
草鱼日粮中添加饲料桑粉对其生长性能及非特异性免疫的影响
2018-04-16 05:45刘家星陈霞曹英伟李良玉张小丽魏文燕杨马唐洪陈健
水产养殖 2018年4期
刘家星,陈霞,曹英伟,李良玉,张小丽,魏文燕,杨马,唐洪,陈健
(成都市农林科学院水产研究所,四川 成都 611130)
随着粮食耕地的减少以及农作劳动力的减少,在满足人类消费的情况下,剩余用来加工成畜禽、水生动物饲料的粮食越来越短缺,人和动物的粮食竞争矛盾日益突出。因此,为有效缓解饲料原料紧缺的情况,开发和寻找新的、优质的饲料原料迫在眉睫[1]。
饲料桑[2]是我国科研人员通过努力培育出的具有超大潜力的桑树新品系,具有较强的抗逆性,很强的耐贫瘠和耐严寒能力[3]。生长非常迅速,在条件良好的情况下当年桑苗可达2.5~3.0 m,单株鲜重达1.5 kg,干重0.3 kg。鲜叶重每1hm2每年可达40 t,干物质10 t,可饲用部分干物质合计13 t[2]。饲料桑的营养价值丰富,含有大量的粗蛋白,其平均含氮量高达3.574%[3],有研究表明饲料桑叶的总干物质中蛋白质的比例为23%,氨基酸种类有18种之多。此外,饲料桑中还含有大量的纤维素、维生素和矿物质元素等营养成分,其营养组成和比例适宜动物的生长发育需要[4-6]。
目前饲料桑已在肉鸡、蛋鸡、猪、肉牛、奶牛、肉羊等动物上得到了广泛应用。一方面可以提高动物的生长性能,动物产品的质量[7-8];另一方面还可以提高动物的免疫能力,改善动物的生长状态[7]。桑叶属天然植物,含有黄酮类、多糖类、植物甾醇等多种天然活性物质,能提高动物免疫力,有利于动物抗病和保健。英国最新研究成果表明桑叶中还含有一种叫做“达菲”的物质,对防治H5N1型禽流感有效[9]。关于饲料桑在淡水鱼养殖的应用中,目前仅见2002年中国工程院三峡工程评估项目组将饲料桑应用于库区养殖鱼中,并取得了不错的效果[10]。但是,目前相关资料的研究相对较少,对于其是否能够提高鱼体的免疫能力相关的研究更是匮乏。因此,本试验选取传统养殖品种草鱼(Ctenopharyngodon idellus)作为试验对象,探究在草鱼日粮中添加饲料桑对草鱼生长性能及非特异性免疫的影响,以期能够为饲料桑在淡水鱼饲料中的开发应用奠定基础。
1 材料和方法
1.1 材料及试剂
草鱼鱼种 1 200 尾,体质量(55±4.5)g;饲料桑原粉200 kg,饲料桑鲜叶若干;规格为直径115 cm、高45 cm的玻钢纤维盆24个;血清总补体测试试剂盒、总超氧化物岐化酶测试试剂盒购自南京建成生物工程研究所;其他试剂购自上海生工生物科技有限公司。所有试剂均为分析纯。
1.2 试验方法
1.2.1 试验动物分组 试验用玻钢纤维盆使用0.1%高锰酸钾盛满浸泡2 h,再反复刷洗消毒后方可用于试验。试验用草鱼购买回来后以3%的食盐水浸泡鱼体15 min,然后在池塘暂养15 d后进行试验分组。试验分为6个组,其中添加饲料桑原粉共5组,添加比例为0%、3%、6%、9%、12%,另单独1组投喂饲料桑鲜叶。随机挑选规格一致、体质健壮、体质量50 g左右的草鱼,每组饲养草鱼100尾,设两个平行。
1.2.2 饲料制备及投喂 试验用饲料配方营养需求参照草鱼NRC 1993,各组添加饲料桑分别为0%(A0)、3%(A1)、6%(A2)、9%(A3)、12%(A4),配制成5种等氮(蛋白水平为 30.8%~30.9%)、等能(能值为13 kJ/g)的试验饲料。各原料经粉碎机粉碎过60目筛,按配方准确称重,逐级放大混合,加水拌匀,用饲料颗粒机制成粒径1.0 mm颗粒饲料,于40~45℃下烘干至水分含量10%以下,保存在-20℃冰箱中备用。饲料桑鲜叶组(A5),在饲喂基础日粮基础上每日采摘饲料桑鲜叶投喂。试验从草鱼分组后第 5 天开始投喂,每日早、中、晚(8:00,12:00,18:00)投喂3次,严格按投喂“四定”(定时、定点、定量、定质)原则进行。日投饵率5%左右,以20 min内食完为宜。玻钢纤维盆施以微流水,水体交换量0.1 m3/h。每日巡视做好防逃防敌害工作,对投喂、吃食等情况做好记录。试验饲料组成及营养成分见表1和表2。
1.2.3 生长性能测定 分别于试验开始前记录各组的初始体质量,饲养90 d后记录末体质量,记录投饲量。每次称重前鱼体饥饿24 h。每组随机选取4尾鱼,测定其体长、体质量,用于计算鱼体增重率、饲料系数、特定生长率。计算公式如下:
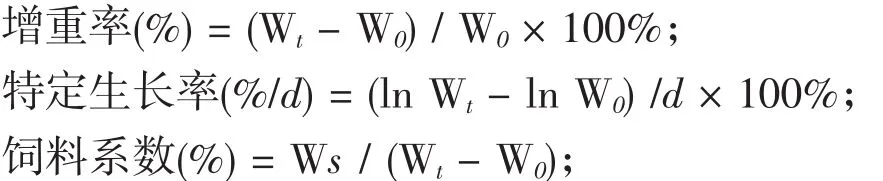
式中,Wt表示试验末均重,W0表示试验初均重,Ws表示试验均投饲量,d表示试验天数。
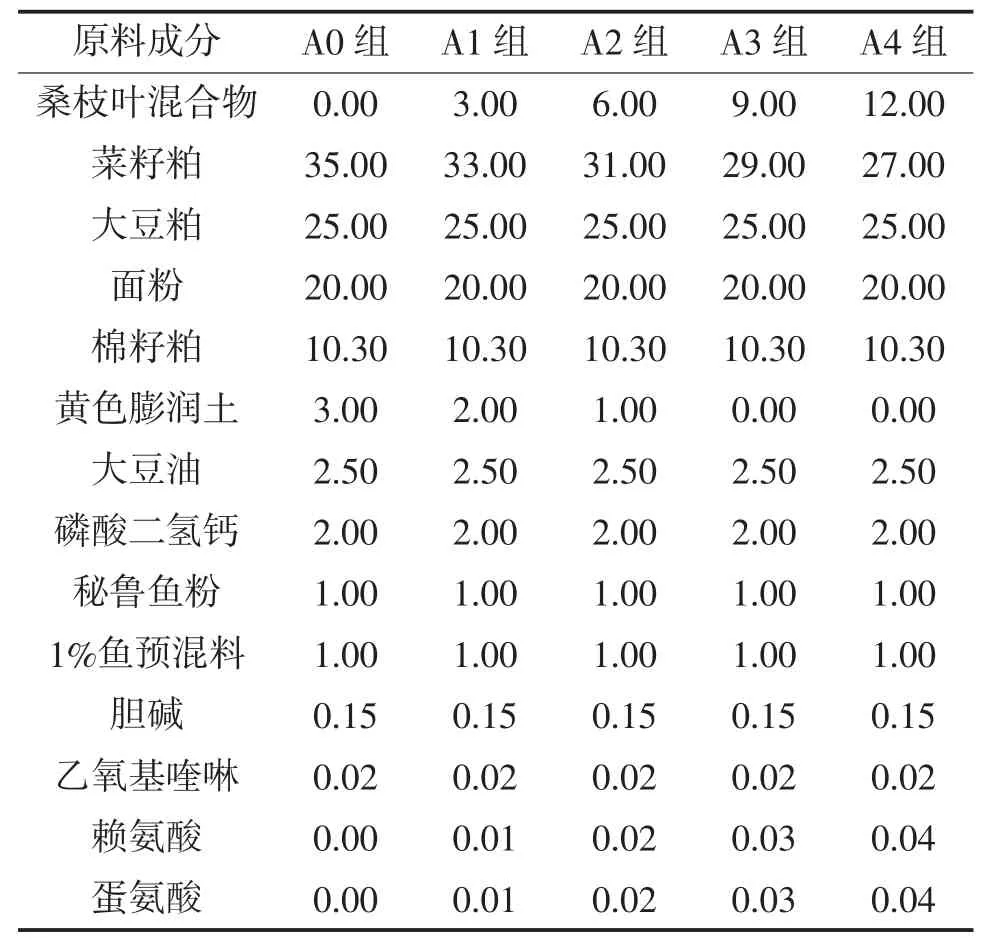
表1试验分组饲料配方表 %
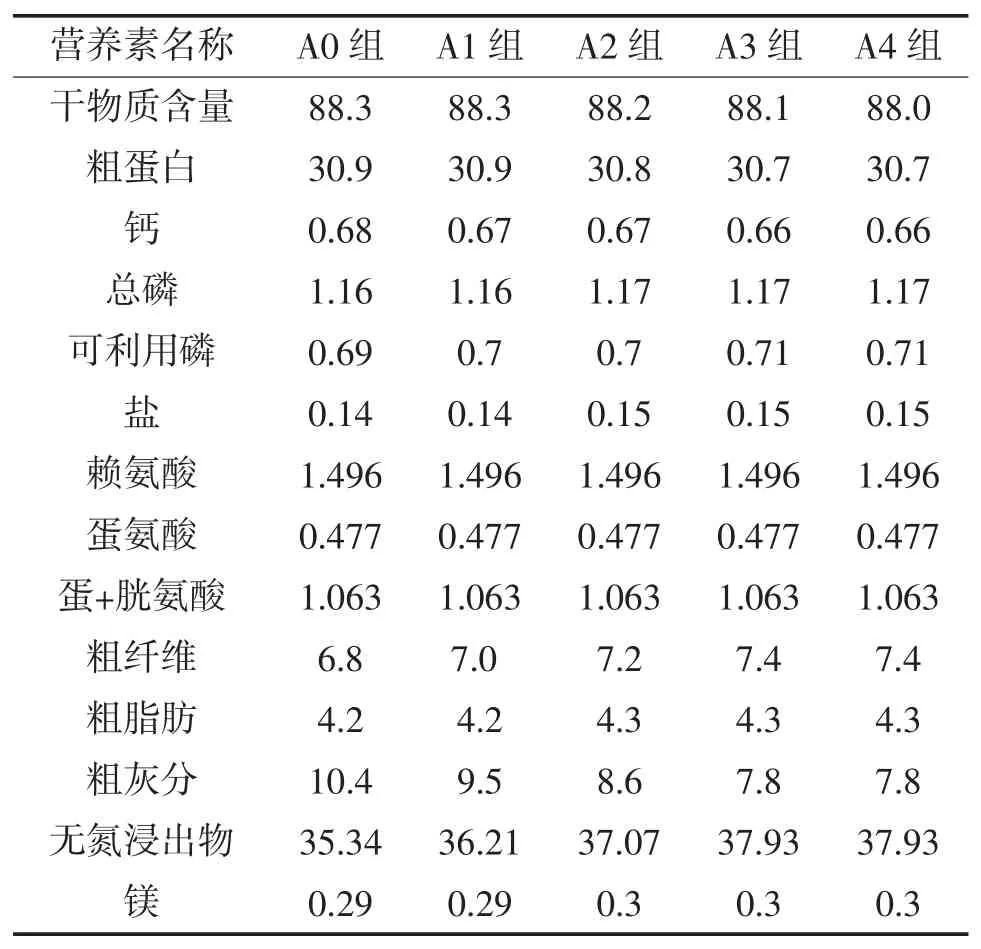
表2草鱼饲料营养成分表 %
1.2.4 非特异性免疫指标测定 分别于投喂饲料0、15、30、45、60、75、90 d 对草鱼非特异性免疫指标进行测定(总超氧化物歧化酶、溶菌酶活性、血清杀菌活性,血清补体C3),每组随机挑选4尾鱼进行尾静脉采血,收集血液置于1.5 mL的PE管中,于4℃静置12 h,4 000 r/min离心l0 min,取上层血清置于-80°C超低温冰箱保存备用。总超氧化物歧化酶、血清补体C3测定方法参照南京建成生物工程研究所试剂盒说明书进行;溶菌酶测定参照Swain P和Behera T等[11-12]文献方法进行;血清杀菌活性参照Sun Y等[13]文献方法进行。
1.2.5 数据统计 使用 SPSS 22.0软件进行统计学分析,对各组试验重复数据作方差同质性检测后进行单因素方差分析,使用LSD法进行事后多重比较,结果以“平均数±标准差(mean±SD)”表示。分别在0.01和0.05差异水平上进行检验,当P<0.01表示差异极具统计学意义,P<0.05表示差异具统计学意义,P>0.05表示差异不具统计学意义。
2 试验结果
2.1 草鱼生长性能
按照1.2.3种所述方法,用添加不同含量的饲料桑粉饲料投喂90 d,各组草鱼在试验结束后计算各组的增重率、特定生长率、饲料系数,结果见表3。从表中可知,添加量为6%的试验组(A2)和饲喂鲜叶组(A5)的草鱼其增重率极显著(P<0.01)高于其他试验组和对照组;添加量为3%的试验组(A1)特定生长率极显著(P<0.01)低于其他试验组而与对照组(A0)相比差异不具统计学意义(P>0.05);添加量为6%的试验组饲料系数极显著(P<0.01)低于对照组,且显著(P<0.05)低于添加量为3%、12%(A4)的试验组。

表3草鱼生长性能测定
2.2 非特异性免疫指标
2.2.1 总超氧化物歧化酶活力 投喂不同时间后采集各组草鱼血清后测得血清总超氧化物歧化酶活力变化情况见图1。在投喂后15 d开始,添加不同比例饲料桑的试验组以及投喂饲料桑鲜叶组(A5)血清的总超氧化物歧化酶活性呈上升趋势,除与对照组(A0)差异极具统计学意义(P<0.01)外,其余各组差异不显著(P>0.05);第30天,添加量为12%的试验组(A4)与其他组具统计学意义(P<0.05);到第60天,各个试验组均达到较高的酶活力,添加量为9%的试验组(A3)酶活力水平为整个试验周期最大值,达到(23.14±0.03)U/mL;随后各试验组酶活力开始下降,但在试验周期内与对照组仍保持极具统计学意义(P<0.01)。对照组在整个试验周期中酶活力水平维持在16.4~16.9 U/mL范围内。添加量为9%的试验组在整个试验周期中,能够持续较好地刺激草鱼鱼体的抗氧化系统。
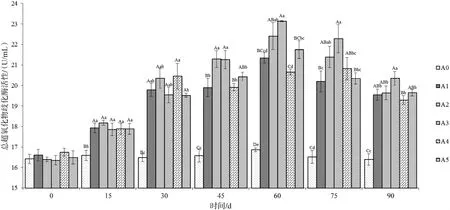
图1血清总超氧化物歧化酶(T-SOD)活力变化
2.2.2 血清溶菌酶活性 参考Swain P等[11-12]方法,在测定试验开始前使用冻干的鸡蛋白溶菌酶做标准曲线(酶活分别为 50 U、40 U、30 U、20 U、10 U、5 U、1 U)。溶菌水平以U/mL表示,0.001 U/mL表示一个单位U。投喂不同时间后采集各组草鱼血清后测得血清溶菌酶变化情况见图2。在投喂后15 d开始,添加不同比例饲料桑的试验组以及投喂饲料桑鲜叶组(A5)血清溶菌酶活性呈上升趋势,除与对照组(A0)和添加量为9%(A3)的试验组差异极具统计学意义(P<0.01)外,其余各组差异不具统计学意义(P>0.05);第30天,各试验组血清溶菌酶活性均有上升,差异显著性与15 d相似;第45天,添加量为3%的试验组酶活性上升较为缓慢,与其他试验组差异极具统计学意义(P<0.01);到第60天,添加量为3%和9%的试验组酶活性与其他试验组差异极具统计学意义(P<0.01);到第75天添加量为6%的试验组血清溶菌酶活性达到整个试验周期最大值,为(637±10)U/mL;随后各试验组血清溶菌酶活性开始下降,但添加量为3%以及鲜叶组下降的速率相对缓慢,与其他试验组仍差异极显著(P<0.01)。对照组在整个试验周期中酶活力水平维持在230~260 U/mL范围内。整个试验周期中,不同添加量的试验组均能较好地刺激鱼体溶菌酶系统的发生,以添加量为6%的试验组以及饲喂鲜叶的试验组能持续更长的时间。
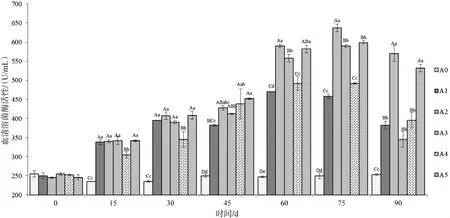
图2血清溶菌酶活性变化
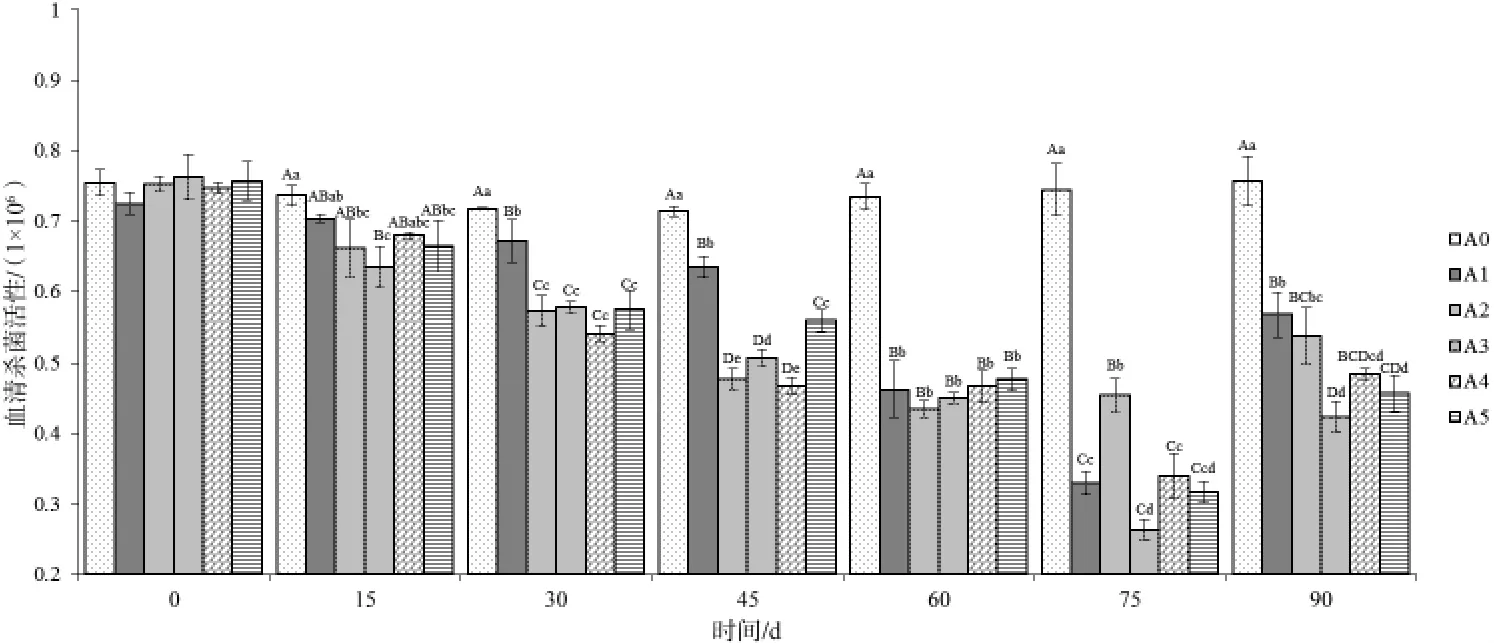
图3血清杀菌活性变化
2.2.3 血清杀菌活性 参考Sun Y等[13]方法,采用平板计数法反映各试验组不同时间点的血清杀菌活性,活菌数少的样本血清杀菌活性高。投喂不同时间后采集各组草鱼血清测得血清杀菌活性变化情况见图3。在投喂后15 d开始,添加不同比例饲料桑的试验组以及投喂饲料桑鲜叶组(A5)血清杀菌活性开始上升,除添加量为9%(A3)的试验组与其他试验组及对照组差异极具统计学意义(P<0.01)外,其余各组变化幅度较小;第30天,各试验组血清杀菌活性均有上升,但添加量为3%(A1)的试验组上升较慢,与其他试验组差异极具统计学意义(P < 0.01);第75天,添加量为9%的试验组(A3)血清杀菌活性达到整个试验周期最大值,平板计数活菌数仅为(0.263±0.015)×106CFU/mL;随后各试验组血清杀菌活性开始下降,但添加量为9%以及鲜叶组下降的速率相对缓慢,与其他试验组仍差异极具统计学意义(P<0.01)。对照组在整个试验周期中平板计数活菌数维持在 0.71×106~0.76×106CFU/mL。
2.2.4 血清补体C3 按照试剂盒说明书,建立标准曲线,根据测得吸光度值对应标准曲线即可得到血清补体C3含量。投喂不同时间后采集各组草鱼血清测得血清补体C3含量变化情况见图4。从结果可知,各试验组血清补体C3含量变化均呈现先升高后降低的趋势,与测定的其他指标变化趋势类似。添加量为9%的试验组(A3)血清补体C3含量在75 d达到整个试验周期最大值,为(48.91±1.21)mg/mL。对照组在整个试验周期中血清补体C3含量维持在12.3~12.7 mg/mL之间。
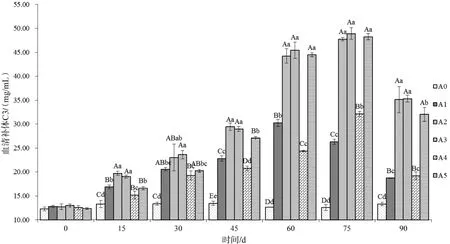
图4血清补体C3含量变化
3 讨论
3.1 添加饲料桑粉对生长性能的影响
在本试验中,在草鱼日粮中添加饲料桑粉或投喂饲料桑鲜叶均呈现较好的促生长作用。目前关于饲料桑在淡水渔业中的应用相关报道较少,仅在陆生动物中有相关研究。在草鱼日粮中添加饲料桑粉一方面可促进草鱼生长,降低饵料系数;另一方面,可减少粮食的投入降低人与动物之间的粮食竞争。同时提高饲料的利用率,减少饲料的投喂以及对环境的污染。
3.2 添加饲料桑粉对非特异性免疫的影响
血清超氧化物歧化酶(SOD)是重要的抗氧化酶之一,广泛存在于生物体内,它的主要功能是清除氧自由基,保护机体免受氧化伤害的一种关键酶。在调节机体氧化与抗氧化之间的平衡中扮演重要角色。在正常机体内氧自由基的形成和清除处于动态平衡,一旦平衡被打破,那么机体产生的氧自由基就会在体内堆积,造成疾病和衰老。此外血清超氧化物歧化酶还关乎生物体的免疫水平,它能够提高巨噬细胞的防御能力以及对整个机体的免疫功能都有一定的影响,它通常作为机体非特异性免疫的一个指标[14]。在本试验中,在草鱼日粮中添加饲料桑粉在一定程度上能刺激鱼体抗氧化系统的发生,通过对不同时间点草鱼血清SOD活性进行检测,结果发现不同添加量的试验组以及饲喂鲜叶的草鱼血清SOD活性呈现先升高后降低的趋势,且以添加量为9%的试验组最好,添加量为6%的试验组和饲喂鲜叶组次之。结果说明在草鱼日粮中添加饲料桑粉能够激发草鱼机体产生抗氧化酶,但相对于免疫增强剂来说,其激发程度相对弱一些,不会对机体产生过氧化损伤。
血清溶菌酶主要在肝脏总合成,广泛存在于鱼类血清、皮肤黏液、巨噬细胞和嗜中性粒细胞[15],是一种能水解致病菌中黏多糖的碱性酶,在防御病原入侵时起到溶解细菌的作用。在本试验中,对饲喂饲料桑粉后的草鱼血清溶菌酶含量进行检测,结果发现饲喂含饲料桑粉的饲料或饲料桑鲜叶后的草鱼血清溶菌酶含量都呈现先升高后降低的趋势,在投喂后的75 d各试验组溶菌酶含量水平较高。这表明饲料桑能够有效提高草鱼的非特异性免疫,刺激肝脏产生溶菌酶。整个试验周期中,添加量为6%的试验组产生的溶菌酶水平最好,添加量为9%的试验组和饲喂鲜叶组次之。
血清补体C3在鱼类上,可以作为抗体和吞噬细胞间连接的中介,是能够增强体液和细胞介导的特异性免疫的关键成分[16];血清杀菌活性反映了血清中杀菌成分抗菌力大小的指标,综合反映了多种血清非特异性免疫物质综合作用的抗菌能力。在本试验中,对这两种非特异性免疫指标检测结果发现,饲喂饲料桑粉和鲜叶均可提高鱼体的抗菌能力,有助于鱼体抵抗病原微生物的入侵,能够综合提高鱼体的抗病能力。
3.3 草鱼日粮中饲料桑粉添加量探讨
在整个试验周期中,在草鱼日粮中添加饲料桑粉含量为6%和9%的试验组能够较好地促进草鱼生长,提高草鱼的增重率,降低饵料系数。同时还可以刺激鱼体非特异性免疫,产生一系列体液因子,有助于提高草鱼的抗病能力。由此可见,饲料桑粉作为新的饲料原料添加在草鱼日粮中具有非常大的价值,根据本试验研究结果可知添加量为9%可作为实际生产中的添加含量。
参考文献:
[1]李有业,耿凤琴.桑叶桑枝在鸡饲料中的应用效果[J].当代畜牧,2009(1):30-31.
[2]孙双印,侯向阳,卢欣石.二十一世纪的绿色神奇功能饲料植物——饲料桑[J].中国奶牛,2007(2):53-54.
[3]孙双印,侯向阳,卢欣石.饲料桑特性研究与加工利用分析[J].草原与草坪,2008(1):63-69.
[4]刘红,叶志毅.桑叶为畜禽饲料的利用价值评价[J].饲料研究,2001(9):13-14.
[5]苏海涯,吴跃明,刘建新.桑叶中的营养物质和生物活性物质[J].饲料研究,2001(9):1-3.
[6]黄自然,杨军,吕雪娟.桑树作为动物饲料的应用价值与研究进展[J].蚕业科学,2006,32(3):377-385.
[7]杨静.饲料桑粉的营养价值评定及在生长育肥猪日粮中的应用研究[D].河北:河北农业大学,2014.
[8]李钢.新的功能性饲料原料——饲用桑叶粉在我国问世[J].畜牧市场,2006(11):31-31.
[9]蔡晶晶,卢玉飞,李军,等.桑叶在饲料资源开发中的研究与利用[J].广东饲料,2011(6):31-33.
[10]许勤.利用饲料桑发展畜禽淡水鱼产业生态经济社会效益显著[J].林业经济,2014(2):128-128.
[11]Swain P,Dash S,Sahoo P K,et al.Non-specific immune parameters of brood Indian major carp Labeo rohita,and their seasonal variations[J].Fish&Shellfish Immunology,2007,22(1-2):38-43.
[12]Behera T,Swain P.Alginate-chitosan-PLGA composite microspheres induce both innate and adaptive immune response through parenteral immunization in fish[J].Fish&Shellfish Immunology,2013,35(3):785-91.
[13]Sun Y,Hu Y H,Liu C S,et al.Construction and comparative study of monovalent and multivalent DNA vaccines against Streptococcus iniae.[J].Fish&Shellfish Immunology,2012,33(6):1303-1310.
[14]艾春香,陈立侨,高露姣,等.Vc对河蟹血清和组织中超氧化物歧化酶及磷酸酶活性的影响[J].应用海洋学学报,2002,21(4):431-438.
[15]杨先乐.鱼类免疫学研究的进展[J].水产学报,1989,13(3):271-284.
[16]Sakai D K.Repertoire of complement in immunological defense mechanisms of fish[J].Annual Review of Fish Diseases,1992,2(2):223-247.
猜你喜欢
农业工程学报(2022年13期)2022-10-09
当代水产(2022年7期)2022-09-20
当代水产(2022年3期)2022-04-26
畜牧兽医学报(2022年4期)2022-04-24
广州化工(2022年20期)2022-01-01
齐鲁工业大学学报(2021年1期)2021-12-30
消费导刊(2020年35期)2021-01-28
中国茶叶(2020年7期)2020-07-31
当代水产(2019年9期)2019-10-08
广东第二课堂·小学(2018年9期)2018-10-24
