
五个欧李品种叶片光合作用与水分利用效率比较研究
2018-04-21 06:18张世军母冰洁卜芳侠薛安明王露宁
陕西林业科技 2018年1期
张世军,母冰洁,薛 东,卜芳侠,薛安明,王露宁
(1. 韩城市水土保持工作站,陕西 韩城 715400;西北农林科技大学 2.资源与环境学院;3.林学院, 陕西 杨凌 712100)
欧李(Cerasushumilis)又称钙果,为蔷薇科樱属中国特有植物[1]。分布于我国内蒙古、黑龙江、河北、陕西、辽宁、山东、河南、吉林等地,生长于海拔100~1 800 m的地区,常生长在阳坡砂地或山地灌丛中[2-3]。欧李果实中含有各种对人体有益的矿质元素,尤其是钙元素的含量比一般的水果都高,为突出其钙含量高这一特点,其商品名被称为“钙果”[4]。欧李果实可以以提取花青素,对于解决人体亚健康,防治高血压、糖尿病、心脑血管疾病和癌症有很好效果。其果仁提取的高档钙果油,有防治高血压和动脉粥样硬化作用[5-7]。欧李叶和果渣、油渣、果壳和枝条可进一步加工或提取有效成分[8]。栽培方面欧李具有极强的抗旱、抗寒、抗风沙、抗病虫、耐盐碱、耐贫瘠特性[9-12]。它可以耐-35℃低温[13],在荒山、荒坡、丘陵、老化压沙地和弱碱性土壤种植[14-15],也可以在乔木果树行间、梯田地边种植,是荒山绿化、水土保持及治沙、治碱不可多得的优良树种[16]。
目前人工引种栽培刚刚开始,山西农业大学已经筛选出农大3号、农大4号、农大5号、农大6号、农大7号五个栽培品种[18-20],目前已被大量引种栽培[21-22]。本试验以这五个主栽品种为研究对象,研究其叶片光合气体交换参数的日变化,以阐明欧李不同品种光合作用的差异性,为区域筛选欧李栽培品种提供依据。
1 材料与方法
1.1 材料
2014年从山西农业大学园艺研究所引进农大3号、农大4号、农大5号、农大6号和农大7号五个欧李品种作为试验材料,以野生欧李为对照。试验材料为大小基本一致的1年生嫁接苗,苗高70.0 cm,地径为1.2 cm。
试验地位于山西省韩城市板桥乡水土保持试验示范区,为丘陵茆顶地形,年平均气温12.4 ℃,年均降水量520 mm,年平均水面蒸发量为 1 850 mm,年日照时数 233.9 h,年太阳总幅射量507.3 KJ·cm-2,干旱指数2.33。土壤为褐土,pH值 7.4~8.2。
1.2 方法
试验采用单因素随机区组法安排试验,将茆顶进行土地平整,耕作层保留50 cm熟土层。每一个品种栽培面积0.3 hm2,株行距0.3 m×2.0 m。2015年2月底定植苗木,以后按常规方法进行松土除草和病虫害防治等日常管理。2016年8月,用Li-6400光合测定系统标准叶室进行生理生态指标测定。对5个欧李品种和野生种,选择长势良好的植株,随机选取标准枝,选择枝条中部无病虫害和机械损伤的阳向叶片作为测定样叶。每个品种选取3个标准株。在每个标准株上分别选择3枚叶片。测定时间6∶00-20∶00,每隔2 h测定一次。叶片光合气体交换参数:叶片光合气体交换参数包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、细胞间隙CO2浓度(Ci)均用LI-6400型光合仪(美国LI—Cor公司)测定,每个处理重复3次。瞬时水分利用效率为Pn/Tr。
1.3 数据处理
利用Excel和SPSS软件进行数据处理与统计分析。
2 结果分析
2.1 不同欧李品种叶片光合气体交换参数的日变化特征
2.1.1 净光合速率(Pn)日变化规律 光合作用是植物生产过程中物质积累与生理代谢的基本过程,植物叶片光合速率的日变化规律反应了不同植物对环境因子的适应性和对各种环境因子的利用能力[1]。由图1可知,欧李叶片的净光合速率(Pn)日变化呈显二次峰值,即双峰值曲线。Pn在10∶00-11∶00之间达到第一次峰值,也是全天的最大值,随后Pn缓慢下降,在午间表现出一定程度的光合“午休”现象,之后Pn缓慢上升,但升高幅度不大,在15∶00-15∶30之间达到第二次峰值,与第一峰值相比有所降低。最后,Pn再次降低。
光合午休现象作为一种生理调节过程,不同欧李品种峰值的大小有一定的差异(图1),第一次峰值时,农大5号的净光合速率最高,农大7号最低,其他品种处于中间水平。下午16∶00第二次峰值出现时农大3号仍然保持较高水平,而农大7号则相对较低。14∶00净光合速率达到低谷,五个欧李品种的净光合速率均高于对照。
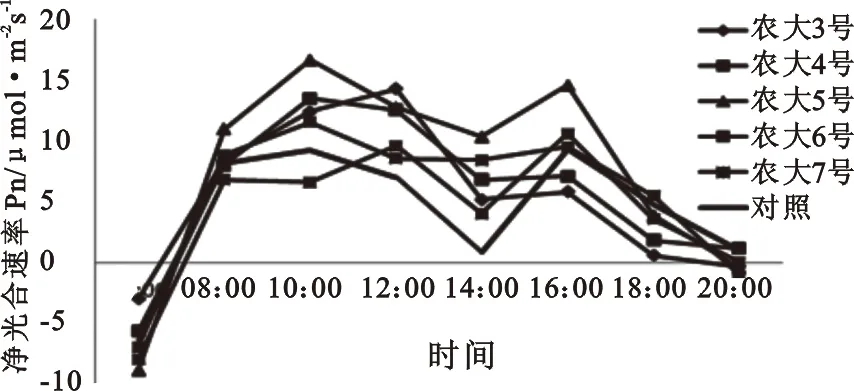
图1 五个欧李品种的叶片净光合速率日变化
就欧李叶片净光合速率日变幅而言,农大3号的净光合速率为14.58 mol·m-2·s-1,4号12.27 mol·m-2·s-1,5号17.06 mol·m-2·s-1,6号12.55 mol·m-2·s-1,7号10.51 mol·m-2·s-1、野生种8.26 mol·m-2·s-1,5个引入品种的日变幅均大于当地野生种的日变幅,具有一定的丰产潜力,而其中“农大5号”日变幅最大,全天净光合速率都较高,说明该品种最能够充分的利用当地的光照条件,具有较大的丰产潜力。
2.1.2 蒸腾速率(Tr)日变化规律 蒸腾是植物的重要生理过程,其主要的作用在于增加植物体对于水分的吸收,增加对于无机离子的吸收和运输以及降低植物的体温[7]。由图2可知,欧李蒸腾作用呈峰值曲线,在12∶00左右达到一天之间的最大值,14∶00时出现“午休”现象,然后蒸腾速率又开始回升,15∶30左右达到第二个峰值,之后蒸腾速率又开始下降。蒸腾速率(Tr)在中午前后相对较高,早晨和傍晚相对保持较低水平。这是植物通过蒸腾扩散水分来降低叶片温度,减轻高温环境对叶片造成的灼伤,表现出对高光照的一种适应。
相比较而言,农大5号的蒸腾速率相对较高,始终保持在较高的水平,12∶00时蒸腾速率达到8.00 mol·m-2·s-1;农大3号的蒸腾速率则相对较低,6∶00和14∶00时期蒸腾速率分别为0.48 mol·m-2·s-1和1.02 mol·m-2·s-1。
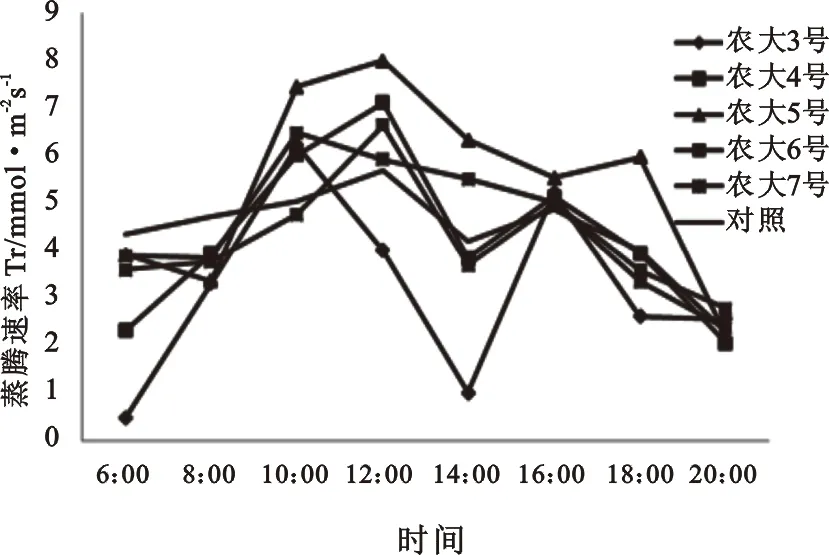
图2 五个欧李品种的叶片蒸腾速率日变化
2.1.3 胞间CO2浓度(Ci)日变化规律 胞间CO2浓度(Ci)一定程度上反应了叶片内光合及呼吸作用的状况,同时也与气孔的开放程度密切相关[12]。由图3可知,欧李Ci在14∶00附近达到最低值,早晨和傍晚相对较高。这是因为傍晚时分欧李的PAR及Gs都处于低水平,不利于进行光合作用;此外夜间光合作用不进行,且植物呼吸作用不断释放出CO2,结果导致傍晚开始Ci逐渐增大,直到早间光合作用开始进行后Ci又缓慢降低。而午间前后由于Pn在这此时开始逐渐升高,细胞内CO2的消耗加大,致使午间前后Ci逐渐下降。
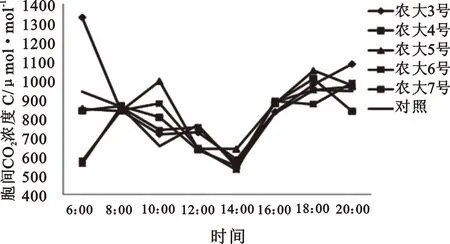
图3 五个欧李品种的叶片胞间CO2浓度日变化
经过对五个欧李品种的比较,6∶00时农大3号胞间CO2浓度相对较高,农大6号和农大7号则处于降低水平。在14∶00达到低谷时,其品种叶片胞间CO2浓度均维持在降低水平。
2.1.4 气孔导度(Gs)日变化规律 气孔是气体进出到植物体内的主要通道,光合作用所需要的CO2以及蒸腾作用所放出的H2O都需要通过气孔进出植物体,因此Gs对植物体的光合作用及蒸腾作用都有着非常重要影响作用。气孔导度受环境因子的影响很大,适宜的光强有利于气孔开张,气孔阻力降低,气孔导度增大[12]。

图4 五个欧李品种的叶片气孔导度日变化
如图4所示,6∶00开始,欧李叶片的欧李叶片的气孔导度(Gs)处于较高水平,随着时间推移逐渐降低,14∶00左右达到最低值,这可能是由于大气中的相对湿度的降低,光合有效辐射的增强以及强烈的叶片蒸腾失水导致气孔部分关闭,气孔阻力逐渐增大,气孔导度降低,净光合速率也随之降低。14∶00以后Gs开始上升,16∶00左右出现第二次峰值,在此之后Gs再次降低。
比较而言,气孔导度达到高峰值时农大5号的气孔导度的为0.29 mol· m-2·s-1,且8∶00-18∶00其气孔导度远远高于其他品种。农大3号的气孔导度变幅较大,14∶00时气孔导度降至最低0.02 mol· m-2·s-1。
2.2 不同欧李品种的水分利用效率(WUE)
水分利用效率是是表示作物水分吸收利用过程效率的一个指标[7,12]。如图5所示,农大3号在8∶00-15∶00有较好的表现,其水分利用效率平均值达到2.43,远高于其他品种。这一时间段光照充足,该品种能较好地利用水分,很好地抵御水分亏缺,应对大气干旱,说明该品种具有一定抵御干旱的潜力。其他品种的水分利用效率表现基本一致,但都在气温最好时(14∶00)高于野生种的0.22。从水分利用效率可以看出,这些经过人工培育的欧李品种在水分利用方面具有明显的优势。
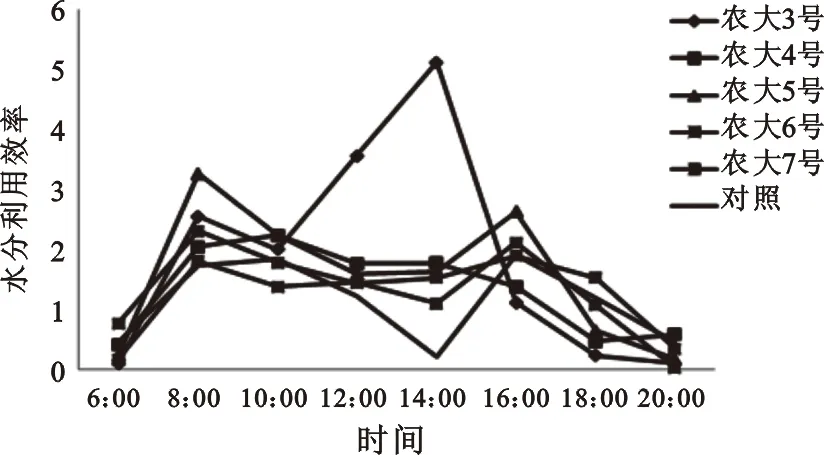
图5 五个欧李品种的叶片水分利用效率
3 结论
五个欧李品种除农大3-7号的叶片的光合日进程表现为不规则的双峰型,光合作用有较明显的光合“午休”现象。净光合速率的第一峰值出现在11∶00左右,第二峰值出现在15∶30左右,第一峰值时的净光合速率最大。
五个品种欧李叶片的净光合速率净光合速率平均值分别为5.46、5.69、7.51、5.58、4.20、4.27 mol·m-2·s-1,除农大7号外其余四个品种都大于野生种,农大5号和农大3号的光合作用性能较好,具有较为明显的光合作用优势。因此,试验地引入的欧李品种大多数具有一定的丰产潜力,欧李的栽培将进一步丰富黄土高原丘陵区水土保持树种资源。
参考文献:
[1] 尹赜鹏,刘雪梅,商志伟,等.不同干旱胁迫下欧李光合及叶绿素荧光参数的响应[J].植物生理学报,2011,47(5):452-458.
[2] 母洁冰,张世军,卜芳侠,等.韩城市欧李新品系引进试验[J].陕西林业科技,2016(4):41-45.
[3] 王鹏飞,曹琴,何永波,等.欧李果实发育期糖和酸组分及其含量的动态变化特性[J].西北植物学报,2011,31(7):1411-1416.
[4] 张世军,母冰洁,褚翔, 等.欧李新品种“农大3号”光合特性研究[J].陕西林业科技,2017(4):10-13.
[5] 郭刚,杜艳丰,石强, 等.盐胁迫条件下对欧李生长与生理特性的影响[J].新疆农业科学,2013(3):471-475.
[6] 王鹏飞,薛晓芳,穆晓鹏, 等.不同酸度欧李果实有机酸积累特性与相关代谢酶活性分析[J].中国农业科学,2013,46(19):4101-4109.
[7] 姜英淑,陈书明,王秋玉, 等.干旱胁迫对2个欧李种源生理特征的影响[J].林业科学,2009,45(6):6-10.
[8] 李学强,李秀珍.盐碱胁迫对欧李叶片部分生理生化指标的影响[J].西北植物学报,2009,29(11):2288-2293.
[9] 刘淑琴,常虹,周家华, 等.我国欧李的开发应用研究现状[J].食品研究与开发,2009,30(12):167-170.
[10] 马建军,张立彬,刘玉艳, 等.野生欧李生长期组织器官中不同形态钙含量的变化及其相关性[J].园艺学报,2008,35(5):631-636.
[11] 张红,张艳敏,陈晓流, 等.欧李自交不亲和S基因的克隆及序列分析[J].西北植物学报,2008,28(5):876-881.
[12] 褚建民,孟平,张劲松, 等.土壤水分胁迫对欧李幼苗光合及叶绿素荧光特性的影响[J].林业科学研究,2008,21(3):295-300.
[13] 孙新政,申顺先,李庆伟, 等.钙果4号欧李组织培养技术研究[J].果树学报,2007,24(1):80-83.
[14] 马建军,张立彬,于凤鸣, 等.野生欧李果实中不同形态钙的含量及分布[J].园艺学报,2007,34(3):755-759.
[15] 周家华,兰彦平,姚砚武, 等.欧李果汁加工工艺研究[J].食品工业科技,2007,(8):146-147,150.
[16] 张美莉,邓秋才,杨海霞, 等.内蒙古欧李果肉和果仁中营养成分分析[J].氨基酸和生物资源,2007,29(4):18-20.
[17] 李学强,李秀珍,司凤云, 等.不同贮藏条件及生长调节剂对欧李花粉生活力的影响[J].西北植物学报,2007,27(11):2251-2256.
[18] 马建军,张立彬.野生欧李生长期矿质营养元素含量的变化[J].园艺学报,2004,31(2):165-168.
[19] 马建军,于凤鸣,张立彬, 等.野生欧李果实发育期不同叶位叶片中主要矿质元素含量的变化[J].果树学报,2010,27(1):34-38.
[20] 李治国,王有信,王玉峰, 等.欧李加工品种与贮藏加工利用[J].农产品加工(学刊),2005(3):21-24.
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
落叶果树(2021年1期)2021-01-29
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
北京农学院学报(2019年2期)2019-05-09
