
油橄榄花芽分化期内源激素及碳水化合物含量的变化研究*
2018-04-25 07:26杜晋城李丕军辜云杰王泽亮邢文曦郑崇文
西部林业科学 2018年1期
杜晋城,李丕军,辜云杰,王泽亮,邢文曦,郑崇文
(四川省林业科学院,四川 成都 610081)
花芽分化是指植物的芽轴生长点经过生理和形态的变化,最终构成各种花器官原基的过程[1],植物内源激素水平和养分状况对花芽分化有着重要的影响。因此,在植物花芽分化过程中,内源激素调节是花芽分化的关键,养分是花芽分化的基础,基因表达是花芽分化的途径[2]。
油橄榄(OleaeuropaeaL.)是木犀科(Oleaceae)木犀榄属油料树种,原产于地中海沿岸地区,有4 000多年的栽培历史,是天然优质的食用植物油,被誉为“植物油皇后”、“液体黄金”[3]。我国自20世纪引种栽培以来,生产中普遍存在着不完全花比例大,座果率低的问题,国内也未对油橄榄花芽分化期的生理机制进行过相关研究,因此,本文以生产中完全花比例大、座果率高的‘阿贝基娜’(O.europaea‘Arbequina’),完全花比例小、座果率低的‘卡琳’(Oleaeuropaea‘Kalinjoti’)2个品种为材料,在对花芽分化过程形态变化进行观察的基础上,分析花芽分化期的内源激素、碳水化合物、总氮含量变化,以期为解决油橄榄不完全花比例大、座果率低的问题提供理论指导。
1 材料与方法
1.1 试验地概况及试验材料
试验地位于四川省金堂县淮口镇西中油橄榄示范园,地处104°20′E,30°29′N,年均温17.3℃,≥10℃有效积温5 450℃,年日照时数1 268h,平均降雨量920mm,无霜期285d,海拔525m,该区内春季气温回升快但不稳定,间有低温和干旱,属亚热带湿润季风区,土壤类型为钙质紫色土。
试验以成都市金堂县西中油橄榄公司5年生座果率高的‘阿贝基娜’与座果率低的‘卡琳’为材料,2个品种采用相同的水肥管理方式,苗木定植株行距均为3m×4m。
1.2 方法
于2015年2月19日到2015年4月23日,每隔7d收集2个品种的花芽样品。花芽主要采集植株东南西北4个不同方位的结果母枝中部的腋芽,对收集的样品分两部分进行处理分别保存,一部分用FAA固定液(70%酒精90mL+冰醋酸5mL+37%-40%甲醛5mL)保存,然后进行石蜡切片,在显微镜下观察其内部形态变化特征,明确‘阿贝基娜’与‘卡琳’2个品种在金堂县进行未分化期、分化初期、花萼形成期、花冠形成期、雄蕊形成期、雌蕊形成期形成的时期,花芽形态分化过程的判断标准以徐纬英[4]报道的油橄榄花芽分化不同期的形态为基础;另一部分称取花芽5g左右,用液氮处理后放于-70℃冰箱,用于测定花芽内源激素与碳水化合物含量。
1.3 测定项目
采用常规石蜡制片法[5]进行花芽形态特征观察。GA3、IAA、ABA均采用高效液相色谱仪测定[6]。总N采用凯氏法测定[7]。碳水化合物包括可溶性总糖及淀粉,采用蒽酮比色法测定[8]。
1.4 数据处理
试验数据运用Excel 2003进行整理和分析。
2 结果与分析
2.1 油橄榄花芽分化期IAA变化
IAA(吲哚乙酸)为植物生长内源激素的一种,既具有促进植物生长的作用,也具有抑制植物生长、促进器官建成、形成花芽的作用[9-10]。由图1、图2可以看出,‘阿贝基娜’的IAA含量变化趋势在2月27日出现大高峰,含量为2.76mg/g,‘卡琳’在3月11日出现大高峰,含量为2.55mg/g,其高峰时间比‘阿贝基娜’晚13d,IAA含量比‘阿贝基娜’低0.21mg/g,在出现高峰后2个油橄榄品种的IAA含量均呈快速下降趋势,而在开花前又快速上升。
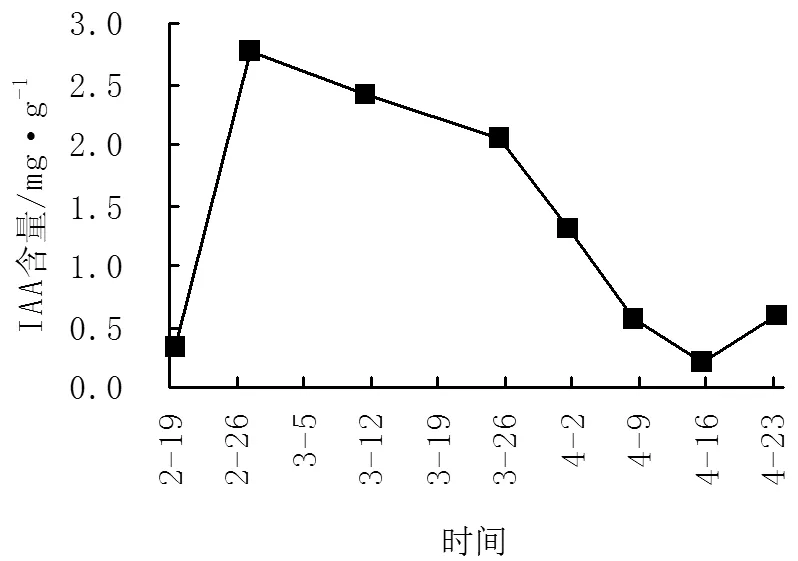
图1 不同时期‘阿贝基娜’IAA含量变化图

图2 不同时期‘卡琳’IAA含量变化图
2.2 油橄榄花芽分化期GA3变化
GA3(赤霉素)也是植物内源激素的一种,主要作用是促进植物伸长生长、解除休眠、促进开花,也可诱导开花结实[9-10]。由图3、图4可知,‘阿贝基娜’GA3含量在花芽分化期出现2个高峰,其中在2月27日-3月11日出现大高峰,GA3含量达85.54-100.45mg/g,此时正处于花芽分化初期。在4月1-8日出现小高峰,此时为花芽各花器官形成期。而‘卡琳’GA3含量在花芽分化期间(2月27日-3月11日)只出现了1个高峰,GA3含量为80.08-84.84mg/g,此时正处于花芽分化初期,低于‘阿贝基娜’此时期GA3含量5.46-15.61mg/g。而4月1-8日之间GA3含量变化不大,此时为花芽各花器官形成期。
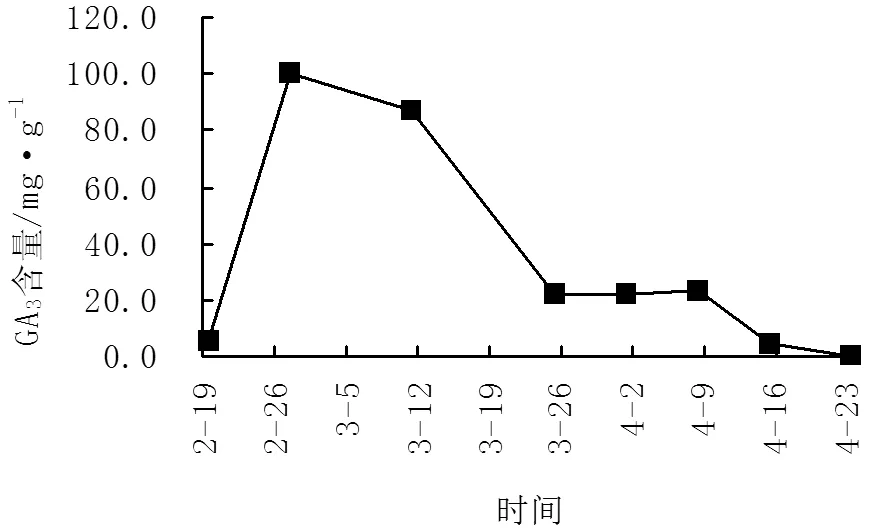
图3 不同时期‘阿贝基娜’GA3含量变化图
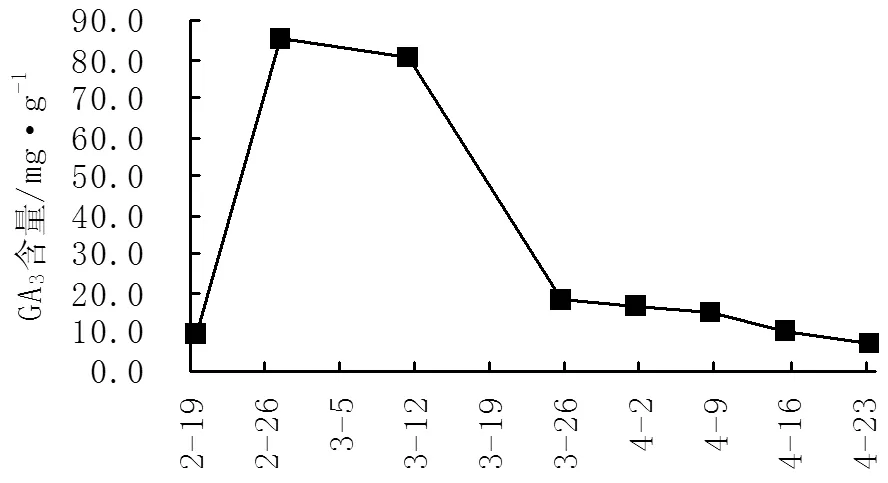
图4 不同时期‘卡琳’GA3含量变化图
2.3 油橄榄花芽分化期ABA变化
ABA(脱落酸)主要作用是引起枝条停止生长,有利于糖分及淀粉的积累[9-10]。由图5、图6可知,‘阿贝基娜’、‘卡琳’2个品种在花芽分化过程中ABA含量均没有检测出,说明在油橄榄花芽分化过程中没有ABA参与分化。

图5 不同时期‘阿贝基娜’ABA含量变化图

图6 不同时期‘卡琳’ABA含量变化图
2.4 油橄榄花芽分化期C/N含量变化
C/N比关系学说由Kraus首先提出 ,他认为果树体内氮和碳水化合物的比例适当,花芽分化旺盛,开花结果也多;但因碳水化合物少引起的C/N比值低,则难成花;因N高引起的C/N比值低,也难成花,枝条容易徒长;因N少引起的C/N比值高,能成花,但座果不良[1]。由表1及图7可知,随着花芽的进一步分化,芽内碳水化合物的含量越来越低,总氮含量越来越高,C/N比值也越来越低。从总体来看,‘卡琳’的C/N比值略低于‘阿贝基娜’,且在花芽过程中C/N比值呈不稳定趋势,分别在3月11日、4月1日左右出现2个拐点,比值为25.42、20.31,分别比‘阿贝基娜’同期C/N比值提高10.14%、11.35%。
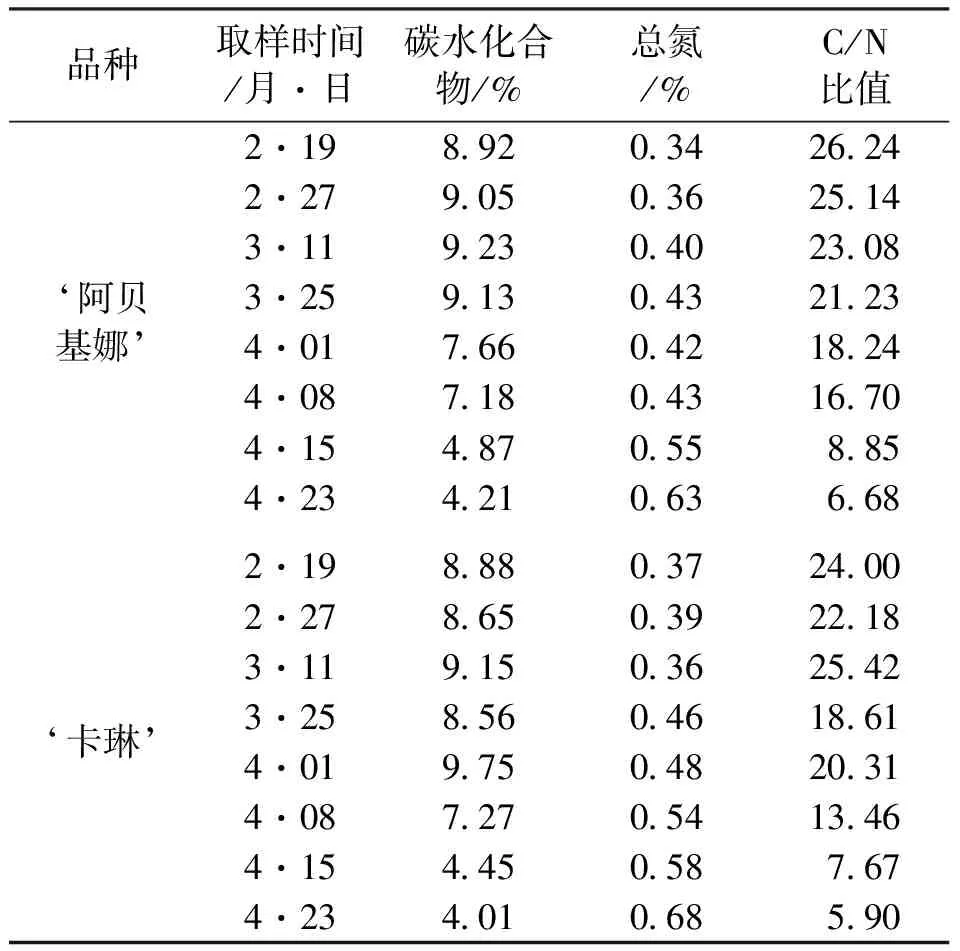
表1 不同分化期芽内C、N含量变化表
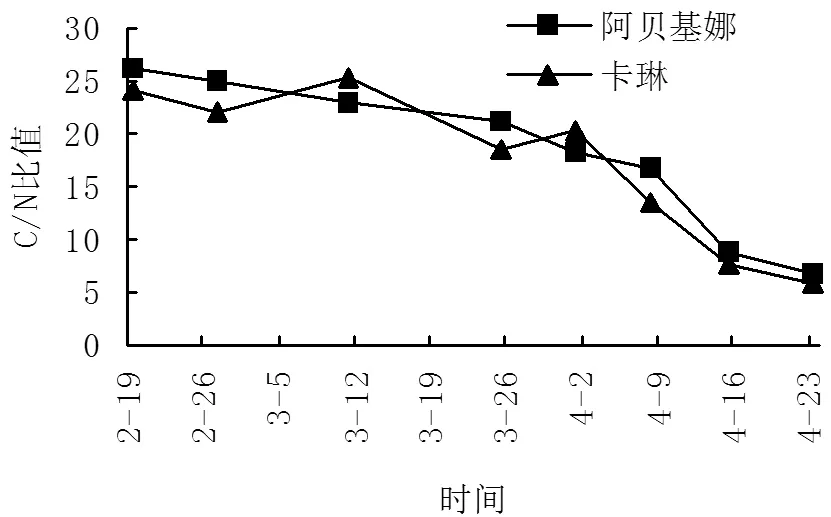
图7 不同分化期C/N比值变化图
3 讨论与结论
植物生长到一定阶段,在适宜的外部环境条件下,通过复杂的生理与形态结构变化,会逐渐转入生殖生长形成花芽[11],在转变过程中,植物激素对花芽分化起重要的调控作用,且这种调控作用是由多种激素综合调节的[12]。根据前人的研究,IAA与GA3对植物成花的作用机理不一致,黄卫东等、吴邦良等、王玉华等[13-15]认为IAA与GA3是花芽形成的抑制因子之一,而史继孔等、黄羌维[16-17]在银杏(GinkgobilobaL.)与龙眼(DimocarpuslonganLour.)中研究认为IAA与GA3对花芽分化起促进作用。究其原因在于IAA与GA3在高浓度时主要作用是促进细胞生长,而在低浓度下促进成花[18]。本试验结果表明,油橄榄在花芽分化前期IAA含量相对较高,之后逐渐降低,而在开花前开始上升,可能是由于油橄榄在开花前需要高浓度的IAA以促进细胞生长完成开花。
油橄榄在整个花芽分化过程中都未检测出ABA含量,与油橄榄成花无直接关系。这与Hoad[19]在苹果(Maluspumila)、梨(Pyrusspp.)上的研究结论一致。
碳水化合物与氮含量同样在花芽分化过程中起着重要作用,是花芽分化的基础。本试验结果表明,在油橄榄花芽生理分化的前期,碳水化合物含量较高,而氮含量相对较低,而碳水化合物能提高细胞液浓度,为花芽的生理分化提供重要的营养物质,有利于促进花芽分化[20],但随着花芽分化的不断进行,碳水化合物含量越来越低,氮含量越来越高,油橄榄进入形态分化阶段,营养生长受到促进。这与钟晓红等[21]的研究结论一致,也符合陈厚彬等的“库一源”理论[22]。根据植物花芽分化C/N比关系学说可知,因为N含量少而引起的C/N比值高则能成花但座果不良,这可能也是在生产中存在‘阿贝基娜’花量大结果量大,而‘卡琳’花量大结果量小的原因之一,因此‘卡琳’在花芽分化过程中需N量高于其它品种,生产过程中特别是花芽分化初期要补充N肥的施用量。
参考文献:
[1]郗荣庭.果树栽培学总论[M].北京:中国农业出版社,2004.
[2]樊卫国,刘国琴,安华明,等.刺梨花芽分化期芽中内源激素和碳、氮营养的含量动态[J].果树学报,2003,20(1):40-43.
[3]李俊霞,黄易,徐骁巍,等.我国及世界油橄榄产业分析[J].消费导刊,2010(1):230-232.
[4]徐纬英.中国油橄榄[M].长春:长春出版社,2001.
[5]李正理.植物制片技术[M].北京:科学出版社,1987.
[6]陈鸿,李智辉,李天来,等.新铁炮百合花芽分化及发育过程中内源多胺及激素含量变化的研究[J].沈阳农业大学学报,2010,41(3):284-288.
[7]汪炳良,邓俭英,曾广文.萝卜花芽分化过程中茎尖和叶片碳水化合物含量的变化[J].园艺学报,2004,31(3):375-377.
[8]文赤夫,董爱文,李国章.蒽酮比色法测定紫花地丁中总糖及还原糖含量[J].现代食品科技,2005,21(3):122-125.
[9]曹尚银,张秋明,吴顺,等.果树花芽分化机理研究进展[J].果树学报,2003,20(5):345-350.
[10]马月萍,戴思兰.植物花芽分化机理研究进展[J].分子植物育种,2003,4(1):539-545.
[11] Rakngan J,Gemma H,Iwahori S.Flower bud formation in Japanese pear trees under adverse conditions and effects of some growth regulators [J] Journal of Japanese Tropical Agriculture,1995,39(1):1-6.
[12]崔薇,吕忠恕.第十二届国际植物生长物质会议简况[J].植物生理学通讯,1986(2):66-72.
[13]黄卫东,原永兵,彭宜.温带果树结实生理[M].北京:北京农业大学出版社,1994.
[14]吴邦良,夏春森,赵宗方.果树开花结实生理和调控技术[M].上海:上海科学技术出版社,1994.
[15] Wang Y H,Fan C H,SHen X.Changes in endogenous hormones during the flower bud differentiation of sweet cherry[J].Acta Agriculturae Boreali—Occidentalis Sinica,2002,11(1):64-67.
[16]史继孔,张万萍,樊卫国,等.银杏雌花芽分化过程中内源激素含量的变化[J].园艺学报,1999,26(3):194-195.
[17]Huang Q W.Changes in endogenous hormone contents in relation to flower bud differentition and on-year or off-year fruiting of Longan[J].Journal of Tropical and Subtropical Botany,1996,4(2):58-62.
[18]马月萍,戴思兰.植物花芽分化机理研究进展[J].分子植物育种,2003,4(1):539-545.
[19]Hoad G V.Hormlnal regulation of fruit-bed formation in fruit rtees [J].Acta Hortic,1984,149:13-23.
[20]罗平源,史继孔,张万萍.银杏雌花芽分化期间内源激素、碳水化合物和矿质营养的变化[J].浙江林学院学报,2006,23(5):532-537.
[21]钟晓红,罗先实,陈爱华.李花芽分化与体内主要代谢产物含量的关系[J].湖南农业大学学报,1999,25(1):1-6.
[22]陈厚彬,黄辉白,刘宗莉.荔枝成花与碳水化合物器官分布的关系研究[J].园艺学报,2004,31(1):1-6.
猜你喜欢
黑龙江粮食(2022年6期)2022-11-23
落叶果树(2020年3期)2020-06-17
河北果树(2020年1期)2020-02-09
中成药(2018年4期)2018-04-26
中国果业信息(2017年8期)2017-09-12
浙江农业科学(2016年11期)2016-05-04
绿色科技(2015年5期)2015-07-05
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
特产研究(2014年4期)2014-04-10
中国果业信息(2014年4期)2014-01-24
