
大气细颗粒物暴露后小鼠肺组织microRNA差异表达谱芯片分析
2018-04-19 00:52侯天芳廖纪萍马元元张成王广发
生态毒理学报 2018年1期
侯天芳,廖纪萍 ,马元元 ,张成 ,王广发,*
1. 北京大学第一医院呼吸和危重症医学科,北京 100034 2. 北京大学第一医院实验动物中心,北京 100034
随着我国工业和现代化的快速发展,交通机动车尾气污染物排放量持续增高,大气污染危害成分中首当其冲的是大气颗粒物,特别是可入肺的细颗粒物(空气动力学当量直径≤2.5 μm的颗粒物,PM2.5),对人群危害风险损伤强度要远高于臭氧(O3)、二氧化硫(SO2)等气态污染物,成为对人群危害风险威胁最大、代表性最强的大气污染物。流行病学已经证实PM2.5与心肺呼吸系统疾病的发病率和病死率有明显的相关关系[1-2]。权威调查显示,在中国室内和室外空气污染导致2 500万多人健康寿命损失和120万人过早死亡,但是PM2.5引起机体损害的具体机制目前尚不十分清晰[3]。近来有关研究发现,PM2.5在中枢神经系统的致病中亦发挥作用[4-5],甚至与神经退行性疾病、精神性疾病、帕金森病等疾病的发生发展亦有相关[6-8]。
MicroRNAs是一类长度为19~23 nt的短序列、非编码、具有调控功能的单链小分子RNA。广泛存在于各种细胞中,主要参与真核生物的发育、生长、分化、免疫反应和适应压力等多种生理过程[9]。近年来,研究发现microRNAs在环境化学物质引起的毒理学过程中起着十分重要的作用[10]。实验研究发现,空气污染能够主要引起microRNA-9,microRNA-223,microRNA-10b,microRNA-143等miRNAs的改变[11],不管是体内还是体外实验均发现暴露于炭黑颗粒物、颗粒物、柴油机废弃颗粒物后,microRNAs表达会发生明显的改变[12-15]。
MicroRNAs稳定存在于血液及组织中,因此作为一种临床早期诊断的新标记物逐渐受到重视,本研究通过PM2.5暴露后建模小鼠肺组织芯片筛选,找到microRNAs差异表达谱,选用三大数据库进行交集分析预测靶基因,根据生物信息学做GO和KEGG通路分析,从而预测及分析大气污染相关疾病的作用机制通路,为后续科研实验奠定理论依据。
1 材料与方法(Materials and methods)
1.1 实验材料
1.1.1主要实验试剂材料
实验用microRNA提取、逆转录试剂盒以及RT-qPCR反应试剂盒均购自北京全式金生物技术有限公司;ABI 7500荧光定量PCR仪。
1.1.2PM2.5颗粒的采集、提取和悬液制备
颗粒物的采样点为北京大学第一医院内科楼楼顶,应用Staplex PM2.5大流量采样器,于2016年1月连续采样96 h。将附有PM2.5颗粒的玻璃纤维滤膜剪成小块于烧杯;加入无菌超纯水,低温超声以混匀悬浮液,使用6层纱布过滤悬液,低温冷冻干燥收获颗粒物于-20 ℃冰箱备用。根据实验小鼠的体重称取PM2.5颗粒物,于动物实验前一天用生理盐水配制不同浓度的细颗粒物混悬液,超声振荡混匀,4 ℃冰箱保存备用,使用前再次超声振荡混匀。
1.2 实验方法
1.2.1实验动物及处理
实验动物为北京维通利华实验动物技术有限公司(SCXK(京)2012-0001)提供的SPF级8周龄雄性BALB/c小鼠(25±2 g),在北京大学第一医院实验动物中心(SYXK(京)2014-0010)饲养。小鼠自由进食消毒颗粒饲料及水,室温23~25 ℃,12 h光:12 h暗,湿度25%~30%。通过了实验动物福利伦理审查(动物伦理编号:J201609),小鼠随机分为生理盐水组、低剂量暴露组(PM2.5,2.5 mg·kg-1)、高剂量组(PM2.5,20 mg·kg-1),每组6只,建模方法参考本课题组既往研究[16]采用气管滴注方法进行,用5%水合氯醛麻醉处理,麻醉后的小鼠仰卧固定于操作板,借助冷光源透视,清晰看到声门裂,用小型注射器与改装后的留置针吻合固定,进行快速气管滴注(滴注量为50 μL·次-1),一周滴注2次,气管滴注14 d结束后过量麻醉处死动物并取材。
1.2.2小鼠肺组织提取RNA并芯片处理
随机从3组中选出3只小鼠,获取肺组织后提取RNA,样品总RNA利用NanoDrop ND-2000 (Thermo Scientific)定量,检测RNA完整性使用安捷伦生物分析软件,样本检测合格后进行芯片处理,严格按照操作流程进行(委托上海欧易公司进行芯片检测)。
1.2.3实时荧光定量PCR验证
严格按照北京全式金生物技术有限公司操作说明书进行microRNA的提取、逆转录和RT-qPCR实验。逆转录参数条件设置为37 ℃、60 min,85 ℃、5 s灭活。后续进行qRT-PCR,设置反应条件为50 ℃、2 min 灭活UNG 酶,94 ℃、30 s 预变性,94 ℃、5 s,60 ℃、34 s,共40个循环。每个标本均采用三复孔。表达水平以2-ΔΔCT表示,引物序列如下:
mmu-miR-139-5p:5’-TCTACAGTGCACGTGTCTCCAG-3’;mmu-miR-691:5’-ATTCCTGAAGAGAGGCAGAAAA-3’; mmu-miR-340-3p:5’-TCCGTCTCAGTTACTTTATAGC-3’;U6 forward:5'-CTCgCTTCggCAgCACA-3'; U6 reverse:5'-AACgCTTCACgAATTTgCgT-3'。
1.3 生物信息学分析(GO和KEGG分析)
应用TargetScan,PITA,microRNAorg这三大数据库进行microRNAs靶基因预测,输入差异表达的miRNAs,取三大软件共同预测到的靶基因的交集,再对靶基因进一步进行GO分析,主要包括三大部分,分别为生物过程(biological process)、分子功能(molecular function)、细胞组分(cellular component),并采用KEGG分析主要参与的信号通路,综合分析富集结果。其中利用T检验得到的差异显著性P值和标准化信号值的差异倍数fold change值进行筛选,标准为fold change≥2.0且P≤0.05。
2 结果(Results)
2.1 肺组织提取RNA纯度测定
结果显示各组RNA样本的OD260/OD280比值均在1.8~2.2之间且28S/18S≥0.7,28S rRNA 和 18S rRNA 条带清晰,5S rRNA条带很弱,经Agilent Bioanalyzer 2100 (Agilent Technologies)检测RNA完整性好,达到芯片实验上机要求。
2.2 PM2.5暴露后小鼠肺组织microRNAs变化
大气细颗粒物染毒后小鼠肺组织microRNAs有显著差异变化,表1结果显示,高剂量处理组与对照组比较有4个microRNAs上调,低剂量处理组与对照组比较有2个microRNAs上调,高剂量组与低剂量组比较,有4个miRNAs上调(标准为fold change值>= 2.0且P值<= 0.05)。
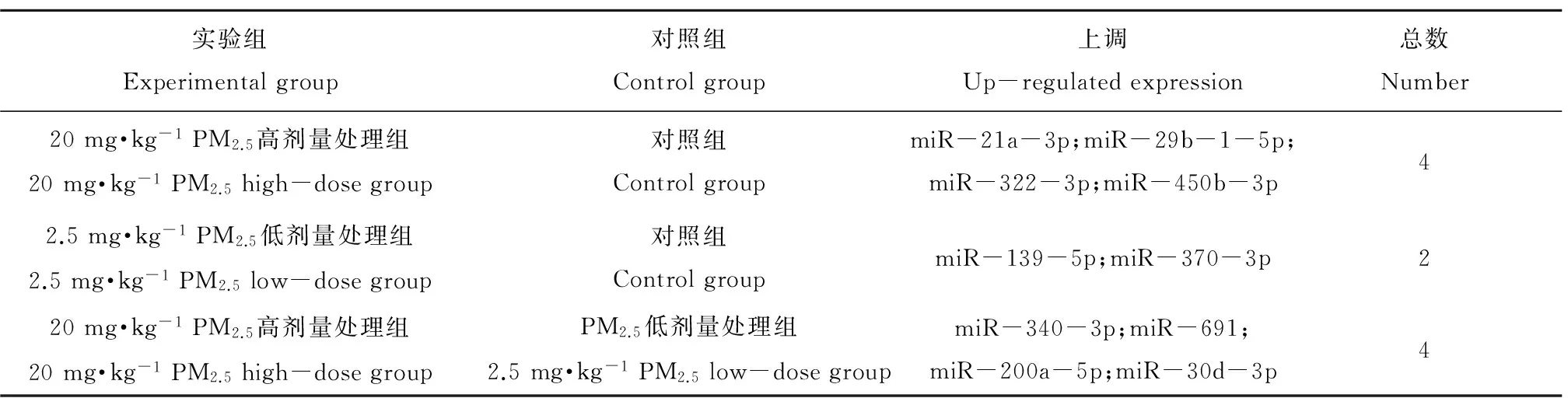
表1 PM2.5暴露对肺组织microRNAs的影响Table 1 microRNAs alterations in mice lung tissue after PM2.5 exposure
2.3 实时荧光定量PCR(qRT-PCR)验证差异表达的microRNAs
实时荧光定量PCR验证差异表达的miRNAs的结果如图1所示。
2.4 差异表达明显的microRNAs靶基因预测结果
经芯片筛查发现10个差异表达的microRNAs,其中综合考虑上调的倍数以及2组进行两两比较的结果,利用TargetScan,PITA,microRNAorg数据对差异miRNA进行靶基因预测,其中有4个miRNAs可以预测到靶基因,分别是miR-139-5p、miR-691、miR-340-3p及miR-450b-3p,具体预测靶基因结果见表2及图2。

表2 microRNAs 靶基因预测结果Table 2 Results of microRNAs targets
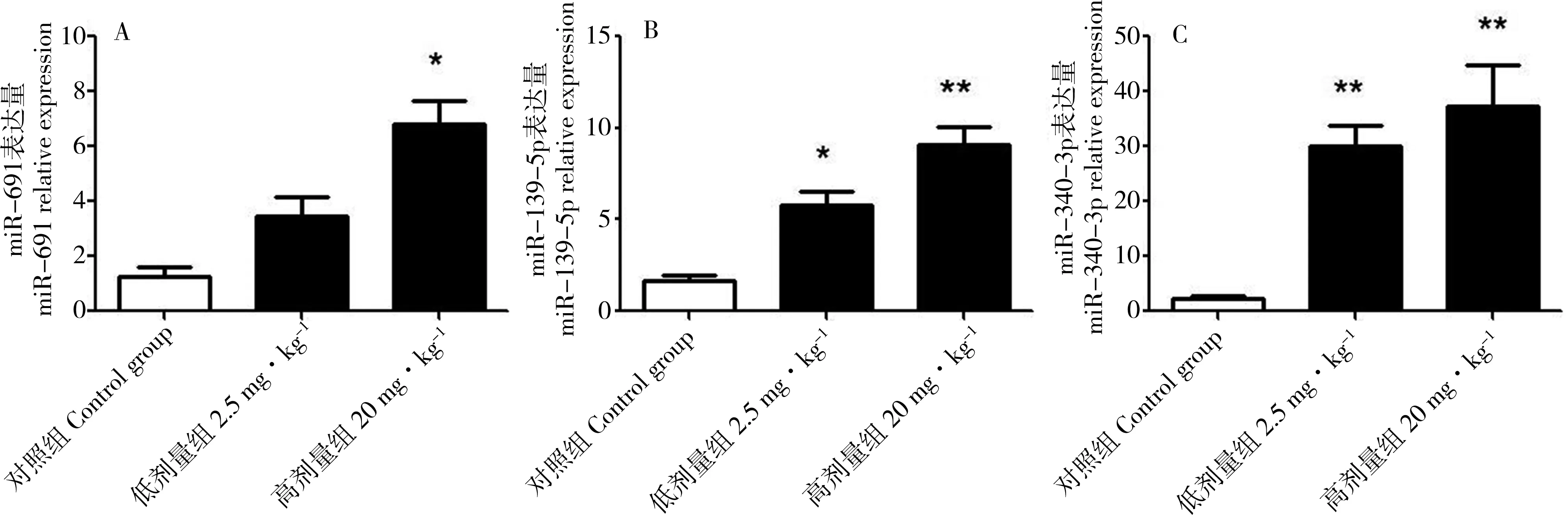
图1 差异表达的miRNAs的qRT-PCR验证结果(n=6)注:与对照组比较 *P<0.05, **P<0.01。Fig. 1 Validation of differentially expressed microRNAs (n=6)Note: *P<0.05, **P<0.01, compared with the control group.
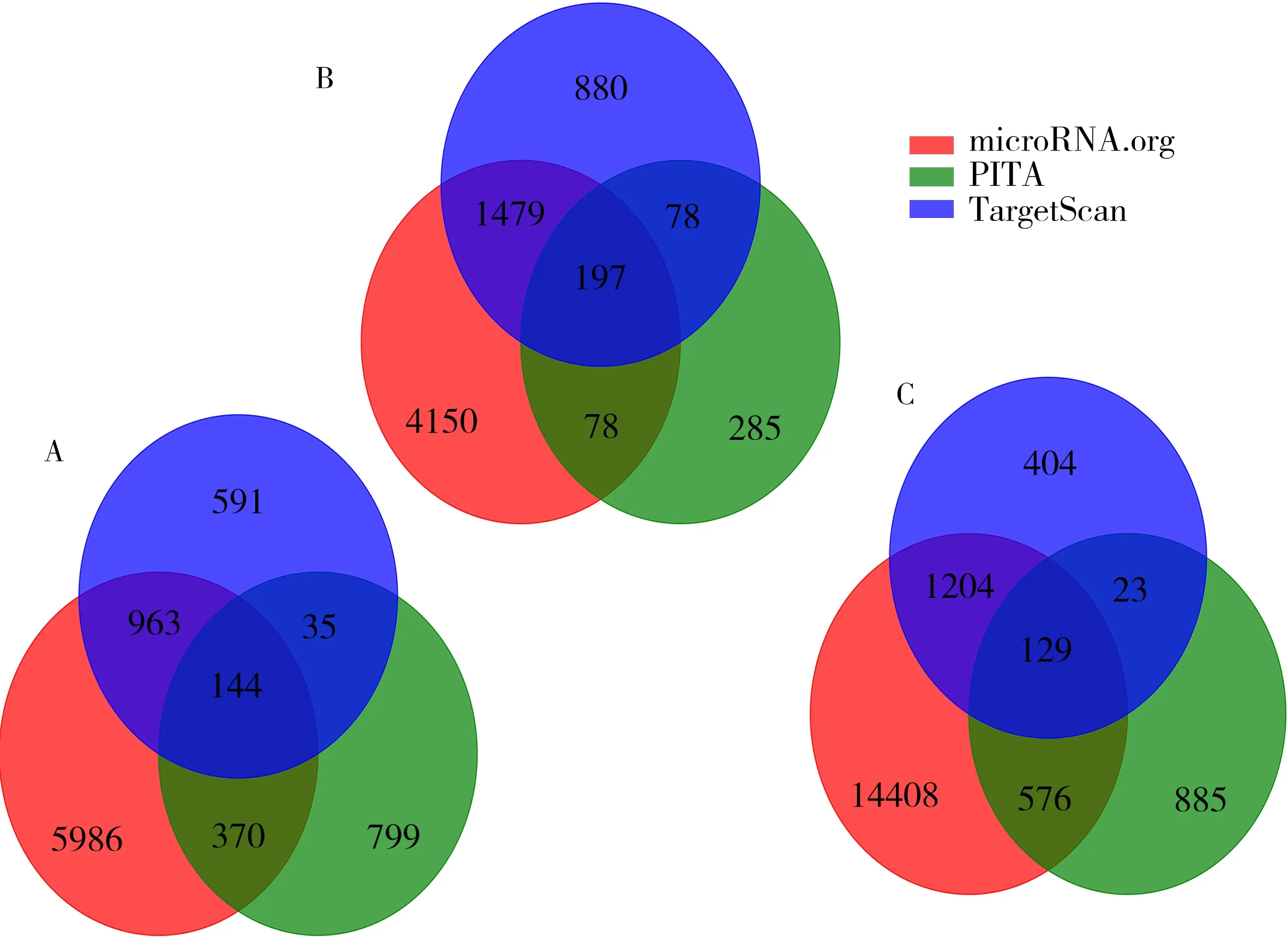
图2 3个数据库预测到的靶基因交集图注: A, miR-139-5p预测靶基因数; B, miR-691及miR-340-3p预测靶基因数; C, miR-450b-3p预测靶基因数。Fig. 2 The target gene intersection map predicted by three databasesNote: A, results of miR-139-5p targets; B, results of miR-691 and miR-340-3p targets; C, results of miR-450b-3p targets.
2.5 差异表达miRNAs靶基因显著富集GO和KEGG分析结果
差异表达miRNAs靶基因显著富集GO和KEGG分析结果如表3和图3所示。
3 讨论(Discussion)
2013年以来,以PM2.5为主要污染成分的雾霾频发,是我国目前最受关注的重大环境问题之一,北京灰霾污染尤甚。本研究的PM2.5采集于北京冬季雾霾高发期(位于北京大学第一医院内科楼顶楼),尽管大气颗粒物成分复杂,表面附着多种有毒物质,但本研究采用大气颗粒物染毒小鼠探索分子毒理学机制过程有其特有的参考价值。
研究证实,miRNAs与其靶mRNA的3′端非编码区(3'-UTR)互补结合,在翻译水平上特异性抑制基因表达[11]。实验研究发现,约60%的编码蛋白基因是受microRNAs调控的[17],值得注意的是,microRNA很可能与很多种不同的靶基因有调控关系[18]。有研究发现,长时间大气颗粒物暴露后人体外周血中microRNAs(let-7g-5p,miR-126-3p,miR-130a-3p,miR-146a-5p,miR-150-5p, miR-191-5p和 miR-23a-3p)发生变化,并实验验证其靶基因(CCR3,CD40,COL1A2,CSF1,CXCL12,CXCL8,CXCR4,IL1F10,IL36A,IL36B,IL36G,IL36RN,IL37,IL6,PDGFB和VCAM1)参与了心血管疾病相关的信号通路[19]。
第一个人类全基因组草图已经于2010年完成,目前对于基因组的研究也进入了功能基因组时代,即后基因组时代。生物信息学中的GO分析可指导不同样本的差异基因可能和哪些基因功能的改变有关,而KEGG则着重于找出广泛参与的信号通路,确定参与发生发展过程的差异表达基因[20-21]。目前针对软件预测而得的靶基因群,最常用的生物信息学方法包括:GO (Gene Ontology) 和KEGG分析 (the Kyoto analysis of the Encyclopedia of Genes and Genomes)通路分析。实验发现差异表达的miRNAs所调控的靶基因显著富集在34个GO通路(P<0.001),具体为15个GO涉及到生物过程,主要包括:RNA拼接,RNA转录酶II启动子的转录,DNA模板,转录调节、激活JNKK活动等;9个GO涉及细胞组分,主要包括:细胞核、质、膜及突触等;10个GO涉及分子功能,主要包括:蛋白结合、RNA和DNA结合、RNA聚合酶II启动子结合、R-SMAD 结合等。将差异表达的miRNAs进行Pathway富集分析,有助于找到实验条件下显著性差异变化的生物学调控通路。在Pathway分析(P<0.05,见图3 A B C)中,3组进行通路比较,富集于轴突导向通路值得关注,还有突触囊泡循环、五羟色胺神经突触、神经营养因子信号通路、磷脂酰肌醇信号系统等,都提示差异miRNAs富集信号通路指向中枢神经发育以及神经退化性疾病的发生发展。生长锥是位于神经突起末端的一个扇形结构,其受体可选择性识别某些导向信号,通过特定的信号转导通路发挥生物学反应,进而指导轴突识别达到正确的位置,可见轴突导向是神经发育的一个重要分支[22]。Bellon等[23]发现,miR-182在体内外可调控由Slit2介导的视网膜神经节细胞的轴突导向,且生长锥的miR-182抑制cofilin-1局部蛋白复合体的转录翻译。其中,靶基因Arhgef12、Sema4g、Epha5、Ablim1参与了这个轴突导向重要过程,EphA5是Ephrins家族中新发现的成员,其配体是Ephrin-A5[24]。Cooper等[25]研究发现,EphA5与其配体Ephrin-A5的相互作用与中脑DA神经元的轴突生长有关。研究发现,在坐骨神经损伤的大鼠,在损伤部位的miR-340表达失调影响去除细胞碎片和轴突再生[26]。而miR-691和miR-450与神经系统疾病的相关性鲜见报道。
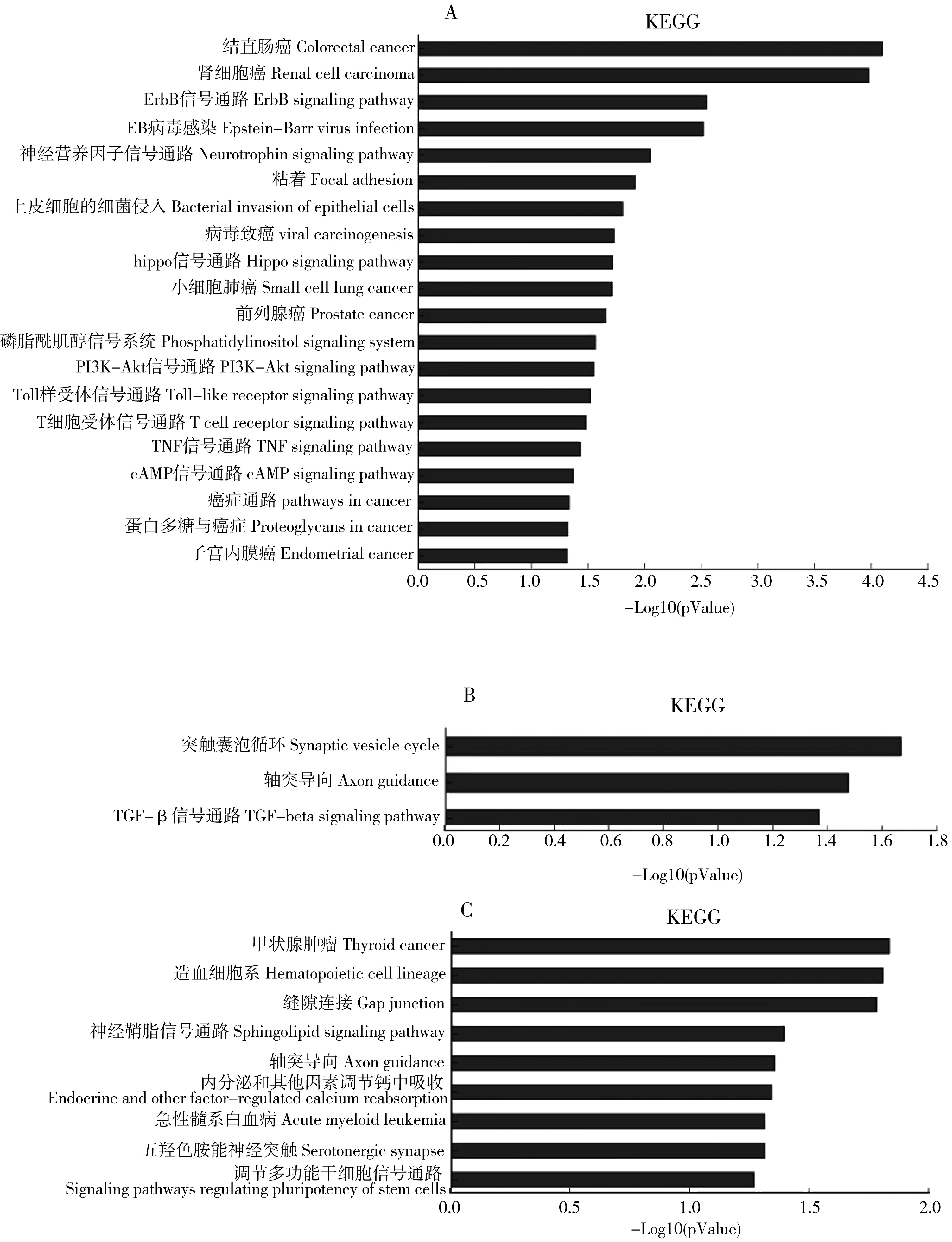
图3 PM2.5暴露后小鼠肺组织差异表达microRNAs靶基因KEGG分析和轴突导向通路注:A为PM2.5低剂量组与对照组比较的差异表达miRNAs靶基因KEGG分析;B为PM2.5高剂量组与PM2.5低剂量组比较的差异表达miRNAs靶基因KEGG分析;C为PM2.5高剂量组与对照组比较的差异表达miRNAs靶基因KEGG分析。Fig. 3 KEGG analysis and axon guidance pathway of differentially expressed lung tissue microRNAs after exposed to PM2.5Note: A, KEGG analysis of differentially expressed microRNAs in low dose group compared to the control group; B, KEGG analysis of differentially expressed microRNAs in high dose group compared to low dose group; C, KEGG analysis of differentially expressed microRNAs in high dose group compared to the control group.
本研究KEGG分析还富集于几条癌症通路,包括:结直肠癌、肾细胞癌、甲状腺癌、急性髓系白血病、前列腺癌等,提示差异表达microRNAs参与癌症通路的某个作用阶段,发挥致病作用。Miyoshi等[27]研究发现在结直肠癌患者中检测到miR-139-5p的升高,并且显著缩短无复发生存率,小鼠模型中发现miR-139-5p的过表达可以加重结直肠患者的腹膜扩散。然而,Wang等[28]分别采用正常结肠黏膜、结肠腺瘤和结直肠癌患者的组织取样进行microRNAs深度测序分析,结果发现与对照组相比,结直肠癌患者中miR-139-5p,miR-204-5p,miR-125b-5p,miR-100-5p和miR-30a-5p表达是明显下调的。再者研究显示,miR-124,miR-137和miR-340通过抵消Warburg效应来修复结直肠癌的生长,主要由于调节替代PKM基因剪接来完成[29]。也有研究证实:可用miR-10b,miR-139-5p,miR-130b 和 miR-199b-5p来检测肾透明细胞癌的转移情况,这4个microRNAs与肾透明细胞癌的转移和预后密切相关,可以作为生物标记物来检测预后[30]。在间充质干细胞分化为胰腺β细胞中,研究发现microRNAs(包括:miR-18a-5p,miR-19b-5p,miR-30d-3p,miR-124,miR-146a-5p, miR-184;miR-335和 miR-433-5p)的过表达是和胰腺β细胞的分化相平衡的[31]。
本文重点研究并验证了大气污染暴露后小鼠肺组织中发现差异表达明显的miR-691、miR-139-5p、miR-340-3p及miR-450,利用生物信息学分析揭示了其改变与神经系统发育、多种癌症通路、调控多能干细胞分化等通路有相关性,为进一步深入研究大气颗粒物引起的毒理学机制提供坚实的科学证据。
参考文献(References):
[1]Lipsett M J, Ostro B D, Reynolds P, et al. Long-term exposure to air pollution and cardiorespiratory disease in the California teachers study cohort [J]. American Journal of Respiratory and Critical Care Medicine, 2011, 184: 828-835
[2]Araujo J A, Nel A E. Particulate matter and athero sclerosis: Role of particle size, composition and oxidative stress [J]. Particle and Fibre Toxicology, 2009, 6: 24-30
[3]Salomon J A, Wang H, Freeman M K, et al. Healthy life expectancy for 187 countries, 1990~2010: A systematic analysis for the global burden disease study 2010 [J]. The Lancet, 2013, 380(9859): 2144-2162
[4]Ku T, Ji X, Zhang Y, et al. PM2.5, SO2and NO2co-exposure impairs neurobehavior and induces mitochondrial injuries in the mouse brain [J]. Chemosphere, 2016, 163: 27-34
[5]Cliff R, Curran J, Hirota J A, et al. Effect of diesel exhaust inhalation on blood markers of inflammation and neurotoxicity: A controlled, blinded cross over study [J]. Inhalation Toxicology, 2016, 28(3): 145-153
[6]Pun V C. Association of ambient air pollution with depressive and anxiety symptoms in older adults: Results from the NSHAP study [J]. Environmental Health Perspectives, 2017, 125(3): 342 - 348
[7]Morales-Suárez-Varela M, Peraita-Costa I. Systematic review of the association between particulate matter exposure and autism spectrum disorders [J]. Environmental research, 2017, 153: 150-160
[9]Van Rooij E, Sutherland L B, Qi X, et al. Control of stress-dependent cardiac growth and gene expression by a microRNA [J]. Science (New York, N.Y.), 2007, 316(5824): 575-579. DOI: 10.1126/science.1139089.
[10]Li X, Lv Y, Gao N, et al. microRNA-802/Rnd3 pathway imposes on carcinogenesis and metastasis of fine particulate matter exposure [J]. Oncotarget, 2016, 7(23): 35026-35043
[11]Vrijens K, Bollati V. MicroRNAs as potential signatures of environmental exposure or effect: A systematic review [J]. Environmental Health Perspectives, 2015, 123(5): 399-411
[12]Jardim M J, Fry R C, Jaspers I, et al. Disruption of microRNA expression in human airway cells by diesel exhaust particles is linked to tumorigenesis-associated pathways[J]. Environmental Health Perspectives, 2009, 117: 1745-1751
[13]Bleck B, Grunig G, Chiu A, et al. MicroRNA -375 regulation of thymic stromal lymphopoietin by diesel exhaust particles and ambient particulate matter in human bronchial epithelial cells [J]. Journal of Immunology, 2013, 190: 3757-3763
[14]Farraj A K, Hazari M S, Haykal-Coates N, et al. ST depression, arrhythmia, vagal dominance, and reduced cardiac micro-RNA in particulate exposed rats[J]. American Journal of Respiratory Cell and Molecular Biology, 2011, 44: 185-196
[15]Bourdon J A, Saber A T, Halappanavar S, et al. Carbon black nanoparticle intratracheal installation results in large and sustained changes in the expression of miR-135b in mouse lung [J]. Environmental and Molecular Mutagenesis, 2012, 53: 462-468
[16]Zhao C, Liao J, Chu W, et al. Involvement of TLR2 and TLR4 and Th1/Th2 shift in inflammatory responses induced by fine ambient particulate matter in mice [J]. Inhalation Toxicology, 2012, 24(13): 918-927
[17]Lewis B P, Burge C B, Bartel D P. Conserved seed pairing, often flanked by adenosines, indicates that thousads of human genes are microrna targets [J]. Cell, 2005, 20: 15-20
[18]Esteller M. Non-coding RNAs in human disease [J]. Nature Reviews Genetics, 2011, 12: 861-874
[19]Rodosthenous R S, Coull B A, Lu Q, et al. Ambient particulate matter and microRNAs in extracellular vesicles: A pilot study of older individuals [J]. Particle and Fibre Toxicology, 2015, 13(1): 1-13
[20]Kanehisa M, Goto S, Sato Y, et al. KEGG for interpretation of large-scale molecular data sets[J]. Nucleic Acids Research, 2012, 40: D109-D114
[21]Ashburner M, Ball C A, Blake J A, et al. Gene ontology: Tool for the unification of biology [J]. The Gene Ontology Consortium Nature Genetics, 2000, 25(1): 25-29
[22]Rajasekharan S, Kennedy T E. The netrin protein family[J]. Genome Biology, 2009, 10(9): 100-105
[23]Bellon A, Iyer A, Bridi S, et al. MiR-182 regulates slit2-mediated axon guidance by modulating the local translation of a specific mRNA [J]. Cell Reports, 2017, 18(5): 1171-1186
[24]Birgbauer E, Cowan C A, Sretavan D W. Kinase independent function of EphB receptors in retinal axon pathfinding to the optic disc from dorsal but not ventral retina[J]. Development (Cambridge, England),2000,127(6):1231-1241
[25]Cooper M A, Kobayashi K, Zhou R. Ephrin-A5 regulates the formation of the ascending midbrain dopaminergic pathways [J]. Developmental Neurobiology, 2009, 69(1): 36-46
[26]Li S, Zhang R, Yuan Y. MiR-340 regulates fibrinolysis and axon regrowth following sciatic nerve injury [J]. Molecular Neurobiology, 2016, 54(6): 1-11
[27]Miyoshi J, Toden S, Yoshida K, et al. MiR-139-5p as a novel serum biomarker for recurrence and metastasis in colorectal cancer [J]. Scientific Reports, 2017, 7: 43393
[28]Wang X, Chen L, Jin H, et al. Screening miRNAs for early diagnosis of colorectal cancer by small RNA deep sequencing and evaluation in a Chinese patient population[J]. Oncotargets and Therapy, 2016, 9: 1159-1166
[29]Sun Y, Zhao X, Zhou Y, et al. miR-124, miR-137 and miR-340 regulate colorectal cancer growth via inhibition of the Warburg effect [J]. Oncology Reports, 2012, 28(4): 1346-1352
[30]Wu X, Weng L, Li X, et al. Identification of a 4-microRNA signature for clear cell renal cell carcinoma metastasis and prognosis [J]. PLoS one, 2012, 7(5): e35661
[31]Coskun E, Ercin M. The role of epigenetic regulation and pluripotency-related microRNAs in differentiation of pancreatic stem cells to beta cells [J]. Journal of Cellular Biochemistry, 2018, 119(1): 455-467
猜你喜欢
学苑创造·A版(2020年9期)2020-10-13
小学生学习指导(低年级)(2017年10期)2017-10-10
环境保护与循环经济(2017年2期)2017-09-26
环境科技(2016年1期)2016-11-08
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
中国病理生理杂志(2015年8期)2015-12-21
化工进展(2015年3期)2015-11-11
医学研究杂志(2015年3期)2015-06-10
浙江大学学报(工学版)(2015年1期)2015-03-01
创业家(2015年1期)2015-02-27
