌体与肿瘤及自噬之间的研究较少。本研究观察了大鼠正常血清外泌体对乳腺癌(MCF-7)细胞自噬的影响。
1 材料与方法
1.1 药物、试剂及仪器
人乳腺癌细胞株MCF-7;高糖DMEM培养基;四季青胎牛血清;胰岛素(Sigma公司);胰酶;雷帕霉素(Rapamycin);CCK-8试剂盒、LC3B抗体、GAPDH抗体、鼠抗、兔抗(碧云天公司);脱脂奶粉
1.2 细胞培养及血清外泌体提取及鉴定
MCF-7细胞用DMEM培养液(含10 %胎牛血清、0.01 mg/ml胰岛素)培养。培养条件为37℃、5%CO2。取1 ml正常SD大鼠血清,试剂盒法提取,用PBS重悬至200 μl,加入DMEM(含10%FBS)培养液,配置成每100 μl培养液含10 μl外泌体,0.22 μm微孔滤膜过滤。对所提取的血清外泌体采用透射电镜检测其形态特征,采用WB法鉴定特征性蛋白质。
1.3 血清外泌体对MCF-7细胞存活率的影响观察
取对数生长期的MCF-7细胞,PBS洗3次,用0.25%的胰酶消化,按4×104个/孔接种于96孔细胞培养板,分为Control、雷帕霉素(Rapa)、外泌体(Exo)、Exo+Rapa组,每组设3个平行复孔。Control组不加任何药物培养;Rapa组加入30nM Rapa,作用24 h;Exo组加入过滤后的含10%外泌体的培养液,培养48 h;Exo+Rapa组先加入含10%外泌体的培养液,培养48 h,然后加入30 nM Rapa,作用24 h。处理结束后使用CCK-8试剂盒,孵育1 h,检测450 nm波长处的吸光度(OD)。取3孔OD值的平均值。
1.4 MCF-7细胞LC3B的荧光检测
细胞分组同1.3。吸弃培养基,PBS洗3次,固定15 min。去除固定液,洗涤后加入封闭液,37℃下封闭1 h。加入LC3B抗体(1:100)4℃孵育过夜,移除抗体洗涤后加入绿色DyLight 488荧光二抗,37℃孵育1 h,洗涤后加入DAPI染核,加入抗荧光淬灭剂,封片。用荧光显微镜观察,在相同曝光时间下拍照。
1.5 MCF-7细胞LC3B蛋白检测
细胞分组同1.3。采用Western blotting法,提取MCF-7细胞总蛋白,用BCA法检测蛋白浓度,SDS-PAGE电泳后转膜2 h,5%奶粉室温封闭2 h,TBS/T洗涤后加入一抗(LC3B,1:1000;GAPDH,1:1000)4℃孵育过夜。洗涤后加入相应二抗,37℃孵育1 h。使用ECL进行显影,用Image J分析软件定量分析目的条带的灰度值。
1.6 统计学方法
2 结 果
2.1 血清外泌体的分离鉴定
血清外泌体在透射电镜下呈茶托样、双分子层结构。Western blotting法结果显示,试剂盒提取的血清外泌体具有标志性的CD63及HSP70蛋白。
2.2 MCF-7细胞存活率的比较
Control、Rapa、Exo及Exo+Rapa组的OD值分别为0.367±0.043、0.345±0.031、0.650±0.051、0.420±0.020。结果显示,雷帕霉素对MCF-7细胞的生长有一定影响,但抑制作用并不明显,血清外泌体则明显促进MCF-7细胞的增殖。
2.3 MCF-7细胞LC3B表达的荧光结果比较
如图1所示,颗粒状绿色荧光为自噬标志性蛋白LC3B。Control组中,MCF-7细胞有少量的自噬现象;Rapa组中,由于加入自噬诱导剂,细胞的自噬显著增多;而Exo组的细胞自噬现象与正常对照组相比更少,几乎无自噬现象;至于Exo+Rapa组的细胞则与Control组相类似。
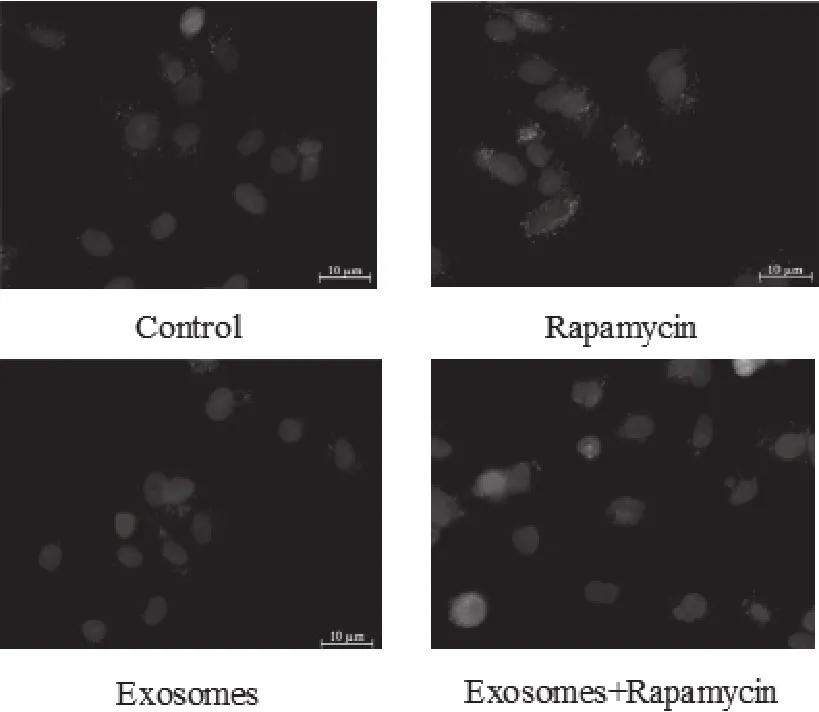
图1 MCF-7细胞LC3B表达的荧光结果比较
2.4 MCF-7细胞中LC3B的相对表达量比较
LC3B 2与LC3B 1比值的大小通常用于估计自噬水平的高低,比值越大则自噬水平越高。Control、Rapamycin、Exo、Exo+Rapa组LC3B2/LC3B1的比值分别为1.317±0.430、2.308±1.085、0.808±0.350、1.110±0.496。结果显示,Exo组与Rapa组的差异具有统计学意义(P<0.05)。Exo+Rapa组与Rapa组的差异有统计学意义(P<0.05)。外泌体能够降低MCF-7细胞的自噬且能抑制雷帕霉素诱导的自噬。见表2。

图2 MCF-7细胞LC3B的相对表达量比较
3 讨 论
乳腺癌时至今日仍是发展中国家女性癌症死亡的主要原因,在发达国家发病率亦居高不下[6]。研究发现,外泌体是一种重要的信号分子,参与细胞通讯,在免疫应答和肿瘤转移中起重要作用[7]。现阶段的研究大多集中于肿瘤分泌的外泌体对癌症的发展[8]、转移[9,10]、化学耐药[11]等多方面的影响。对于血清外泌体的研究则多为生物标志物[12],其对肿瘤会产生何种影响仍未可知。
我们的研究结果发现,血清外泌体能够抑制乳腺癌细胞的自噬。而自噬与癌症紧密相关,并在肿瘤中起双重作用,能够抑制肿瘤发生但对肿瘤生长又有着促进作用[13-16]。那么血清外泌体是否能通过抑制肿瘤细胞的自噬,从而抑制肿瘤的生长呢,其抑制自噬的机制又是什么,以及血清外泌体是怎样从血液进入到肿瘤细胞中的?这些问题值得进一步深入的探讨。总而言之,本研究的发现,为抑制肿瘤生长提供了一个新的靶点,为血清外泌体的功能研究开拓了新的方向。
[1]D.J.Klionsky,Autophagy revisited:a conversation with Christian de Duve[J]. Autophagy.2008,4(6):740-3.
[2]J. M.M.Levy,C. G.Towers, A. Thorburn, Targeting autophagy in cancer[J]. Nat Rev Cancer.2017,17(9):528-542.
[3]S.Keller,M.P.Sanderson, A. Stoeck,et al.Exosomes: from biogenesis and secretion to biological function[J].Immunol Lett.2006,107(2):102-8.
[4]E.van der Pol,A.N.Boing,P.Harrison,et al.,Classification, functions,and clinical relevance of extracellular vesicles[J].Pharmacol Rev.2012,64(3):676-705.
[5]F. Baixauli,C.Lopez-Otin,M.Mittelbrunn, Exosomes and autophagy: coordinated mechanisms for the maintenance of cellular fitness[J]. Front Immunol. 2014, 5(403.
[6]L. A.Torre,F.Bray,R. L.Siegel,et al.,Global cancer statistics,2012[J]. CA Cancer J Clin.2015,65(2):87-108.
[7]L. Murrow,R.Malhotra,J.Debnath,ATG12-ATG3 interacts with Alix to promote basal autophagic flux and late endosome function[J]. Nat Cell Biol.2015,17(3):300-10.
[8]Y.L.Hsu,J.Y.Hung,W.A.Chang, et al., Hypoxic Lung-Cancer-Derived Extracellular Vesicle MicroRNA-103a Increases the Oncogenic Effects of Macrophages by Targeting PTEN[J].Mol Ther.2018,26(2):568-581.
[9]W. Zhou,M.Y.Fong,Y.Min, et al.,Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis[J].Cancer Cell. 2014, 25(4): 501-15.
[10]A. Hoshino, B. Costa-Silva,T.L.Shen, et al., Tumour exosome integrins determine organotropic metastasis[J].Nature.2015,527(7578):329-35.
[11]B.Y.Nabet,Y.Qiu,J. E.Shabason, et al., Exosome RNA Unshielding Couples Stromal Activation to Pattern Recognition Receptor Signaling in Cancer[J].Cell.2017,170(2):352-366.
[12]K. G.Ronquist, G.Ronquist.A.Larsson, et al., Proteomic analysis of prostate cancer metastasis-derived prostasomes[J].Anticancer Res.2010,30(2): 285-90.
[13]E. White,Deconvoluting the context-dependent role for autophagy in cancer[J]. Nat Rev Cancer.2012,12(6):401-10.
[14]A. F.Fernandez, C.Lopez-Otin,The functional and pathologic relevance of autophagy proteases[J]. J Clin Invest.2015,125(1):33-41.
[15]D. Herranz,A. Ambesi-Impiombato,J. Sudderth, et al., Metabolic reprogramming induces resistance to anti-NOTCH1 therapies in T cell acute lymphoblastic leukemia[J]. Nat Med.2015,21(10):1182-9.
[16]C. M.Sousa, D. E. Biancur,X.Wang, et al.Pancreatic stellate cells support tumour metabolism through autophagic alanine secretion[J].Nature.2016,536(7617):479-83.
猜你喜欢
——雷帕霉素
食品与健康(2022年8期)2022-10-22
昆明医科大学学报(2022年2期)2022-03-29
国际眼科杂志(2022年3期)2022-03-11
中学生物学(2021年8期)2021-11-02
昆明医科大学学报(2021年5期)2021-07-22
天津医科大学学报(2021年3期)2021-07-21
实用药物与临床(2021年2期)2021-03-18
老年医学研究(2021年6期)2021-03-09
中老年保健(2020年2期)2020-12-04
山西农业大学学报(自然科学版)(2020年1期)2020-03-04
