
黔金丝猴研究进展
2018-04-17 07:38潘有福赵彦禹张志敏陈忠杰
生物学杂志 2018年2期
潘有福, 赵彦禹, 郑 翔, 张志敏, 陈忠杰, 徐 丹
(遵义医学院 遗传学教研室, 遵义 563000)
黔金丝猴(Rhinopithecusbrelichi)又名灰金丝猴、白肩仰鼻猴、牛尾猴、线狨和猓然兽,是国家Ⅰ级重点保护野生动物, 被IUCN 濒危物种红色名录列为“EN”(濒危) ,CITES 附录Ⅰ物种[1-2]。黔金丝猴隶属灵长目(Primates)猴科(Cercopithecidae)疣猴亚科(Colobinae)仰鼻猴属(Rhinopi-thecus)。一般认为,仰鼻猴属现有5个种:黔金丝猴、川金丝猴(Rhinopithecusroxellana)、滇金丝猴(Rhinopithecusbieti)、越南金丝猴(Rhinopithecusavunculus)和怒江(缅甸)金丝猴(Rhinopithecusstrykeri)。其中黔金丝猴、川金丝猴和滇金丝猴是中国特有种。怒江金丝猴于2010年首次在缅甸被发现,后来在中国云南怒江也有发现。化石证据显示金丝猴在第四纪更新世时期在中国及周边邻近国家广泛生存。但目前它们分布范围十分狭小,仅分布在中国境内四川、云南、贵州、湖北、陕西、甘肃、西藏等省和自治区,以及越南北部和缅甸东北部局部区域。其中,黔金丝猴仅分布于贵州西北部梵净山。最新种群数量统计显示,黔金丝猴约有800只, 少于川金丝猴(约25 000只)和滇金丝猴(约2000只),但多于怒江金丝猴(约300只)和越南金丝猴(约200只)[1,3]。
1 黔金丝猴的生境和食性
位于贵州省西北部的梵净山是黔金丝猴唯一分布区。梵净山于1986年成为国家级自然保护区。该区覆盖着较大面积原生性较强的森林植被, 从上至下呈现3个垂直带谱:海拔2200 m以上主要为寒温性针阔混交林和灌丛,海拔1300~2200 m主要是常绿、落叶阔叶混交林和针阔混交林, 海拔1300 m以下主要是常绿阔叶林和低山针叶林。黔金丝猴主要栖息于2000 m以下的常绿、落叶阔叶混交林和低山常绿阔叶林,位于27°49′50″~28°01′30″N,108°45′55″~108°48′30″E间。其中最适宜黔金丝猴生存的区域位于梵净山北部,即保护区的核心区,面积约204.53 km2的狭小范围,占保护区总面积47.27%;而保护区南部不太适宜黔金丝猴生存,面积约229.00 km2,占保护区面积52.73%[2-5]。
对黔金丝猴食性的研究表明,黔金丝猴食物种类较多,随季节不同而变化。春季主要食用叶和花,夏秋季以叶兼成熟或未成熟果实为主,冬季则以树芽、树皮、花苞等为食。此外,落地果实(如四照花果实)、昆虫等无脊椎动物都是采食对象[5-8]。与川金丝猴和滇金丝猴不同,近年观测未发现黔金丝猴采食地衣,可能与栖息地环境有关。黔金丝猴主要生活在700~2000 m间常绿、落叶阔叶混交林中,主要食物常年供应相对充足,地衣类植物分布很少[6]。
2 黔金丝猴主要行为特征及群体结构
黔金丝猴是白天活动的灵长类,并有日常活动中的垂直迁移现象,即选择低海拔夜宿和到高海拔采食的日活动模式。可能是获得食物和躲避天敌的一种权衡策略。另外黔金丝猴虽是一种昼行性灵长类,但也会在黄昏或夜晚活动。Tan等[9]用红外相机陷阱捕捉到黔金丝猴夜间活动情况,可能是一种适应性行为,通过增加活动时间来获得更多的食物资源和能力供给。
黔金丝猴的运动方式与其他几种金丝猴运动方式相同,主要是“半臂摆荡”(Semi-brachiation)型,介于“四足行走型”和“臂摆荡型”间。而其他灵长类的运动方式有“垂直攀爬型”如蜂猴(Nycticebuscoucang),“四足行走型”如猕猴(Macacamulatta), “臂摆荡型”如长臂猿科(Hylobatidae)动物。黔金丝猴的半臂摆荡式使它们既能适应树栖生活,又能适应地面生活[2,10-11]。
黔金丝猴行为学可以通过PAE(Posture-act-environment) 编码系统来进行分类和系统编码,构建PAE 行为谱来研究。崔多英等依此观察记录了野外和圈养黔金丝猴的14 类行为特征,包括16 种姿势、107 种动作及163 种行为。结果表明,黔金丝猴行为与川金丝猴和滇金丝猴行为有较多共同点,与猕猴类的行为区别较大。同时发现黔金丝猴在个体行为和社会行为方面,与川金丝猴和滇金丝猴相比也有不同点,可能与三者生活环境差异有关[12-15]。
关于黔金丝猴的群体结构,曾认为黔金丝猴群体是多雄配多雌的社会结构[5],目前认为黔金丝猴是以一成年雄猴和多个雌猴(One-male unit,OMU)的小家庭群体作为最基本单元(group),一般由5~10只组成;数量不等的一雄多雌社会单元构成一个更大群体(Bang, 帮);但也可在帮的周边观察到2~5个个体的全雄群。这种雄性替代的母系群是川、滇和黔3 种金丝猴社群结构的共同特征[ 5-6,11-12]。
在野外对黔金丝猴迁移和聚群行为观察过程中发现一个有趣的规律性现象,即黔金丝猴每年有2 次形成100~400只甚至更多个体的超级聚合群(large aggregation)。其生态适应机制尚有待深入研究[2,5], 但这种聚群活动明显与黔金丝猴生育季节性相关。
这种较大聚合群一次出现在春季4—5月,正是成年雌猴集中产仔时期(主要在3月底到4月底);另一次在秋季9—10 月,是成年雌猴受孕的有效交配时期。雌黔金丝猴的性成熟年龄为5~7岁,首次生育年龄为8~9岁,川金丝猴和滇金丝猴为5~6岁;如果幼猴存活至6个月大母猴的生育间隔周期为3年左右;川金丝猴和滇金丝猴只需要2年[11,16]。
3 黔金丝猴分子遗传学研究
分子遗传学技术是研究种系发育的有力工具。早期利用DNA聚合酶链式反应(PCR) 扩增黔金丝猴线粒体进化中较保守的基因如细胞色素b基因或变异较大的DNA控制区就已表明,黔金丝猴、滇金丝猴和川金丝猴隶属同一个属[17-18]。Li等[18]利用线粒体DNA细胞色素b基因402 bp片段和12SrRNA 387 bp片段分析3只黔金丝猴、9只滇金丝猴和7只川金丝猴的皮肤、肌肉、毛发和粪便样品,显示4种金丝猴与其他叶猴存在显著差异,将金丝猴立为一个独立属是合理的;越南金丝猴的种系关系不明,但不适合单列为一个亚属;文中数据支持将黔金丝猴作为一个独立种而非川金丝猴的一个亚种, 这是对形态学研究的一个重要支撑和补充[18-19]。最近Yang等[3]的系统进化分析和分子多态分析显示3个物种间存在清晰和明显的差异,各自为独立种。另外分子进化研究显示,黔金丝猴的共同祖先生活于大约36万年前,比川金丝猴(大约91万年前)和滇金丝猴(133万年前)近很多年[3]。
为了研究黔金丝猴的遗传多样性,Pan等[20]对野外黔金丝猴128只个体扩增线粒体DNA控制区可变区I 400 bp DNA片段,在400个阳性PCR中,发现无变异,仅存在一个单元型。但Yang等用线粒体DNA控制区可变区I 650 bp片段对141只黔金丝猴粪便样品分析发现,存在5个单元型,不过多态位点最多只有25个,相比于川、滇金丝猴,仍具有极低的单倍型遗传多样性[3]。
核基因组位点具有更多选择性,但受限于技术条件而进展较慢。从近缘种借用多态性标记适合于濒危物种;采用粪便、毛发等非损伤性取样方法的进步能减少对濒危物种的伤害,保证研究中材料来源;将二者合理结合既能不伤害物种又能进行遗传多样性分析,是黔金丝猴分子遗传学研究的一个重要方向。如Liu等借用人源的72个微卫星标记扩增滇金丝猴粪便基因组样品,13个标记获得成功,呈现适当多态性,有效分析了群体遗传结构和生殖策略[21]。我们最近采用非损伤性取样法,对粪便基因组DNA提取方法进行改进,从黔金丝猴粪便中成功提取总基因组DNA以及线粒体DNA,扩增得到特异性片段,并在分子生物学水平对黔金丝猴线粒体基因序列进行了分析。这不仅为黔金丝猴保护和繁育提供参考资料,也为非损伤性取样在珍稀濒危野生动物研究中应用拓宽思路[22]。
以上对黔金丝猴种群遗传多样性和群体结构研究表明,由于居住地丧失和片段化已造成种群隔离,使黔金丝猴成为只有一个连续群体的物种,具有极低遗传多样性。相比之下,川金丝猴在所有金丝猴中分布最广,滇金丝猴在云南省和西藏自治区西南部有大约11个群体。因此,在川、黔、滇3种金丝猴中,黔金丝猴群体增长缓慢,种群恢复潜力最低,该物种灭绝风险更高[3,16]。由于遗传多样性低和种群数量少,黔金丝猴的进一步研究需要在非损伤性取样基础上,进行较大规模的线粒体全基因组分析、核分子标记开发或核基因组研究。
4 黔金丝猴基因组学研究
黔金丝猴的线粒体基因组全序列已测定,在NCBI数据库中有两个全序列(JN540032.1 & JQ821836.1),约为16.5 kb[23-24]。序列比对发现两个黔金丝猴线粒体基因组序列相似度为99%;和人的相似度达81%,并具有与人线粒体基本一致的结构特点:同样含13个蛋白质编码基因、22个tRNA基因和2个rRNA基因;除ND6 亚基基因和8个tRNA基因位于轻链上,其他基因都位于重链上[23]。另外,两个线粒体全基因组中碱基组成和轻重链的化学特点虽有差异,仍十分相似。其碱基组成和轻链、重链分子质量如表1所示。
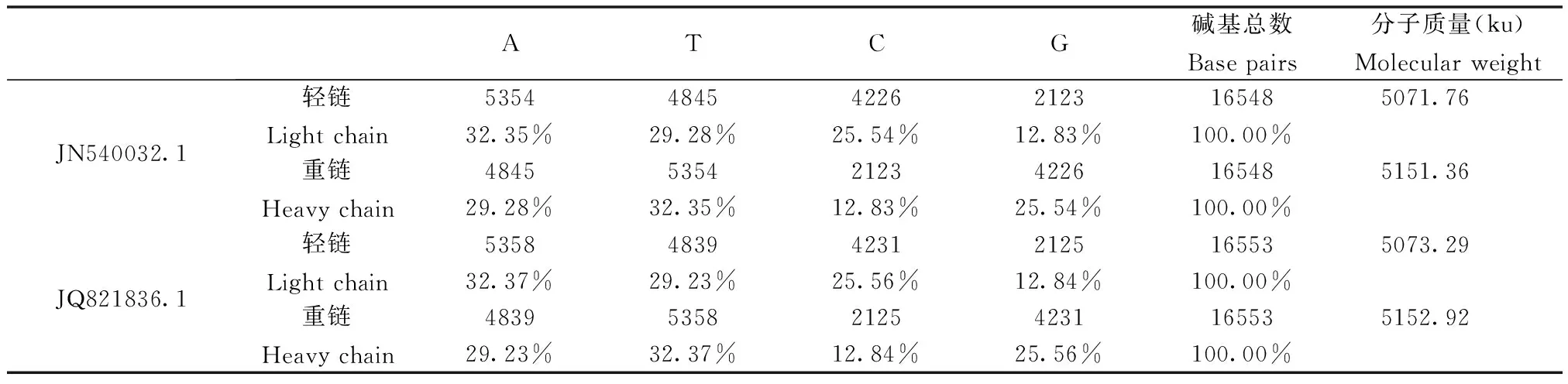
表1 黔金丝猴线粒体碱基组成分析
Liedigk等利用线粒体基因组序列和12个核基因组位点比较分析了3种叶猴(langur)和10种怪鼻猴(odd-nosed monkeys)(包括黔金丝猴)的系统发生[24]。依据线粒体数据(表2)发现仰面猴属(金丝猴属)与其他怪鼻猴的分异时间约为728万年前。对金丝猴属而言,首先是越南金丝猴与其他金丝猴分离,分异时间为237万年前。随后是可归为一支的黔金丝猴和川金丝猴(川-黔金丝猴姊妹群),与另一支的滇金丝猴和怒江金丝猴(滇-怒金丝猴姊妹群)分异于199万年前。黔金丝猴和川金丝猴分异时间是175万年前,滇金丝猴两个单倍型和怒江金丝猴的分异时间在24~60万年前。依据12个核基因座数据表明,首先可能是姊妹群之一的川-黔金丝猴姊妹群与其他金丝猴(滇、怒江、越南金丝猴)分异于169万年前,然后黔金丝猴和川金丝猴分异时间于131万年前。越南金丝猴与滇-怒金丝猴姊妹群分异约在73万年前,随后是滇金丝猴和怒江金丝猴分异于22~40万年前(表2)。以上线粒体数据和核基因组数据既呈现出大致相似的进化趋势,又存在着相当显著的差异。比如:1)黔金丝猴和川金丝猴形成进化一支(川-黔金丝猴姊妹群),而滇金丝猴和怒江金丝猴归为一支(滇-怒金丝猴姊妹群),由线粒体基因组与有限的基因座的分析结果是吻合的; 2)越南金丝猴的归属在两种分析中差异很大,导致越南金丝猴在两种分析中处于不同进化位置;3)由线粒体数据估计的分异时间远早于核基因数据,如黔金丝猴和川金丝猴分异时间分别为175万年(线粒体数据)和131万年前(核基因组数据)。
我们利用NCBI数据库中已发表的几种金丝猴所有线粒体基因组序列进行比较分析发现,两种黔金丝猴的线粒体基因组差异最小;黔金丝猴与川金丝猴最接近;滇金丝猴与怒江金丝猴或缅甸金丝猴较近;越南金丝猴与以上4种金丝猴的亲缘关系较远。这与上文基于线粒体分析的结果一致。另外发现一个滇金丝猴数据与怒江金丝猴的更接近。这显示滇金丝猴和怒江(缅甸)金丝猴间更为相近以及两种金丝猴之间存在的基因交流。
除了开展线粒体全基因组研究,黔金丝猴核基因组全序列也已被测定。最近Zhou等对4种金丝猴的38个个体作了全基因组测序,其中包括4只黔金丝猴[25];Yu等也对5种金丝猴进行全基因组测序,并做了和Liedigk 等类似的聚类分析[26]。邻接树法分析表明黔金丝猴和川金丝猴亲缘较近,形成了川-黔姊妹群;滇金丝猴和怒江(缅甸)金丝猴较近,形成了滇-怒姊妹群。与Liedigk等[24]的核基因组位点结果基本一致,与我们图1中线粒体的结果大体一致,只是推测的分异时间有差别(表2)。
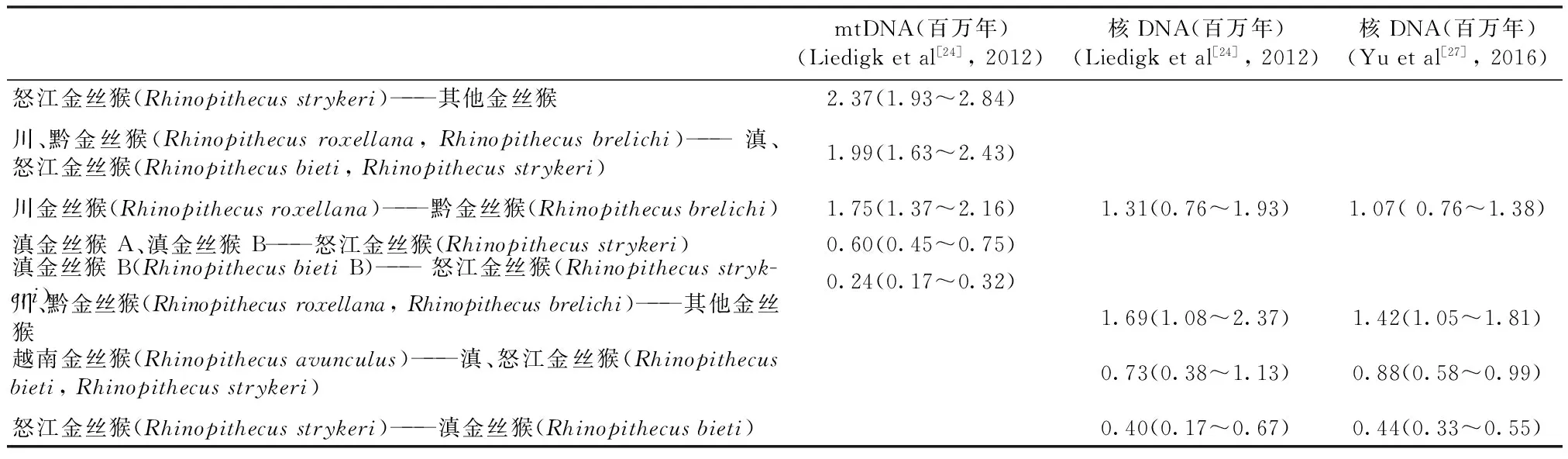
表2 金丝猴各个种群间的分异时间

图1 5种金丝猴(11个个体)的线粒体基因组聚类分析
Fig 1 Cluster analysis of mitochondrial genomes about fiveRhinopithecus(11 individuals)
基于NCBI的线粒体全基因组序列分析构建了如上系统发育树,其中共包括11条序列,黔金丝猴2个,滇金丝猴3个,怒江(缅甸)金丝猴1个,川金丝猴3个,越南金丝猴2个;序列号显示在图1括号中,越南金丝猴进化上比较古老,其次为黔金丝猴,川金丝猴等;一条滇金丝猴的线粒体序列和怒江金丝猴的相似度最高
综合以上数据,我们构建了黔金丝猴与其他金丝猴的种系发育的两种可能模型(图2),与王晓萍的分析部分一致[26]。线粒体数据表明越南金丝猴在较早时期便与其他金丝猴分异,是较为古老的种,其次是川-黔金丝猴姊妹群与怒-滇金丝猴姊妹群的分异(图2-A)。核基因组数据除了对于越南金丝猴进化地位的认知与线粒体数据截然不同外,对黔金丝猴与其他3种金丝猴的种系发育关系认知基本一致。显示川金丝猴和黔金丝猴亲缘较近,而滇金丝猴和怒江金丝猴较近;但越南金丝猴居于川-黔金丝猴姊妹群和滇-怒金丝猴姊妹群间(图2-B)。由于线粒体的母系遗传特性,线粒体在不同种群间保持一定独立性,其数据可能揭示曾经发生的分异事实;而核基因组数据中越南金丝猴地位的变化可能反映了怒江(缅甸)甚至滇金丝猴与越南金丝猴发生了一定程度的种群间杂交导致基因交流,最后使其序列同源性相近因而聚类分析靠近的现象。如果要得到更科学的结论,除了要适当增加样本量,并充分考虑种群间存在基因交流情况外,还要开发更精确的估算方法,解决线粒体数据和核基因组数据差异较大(包括估算分异年代)的问题。

图2 基于线粒体和核基因组数据构建的仰鼻猴属系统发育关系
A:基于线粒体序列构建的仰鼻猴属系统发育关系,显示川金丝猴和黔金丝猴亲缘较近(川-黔金丝猴姊妹群),而滇金丝猴和怒江金丝猴较近(滇-怒金丝猴姊妹群);而越南金丝猴进化上来说相对较为古老。B:基于核基因组序列,同样显示川金丝猴和黔金丝猴亲缘较近,而滇金丝猴和怒江金丝猴较近;但是越南金丝猴居于川-黔金丝猴姊妹群和滇-怒金丝猴姊妹群之间,黔金丝猴从进化上来说相对较为古老
由于地质事件、气候变化、地理隔离等原因造成金丝猴不同地理区域分布,使不同金丝猴种成为研究物种演化、环境适应的良好材料, 也成为研究基因功能、生物医学如肿瘤等研究的动物。因为在肿瘤发展过程中,会形成低氧微环境,从而诱导或稳定一些低氧诱导因子;而高原生物在长期进化中适应高原低氧低压环境,激活了与肿瘤中部分相似的信号通路。Yu等比较了低纬度生活的黔金丝猴、越南金丝猴(海拔2000 m以下)和高纬度(海拔3000 m以上)生活的其他3种金丝猴,发现高纬度金丝猴中,在6个和肺功能、DNA修复、血管再生的基因中,有8个共有的碱基替代;功能实验表明CDT1的变异(Ala537Val)使细胞抗紫外线辐射能力较强;RNASE4的变异体(Asn89Lys和Thr128Ile)增强了体外实验中内皮管的形成[27]。表明充分利用这一资源,可开展更多医学生物学研究。
5 结语
威胁黔金丝猴生存的因素主要包括遗传多样性、环境因素、适合度等。分子遗传学研究已揭示黔金丝猴具有极低遗传多样性,群体小及个体数量少是一个重要原因。其他研究也显示,黔金丝猴在食性、生境分布上比其他金丝猴更狭窄,在生育周期上比其他种更长。加之数量少造成的近亲交配会让黔金丝猴进一步丧失遗传多样性,导致生存力日渐衰退,适合度降低。环境因素包括森林火灾等自然灾害、人类活动如过度放牧、森林资源采集等。2013年建成的环梵净山公路,会使游人数量增加,加剧保护区的岛屿效应和生境破碎化。这些环境因素的变化目前虽尚未造成严重的影响,但必然是以黔金丝猴为旗舰物种的森林生态系统的不利因素,需给予高度关注。
遗传多样性贫乏和狭小地理分布现状,对黔金丝猴生存和发展极为不利,但这并非意味着将来该物种必然灭绝。因此,加强对梵净山黔金丝猴的研究与保护极为必要。我们建议:1)采用遗传管理的方法进行拯救,并对人工圈养的黔金丝猴,分析遗传多样性情况,在繁殖中优先考虑有差异或差异大个体的基因,以最大限度维持圈养群体遗传多样性;2)各自然保护区、各研究金丝猴团队彼此间应加强合作,共建网络平台,共享信息;3)鉴于怒江(缅甸)金丝猴的跨区域分布,加强与东盟成员国尤其是缅甸和越南合作,共同保护金丝猴栖息环境,展开国际合作研究;4)从技术层面,应加强基因组学方面研究,尤其是利用少量细胞高通量测序技术,获得更多个体遗传资料, 了解并保护黔金丝猴及其他金丝猴的遗传多样性,避免金丝猴不同种群部分或全部灭绝的命运。
致谢:感谢云南大学于黎教授阅读该文并提出的宝贵意见。
[1]汪 松. 中国濒危动物红皮书: 哺乳类[M]. 北京: 科学出版社, 1998: 62-64.
[2]崔多英, 牛克锋, TAN C L, 等. 分布在中国贵州梵净山的黔金丝猴[J]. 动物学杂志, 2014, 49(5):792.
[3]YANG M, YANG Y, CUI D, et al. Population genetic structure of Guizhou snub-nosed monkeys(Rhinopithecusbrelichi) as inferred from mitochondrial control region sequences, and comparison withR.roxellanaandR.bieti[J]. American Journal of Physical Anthropology, 2012, 147(1): 1-10.
[4]杨海龙, 李迪强, 成 钢,等. 贵州梵净山黔金丝猴生境适宜性评价[J]. 安徽农业科学, 2013, 41(8):3403-3406.
[5]杨业勤. 梵净山研究:黔金丝猴的野外生态[M]. 贵阳:贵州科技出版社, 2002.
[6]吴 钢, 王宏昌, 付海威,等. 樊净山自然保护区黔金丝猴生境选择的研究[J]. 林业研究(英文版), 2004, 15(3):197-202.
[7]聂帅国, 向左甫, 李 明,等. 黔金丝猴食性及社会结构的初步研究[J]. 兽类学报, 2009, 29(3):326-331.
[8]牛克锋, TAN C L, 崔多英,等. 黔金丝猴(Rhinopithecusbrelichi)雪季地表食物利用[J]. 野生动物学报, 2014, 35(1):31-37.
[9]TAN C L, YANG Y, NIU K. Into the night: camera traps reveal nocturnal activity in a presumptive diurnal primate,Rhinopithecusbrelichi[J]. Primates, 2013, 54(1):1-6.
[10]叶智彰. 金丝猴解剖[M]. 昆明:云南科技出版社, 1987.
[11]BLEISCH W, CHENG A S, REN X D, et al. Preliminary results from a field study of wild Guizhou snub-nosed monkeys(Rhinopithecusbrelichi)[J]. Folia Primatologica, 1993, 60(1-2):72-82.
[12]李 勇, 任宝平, 李艳红,等. 滇金丝猴的行为谱及PAE编码系统[J]. 四川动物, 2013, 32(5):641-650.
[13]田军东, 王振龙, 路纪琪,等. 基于PAE编码系统的太行山猕猴行为谱[J]. 兽类学报, 2011, 31(2):125-140.
[14]梁 冰, 戚汉君, 张树义,等. 笼养川金丝猴不同年龄阶段的发育特征[J]. 动物学报, 2001, 47(4):381-387.
[15]崔多英, 牛克锋, TAN C L, 等. 黔金丝猴的行为谱及PAE编码系统[J]. 四川动物, 2014(6):815-828.
[16]YANG M, SUN D Y, ZINNER D, et al. Reproductive parameters in Guizhou snub-nosed monkeys(Rhinopithecusbrelichi)[J]. American Journal of Primatology, 2009, 71(3):266-270.
[17]ZHANG Y P, RYDER O A. Mitochondrial cytochrome b gene sequences of old world monkeys: with special reference on evolution of Asian colobines[J]. Primates, 1998, 39(1):39-49.
[18]LI M, WEI F, HUANG C, et al. Phylogeny of snub-nosed monkeys inferred from mitochondrial DNA, cytochrome B, and 12S rRNA sequences[J]. International Journal of Primatology, 2004, 25(4):861-873.
[19]JABLONSKI N G, PENG Y Z. The phylogenetic relationships and classification of the doucs and snub-nosed langurs of China and Vietnam[J]. Folia Primatologica, 1993, 60(1-2):36-55.
[20]PAN H J, SHI F L, CHANG Z F, et al. Mitochondrial DNA variation analysis suggests extreme low genetic diversity in Guizhou snub-nosed monkeys(Rhinopithecusbrelichi)[J]. Chinese Science Bulletin, 2011, 56(24):2541-2544.
[21]LIU Z J, REN B P, HAO Y L, et al. Identification of 13 human microsatellite markers via cross-species amplification of fecal samples fromRhinopithecusbieti[J]. International Journal of Primatology, 2008, 29(1):265-272.
[22]ZHANG Z M, XIAO D M, QIAN G, et al. An efficient protocol for genomic DNA extraction from the endangered species[J]. Biotechnology & Biotechnological Equipment, 2015, 29(3):530-535.
[23]SHI F, WANG B, LIU Z, et al. Mitochondrial genome of the Guizhou snub-nosed monkey(Rhinopithecusbrelichi) [J]. Mitochondrial DNA, 2011, 22(5-6):191-193.
[24]LIEDIGK R, YANG M, JABLONSKI N G, et al. Evolutionary history of the odd-nosed monkeys and the phylogenetic position of the newly described Myanmar snub-nosed monkeyRhinopithecusstrykeri[J]. PloS One, 2012, 7(5):e37418.
[25]ZHOU X, MENG X, LIU Z, et al. Population genomics reveals low genetic diversity and adaptation to hypoxia in snub-nosed monkeys[J]. Molecular Biology & Evolution, 2016, 33(10):2670-2681.
[26]王晓萍, 张亚平, 于 黎. 疣猴亚科(Colobinae)系统发育研究概述[J]. 科学通报, 2013, 58(16): 1465-1472.
[27]YU L, WANG G D, RUAN J, et al. Genomic analysis of snub-nosed monkeys(Rhinopithecus) identifies genes and processes related to high-altitude adaptation[J]. Nature Genetics, 2016, 48(8):947-952.
猜你喜欢
野生动物学报(2022年4期)2022-11-07
党政干部论坛(2022年3期)2022-04-20
中学生数理化·高一版(2021年1期)2021-03-19
小哥白尼(野生动物)(2020年4期)2020-07-27
传播与制作(2019年10期)2019-12-02
中华诗词(2019年7期)2019-11-25
学生天地(2018年36期)2019-01-28
小学生学习指导(低年级)(2017年9期)2017-08-07
环境保护与循环经济(2017年9期)2017-03-16
河北地质(2016年4期)2016-03-20
