
5个地区长肋日月贝形态性状对主要经济性状的相关性和通径分析
2018-04-12 06:03邓正华陈明强李有宁吴开畅王雨
中国渔业质量与标准 2018年2期
邓正华,陈明强,李有宁,吴开畅,王雨
(中国水产科学研究院南海水产研究所, 农业部南海渔业资源开发利用重点实验室,广州510300;中国水产科学研究院南海水产研究所热带水产研究开发中心, 海南 三亚572018)
生物体表型性状不仅受基因型与环境的各自影响,也受它们互作的影响,表型性状是遗传育种与苗种繁育的重要指标,常被作为生物体进行良种选育及种质复壮的主要性状[1]。而在实际养殖生产中,重量通常直接影响着养殖产业的经济效益[2]。控制生物体形态性状和重量的基因间往往存在着遗传连锁,相比于重量,形态性状更便于测量[1]。在水产动物中,基于形态性状直观易测的优势,研究其对主要经济性状的决定和影响程度,并以形态性状制定选育指标,筛选出具有高产性能的选育群体,对开展人工选育具有重要意义。已有研究表明,针对不同种类水产动物,影响其主要经济性状的形态性状有所不同,如黄边糙鸟蛤(Trachycardiumflavum)[3]、日本镜蛤(Dosiniajaponica)[4]、背瘤丽蚌(Lamprotulaleai)[5]、日本沼虾(Macrobrachiumnipponensis)[6]和日本囊对虾(Marsupenaeusjaponicus)[7]等的研究中都有体现。通径分析早在20世纪20年代就由数量遗传学家Wright提出,通径分析可以确定参数间的相关性,并将参数间的相关系数拆分为直接影响和间接影响,从而通过通径分析建立最优回归方程[8]。利用形态性状与经济性状间的通径分析指导了许多海洋生物的育种工作[9-13]。针对香港巨牡蛎(Crassostreahongkongensis)[14]的研究发现,影响其湿重的主要性状在雌雄间有所不同;在对日本沼虾[15]的研究中发现,影响其湿重的主要性状在不同地理群体中有所不同。
长肋日月贝(Amusiumpleuronectes)隶属于软体动物门(Mollusca)、双壳纲(Bivalvia)、珍珠贝目(Pectinoida)、扇贝科(Pectinidae)、日月贝属(Amusium)[16]。主要分布于热带和亚热带区域,在中国主要分布于广东、广西和海南等沿海地区[17]。长肋日月贝壳薄、个体大、生长快、肉质肥满、闭壳肌发达、肉味鲜美[18],具有较大的食用价值和观赏价值,是中国、泰国、菲律宾和澳大利亚等国的重要经济贝类[17],也是一种极具开发潜力的物种。国内外就长肋日月贝生物学性状与资源时空分布[17]、营养成分分析[19]、环境因子对滤水率的影响[20]、苗种繁育[21-24]以及遗传结构分析[25]等进行了一定的研究,在其他方面研究较少。本研究以广西北海、海南儋州、海南三亚、海南陵水以及海南琼海收集的野生长肋日月贝为研究对象,将不同地区长肋日月贝形态性状对主要经济性状进行多元回归分析;获取不同地区影响其经济性状的主要形态性状,从而建立最优回归方程;对保留的各形态性状,分析其对经济性状的直接作用和间接作用,及其单独或两两共同对经济性状的决定程度,以期为人工选育目标性状时提供数据支持。
1 材料与方法
1.1 实验材料
实验用长肋日月贝采集于2016年7—10月间,其中92枚来自广西北海、100枚来自海南儋州、60枚来自海南三亚、100枚来自海南陵水和100枚来自海南琼海,均为野生2龄贝。
1.2 实验方法
对上述采集的野生长肋日月贝形态性状和主要经济性状进行测量,使用精确度为0.01 mm的数显游标卡尺测定壳长(x1/mm)、壳高(x2/mm)、壳宽(x3/mm)和绞合线长(x4/mm),使用精确度为0.01 g的电子称称量湿重(y1/g)、软体重(y2/g)、性腺重(y3/g)和闭壳肌重(y4/g),各性状示意图如图1。
1.3 数据分析
利用Excel 2016记录、整理各测量数据,采用SPSS 21.0 (IBM, USA)软件分析各性状间相关性,计算形态性状对各经济性状的通径分析和决定系数。分析5个地区形态性状对其经济性状的直接作用和间接作用,经逐步回归方法剔除偏回归系数不显著的性状,并根据被保留的性状计算复相关系数以及建立各形态性状与经济性状间的最优回归方程。采用Origin 9.1(Origin Lab, USA)软件对长肋日月贝5个地区基于形态性状进行聚类分析并作图。
其中,决定系数(di)计算公式为:di=Pi2,Pi为某形态性状对某经济性状的通径系数;多参数共同决定系数(dij)为:dij=2rijPiPj,rij为性状间的相关系数,Pi、Pj分别为单一参数对某一经济性状的通径系数;多元线性回归方程为:y=β0+β1x1+β2x2+ ...... +βkxk,x1、x2、......、xk为偏回归系数对应的自变量,β0为常数,β1、β2、......、βk为偏回归系数。
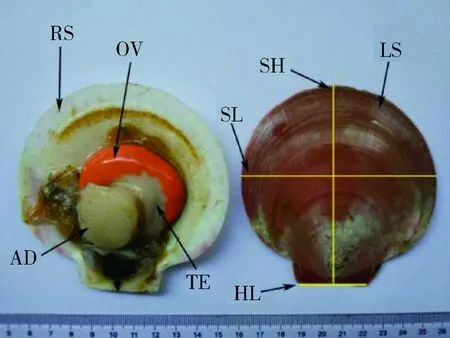
图1 长肋日月贝图解RS:右壳;LS:左壳;OV:卵巢;TE:精巢;AD:闭壳肌;SL:壳长;SH:壳高;HL:绞合线长。Fig.1 Illustration of A.pleuronectesRS: right shell; LS: left shell; OV: ovary;TE: testis; AD: adductor muscle; SL: shell length;SH: shell height; HL: hinge line length.
2 结果
2.1 不同地区长肋日月贝各性状参数估计值
2.2 性状间的简单相关分析
形态性状和经济性状概括如表2,实验所有性状间均表现为相关性极显著(P<0.01)。其中,广西北海、海南儋州、海南三亚、海南陵水和海南琼海地区与湿重相关系数最大的形态性状分别是壳长(0.903)、壳高(0.946)、壳长(0.988)、壳高(0.974)和壳高(0.986),与软体重相关系数最大的分别是壳宽(0.807)、壳高(0.930)、壳长(0.985)、壳高(0.960)和壳高(0.980),与性腺重相关系数最大的分别是壳长(0.478)、壳高(0.506)、壳高(0.826)、绞合线长(0.756)和壳长(0.846),与闭壳肌重相关系数最大的分别是壳宽(0.801)、壳高(0.922)、壳长(0.957)、壳高(0.966)和壳高(0.983)。经济性状中,性腺重与其他性状间的相关系数最小,湿重与其他性状间的相关系数最大。
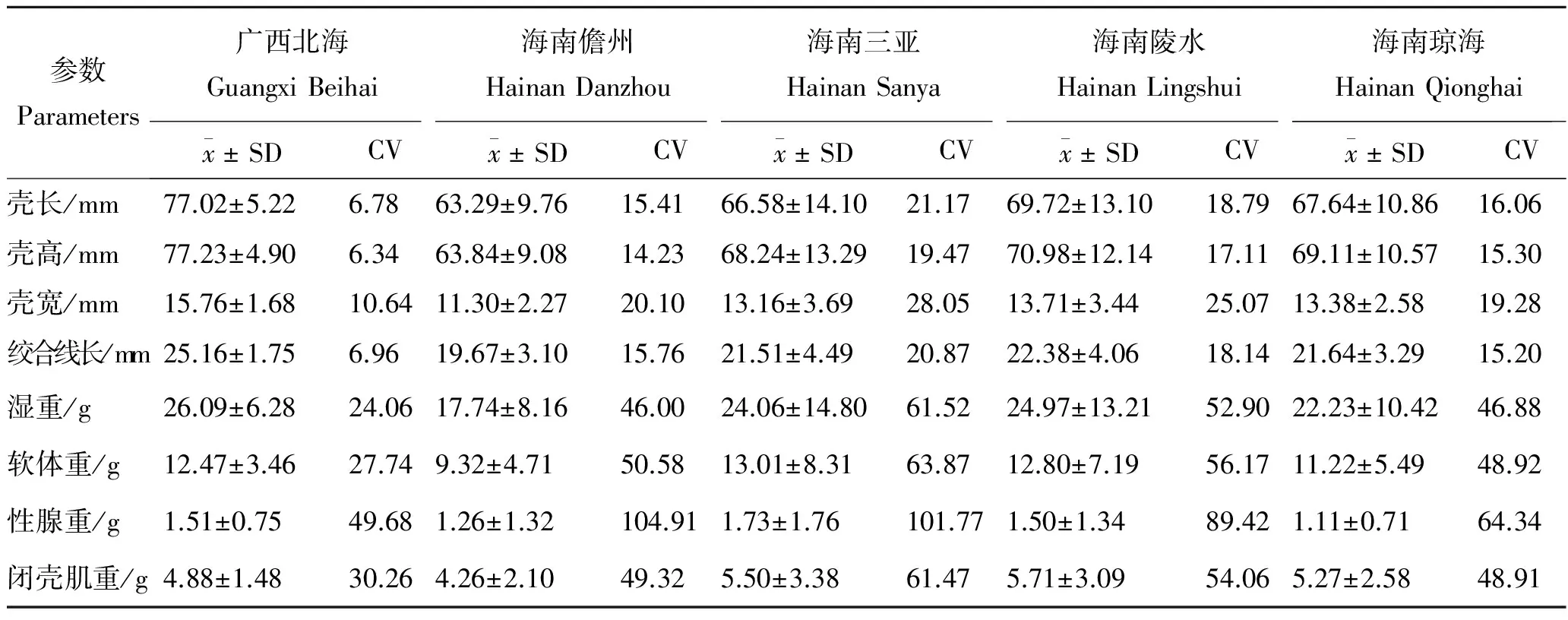
表1 5个地区长肋日月贝的各性状的描述性统计Tab.1 The phenotypic parameter statistics of A.pleuronectes from 5 regions
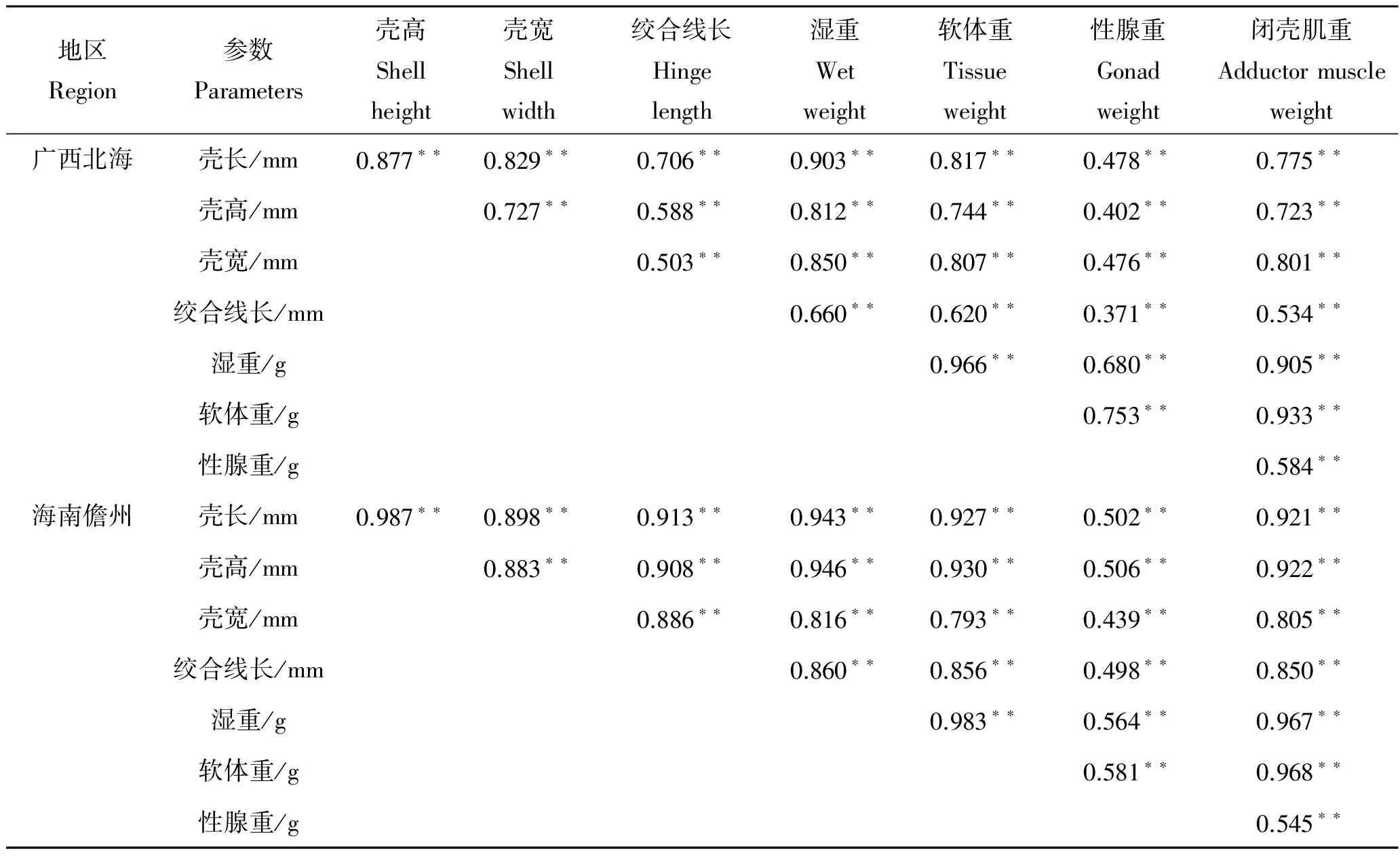
表2 5个地区长肋日月贝各性状间的相关系数Tab.2 Correlation coefficients between the phenotypic traits of A.pleuronectes from 5 regions
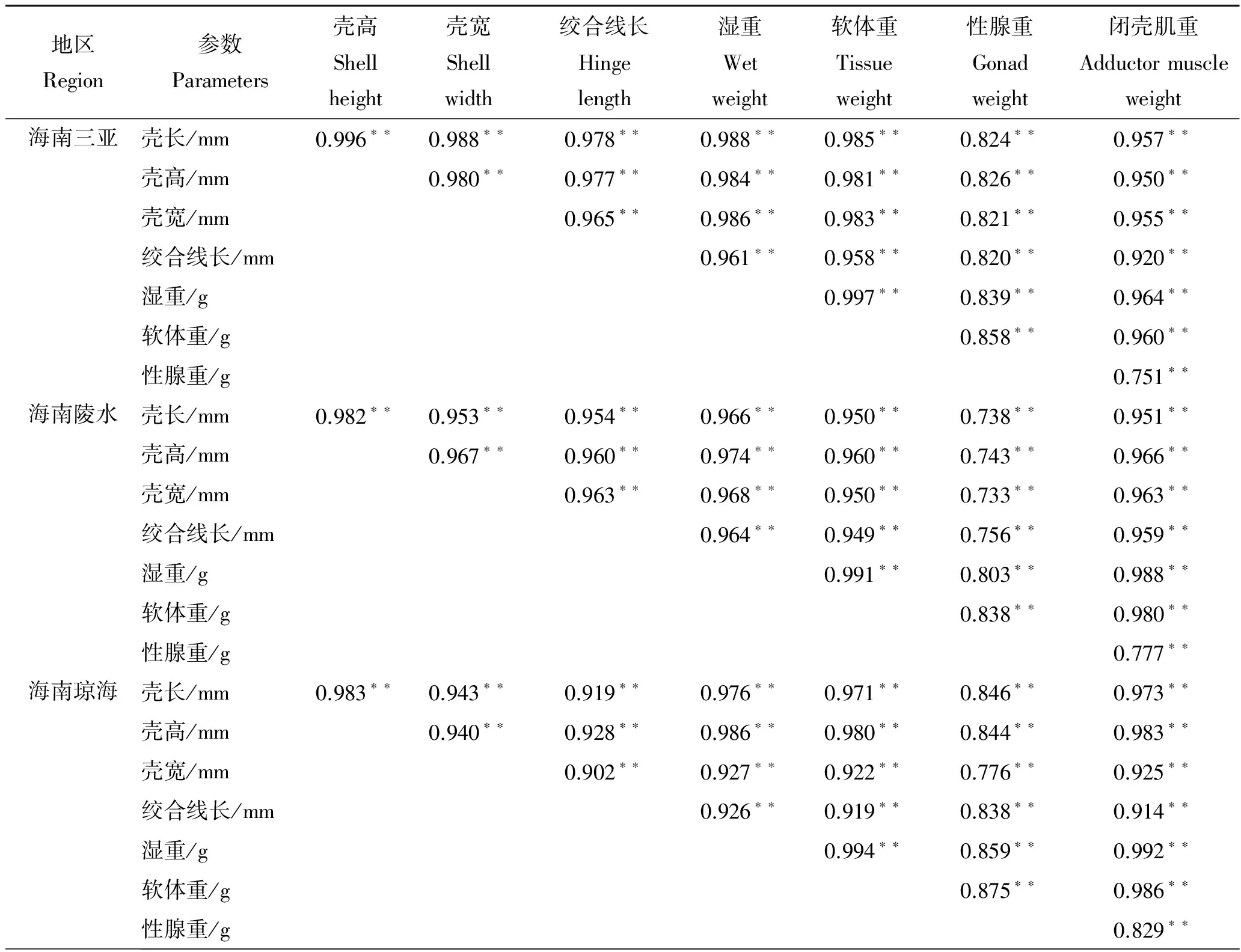
续表2,Tab.2 continued
注:“**”示相关性系数差异极显著(P<0.01),“*”示相关性系数差异显著 (P<0.05),下同。
2.3 形态性状对主要经济性状的通径分析
在相关系数中剔除其他形态性状对湿重的直接作用,得到其他性状对湿重的间接作用如表3。其中,广西北海、海南儋州、海南三亚、海南陵水和海南琼海长肋日月贝对湿重直接影响最大的形态性状分别是壳长(0.635)、壳高(0.565)、壳宽(0.548)、壳高(0.470)和壳高(0.785),间接影响最大的分别是壳长通过壳宽(0.526)、壳高通过壳长(0.558)、壳宽通过壳高(0.537)、壳高通过壳宽(0.454)和壳高通过壳长(0.772)。
在相关系数中剔除其他形态性状对软体重的直接作用,得到其他性状对软体重的间接作用如表4。其中,广西北海、海南儋州、海南三亚、海南陵水和海南琼海的长肋日月贝对软体重直接作用最大的形态性状分别是壳宽(0.529)、壳高(0.574)、壳长(0.584)、壳高(0.617)和壳高(0.764),间接作用最大的分别是壳宽通过壳高(0.385)、壳高通过壳长(0.567)、壳长通过壳宽(0.577)、壳高通过绞合线长(0.592)和壳高通过壳长(0.751)。
在相关系数中剔除其他形态性状对性腺重的直接作用,得到其他性状对性腺重的间接作用如表5。其中,广西北海、海南儋州、海南三亚、海南陵水、海南琼海的长肋日月贝对性腺重影响最大的形态性状分别是壳宽(0.476)、壳高(0.506)、壳高(0.826)、绞合线长(0.756)和壳长(0.737),海南琼海长肋日月贝对性腺重间接作用最大的是壳长通过壳宽(0.695)间接影响。
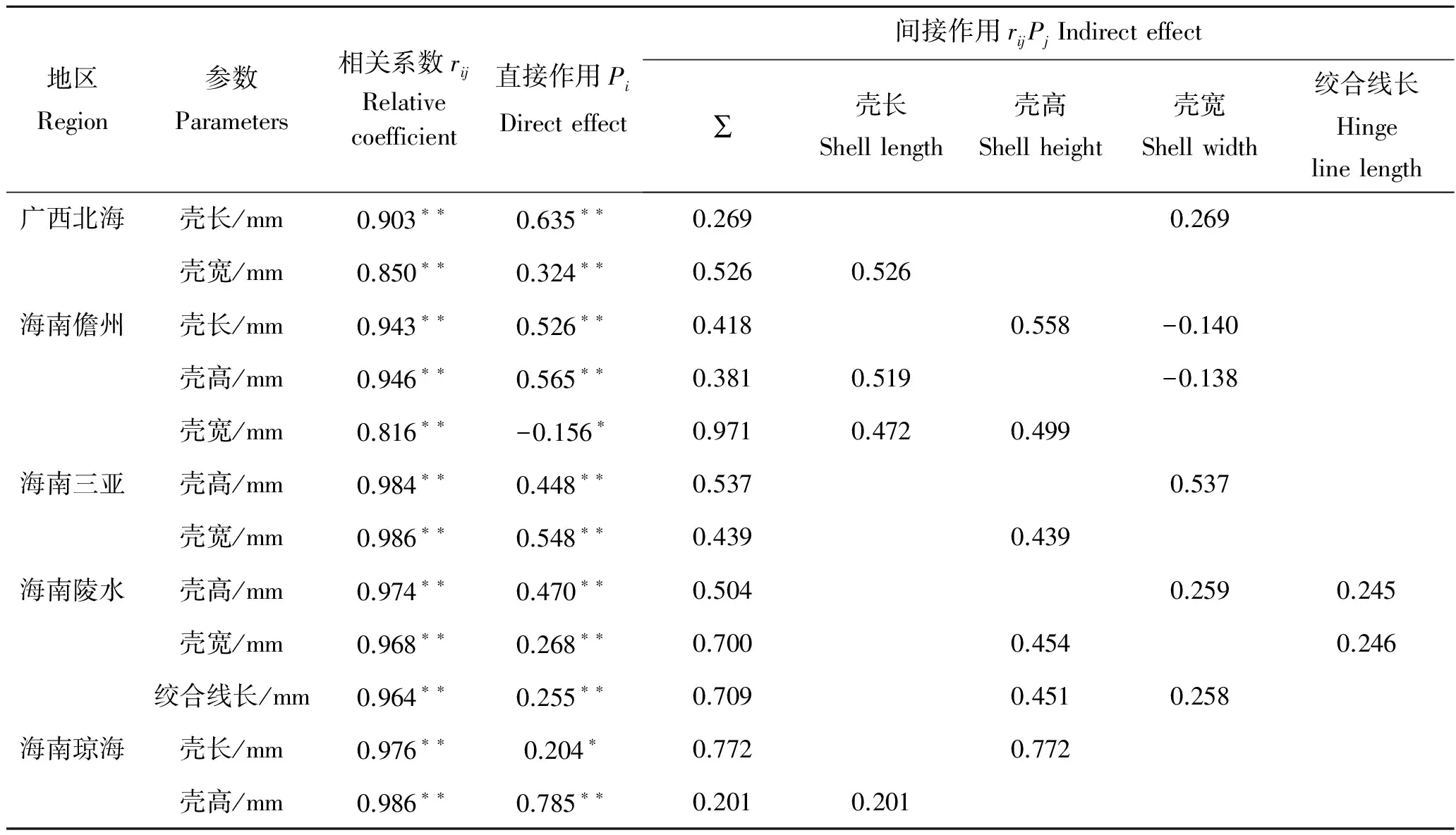
表3 各形态性状对湿重的通径分析Tab.3 The path coefficients of the morphometric to the wet weight of A.pleuronectes
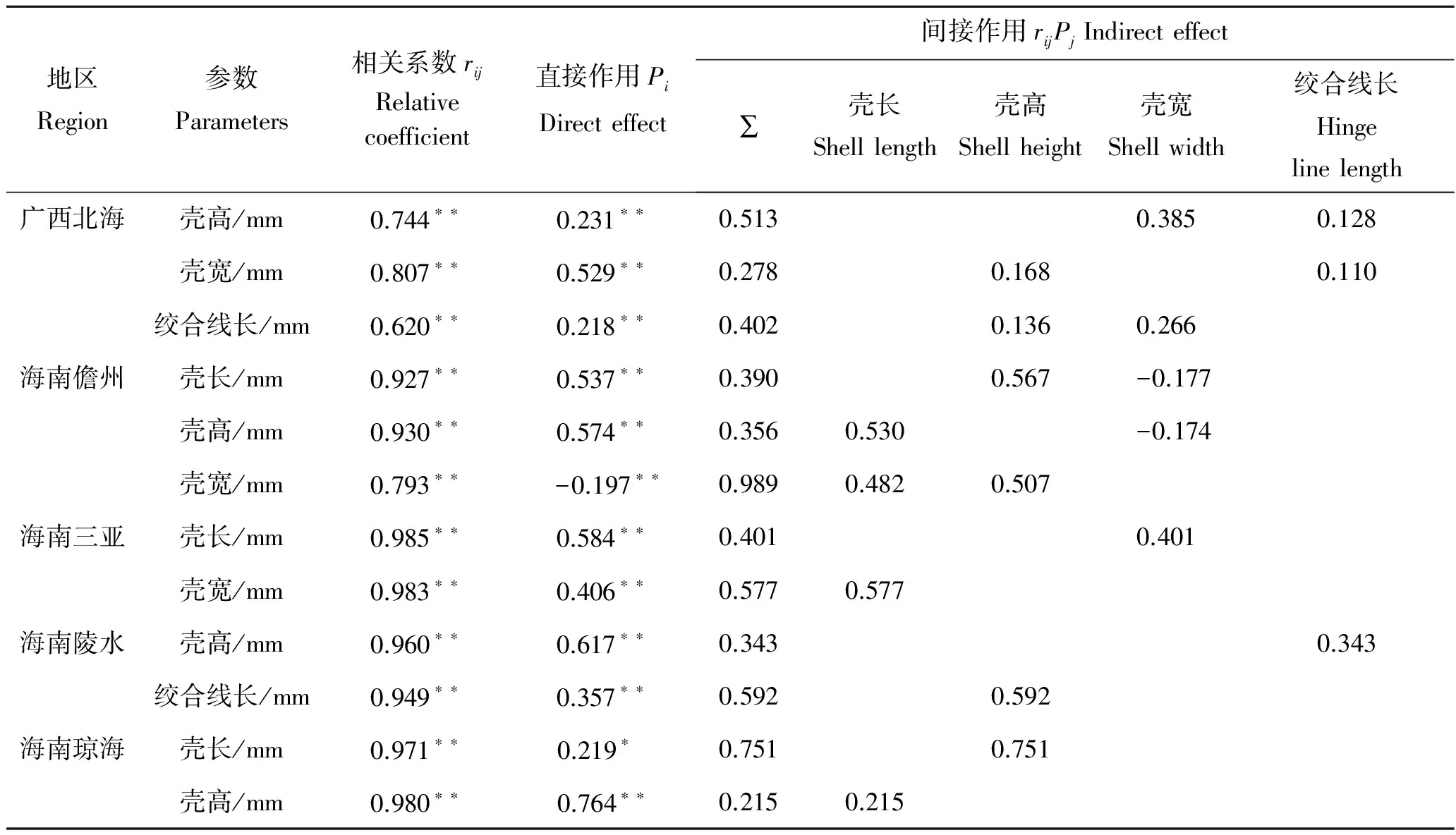
表4 各形态性状对软体重的通径分析Tab.4 The path coefficients of the morphometric to the tissue weight of A.pleuronectes
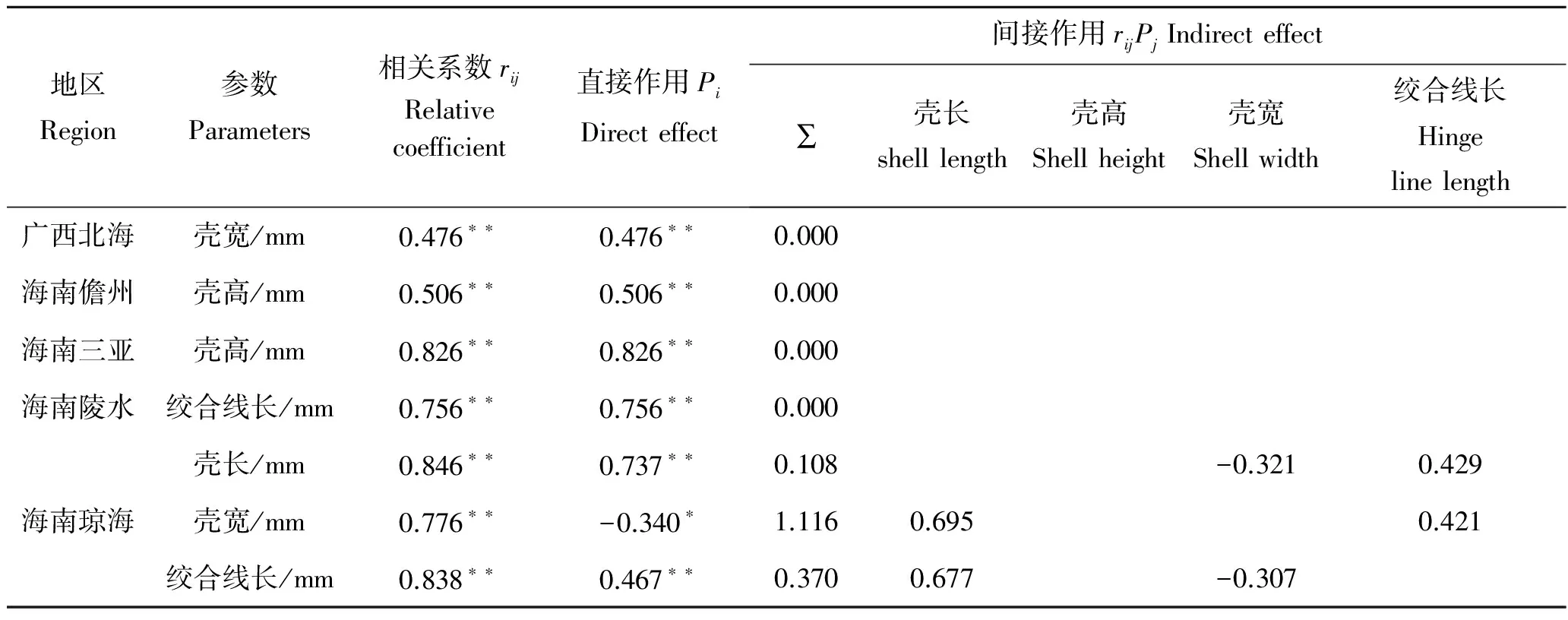
表5 各形态性状对性腺重的通径分析Tab.5 The path coefficients of the morphometric to the gonad weight of A.pleuronectes
在相关系数中剔除其他形态性状对闭壳肌重的直接作用,得到其他性状对闭壳肌重的间接作用如表6。其中,广西北海、海南儋州、海南三亚、海南陵水和海南琼海长肋日月贝对闭壳肌重中直接影响作用最大的分别是壳宽(0.584)、壳高(0.922)、壳长(0.957)、壳高(0.418)和壳高(0.790),对广西北海、海南陵水和海南琼海长肋日月贝间接影响最大的分别是壳宽通过壳高(0.425)、壳高通过壳宽(0.404)和壳高通过壳长(0.777)。
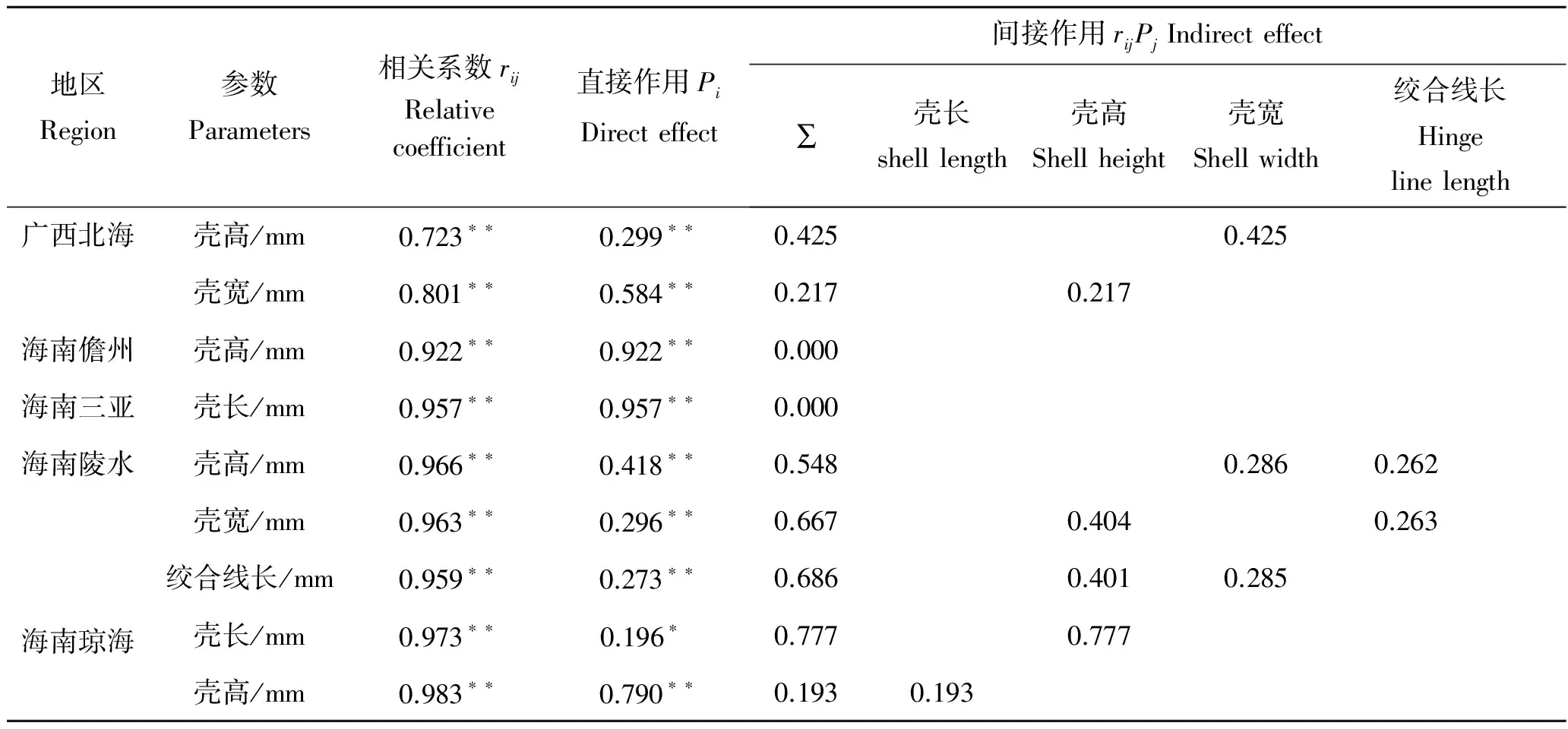
表6 长肋日月贝各性状对闭壳肌重的通径分析Tab.6 The path coefficients of the morphometric to the adductor muscle weight for A.pleuronectes
2.4 形态性状对主要经济性状的决定程度
5个地区中被保留的形态性状对各经济性状直接作用单独或两两共同对经济性状的决定系数如表7~10。其中,对角线上的数据是单性状决定系数,对角线上方的数据是两个形态性状共同对经济性状的决定系数。5个地区中对经济性状决定系数最大的单独性状有所不同,其中海南儋州、海南陵水和海南琼海对湿重决定系数最大的是壳高,广西北海是壳长,海南三亚是壳宽。海南儋州、海南陵水、海南琼海对软体重决定系数最大的是壳高,广西北海是壳宽,海南三亚是壳长。广西北海对性腺重决定系数最大的是壳宽,海南儋州和海南三亚是壳高,海南陵水是绞合线长,海南琼海是壳长。广西北海对闭壳肌决定系数最大的是壳宽,海南儋州、海南陵水和海南琼海是壳高,海南三亚是壳长。
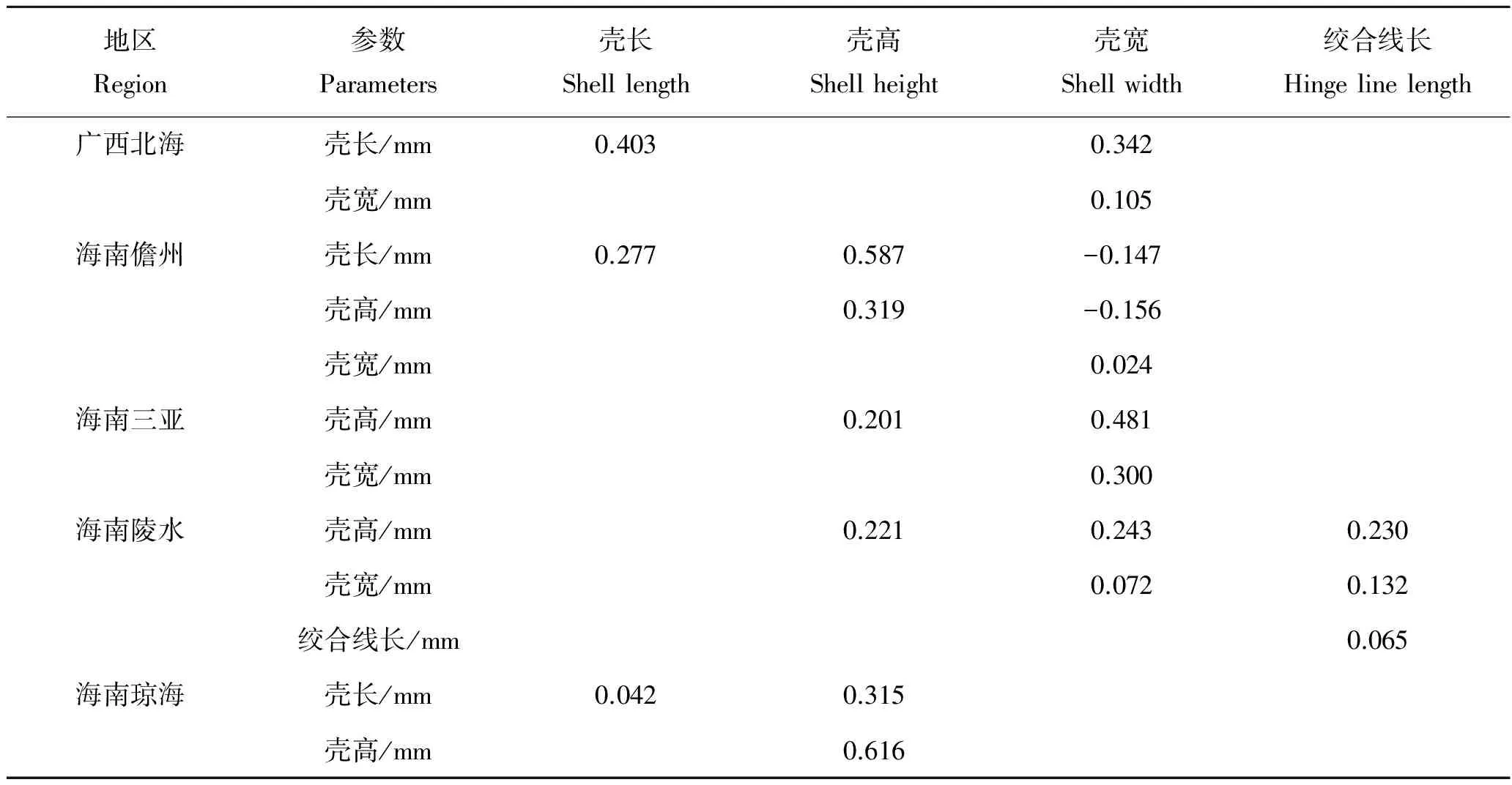
表7 各形态性状对湿重的决定系数Tab.7 The determinant coefficients of morphometric traits to the wet weight of A.pleuronectes
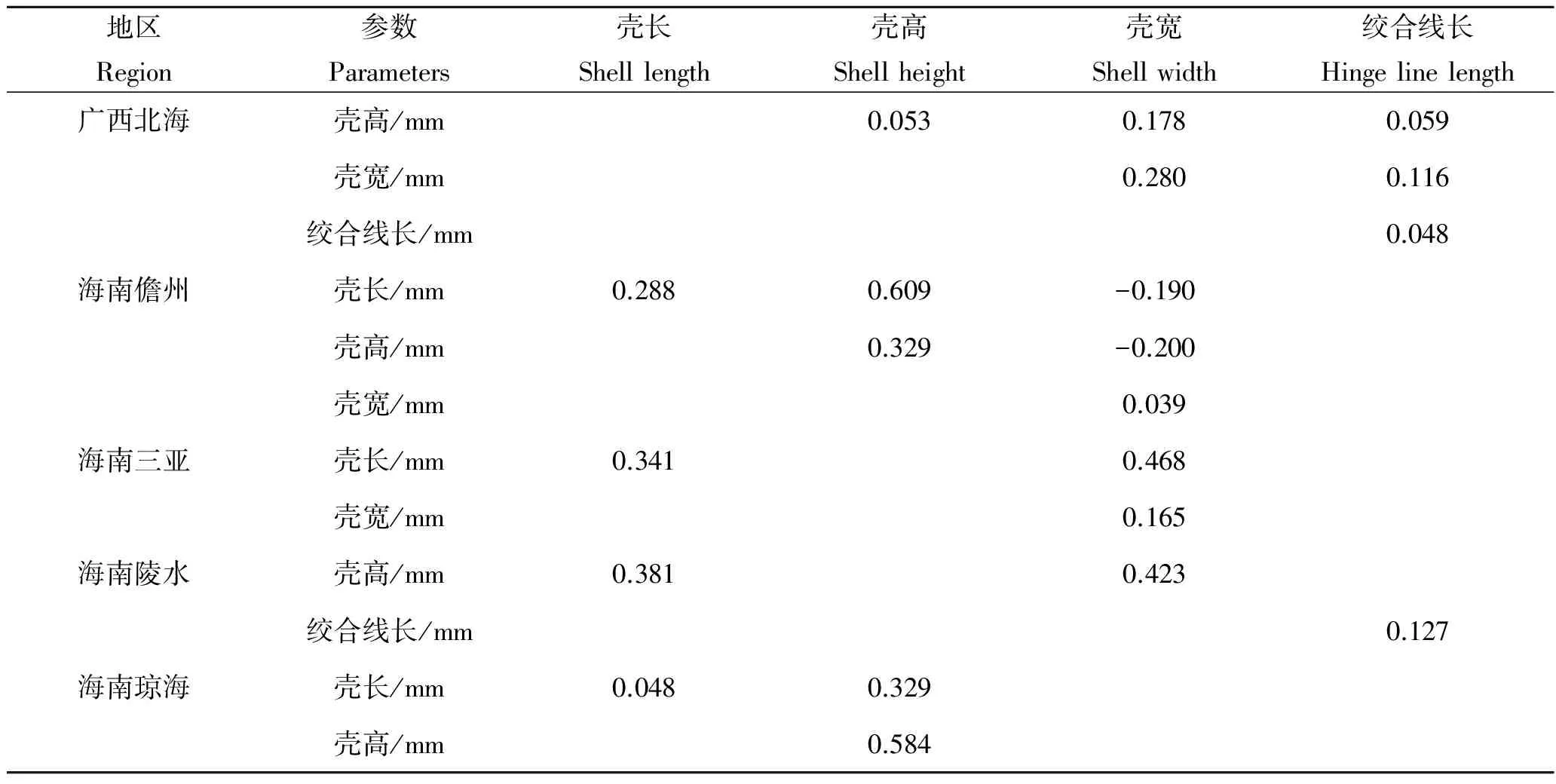
表8 各形态性状对软体重的决定系数Tab.8 The determinant coefficients of morphometric traits to the tissue weight of A.pleuronectes
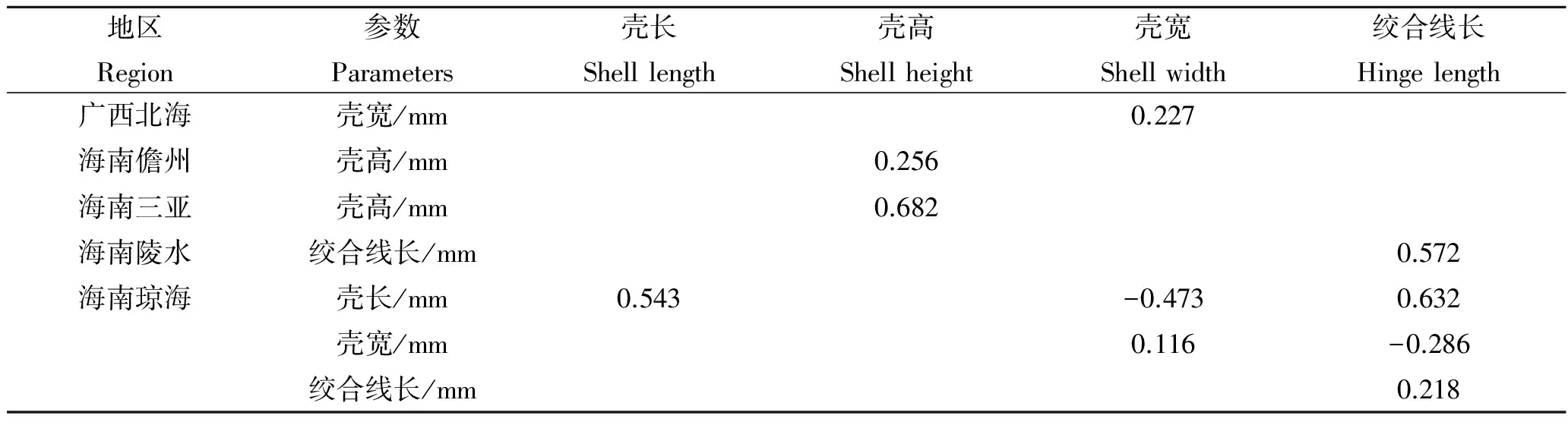
表9 各形态性状对性腺重的决定系数Tab.9 The determinant coefficients of morphometric traits to the gonad weight of A.pleuronectes
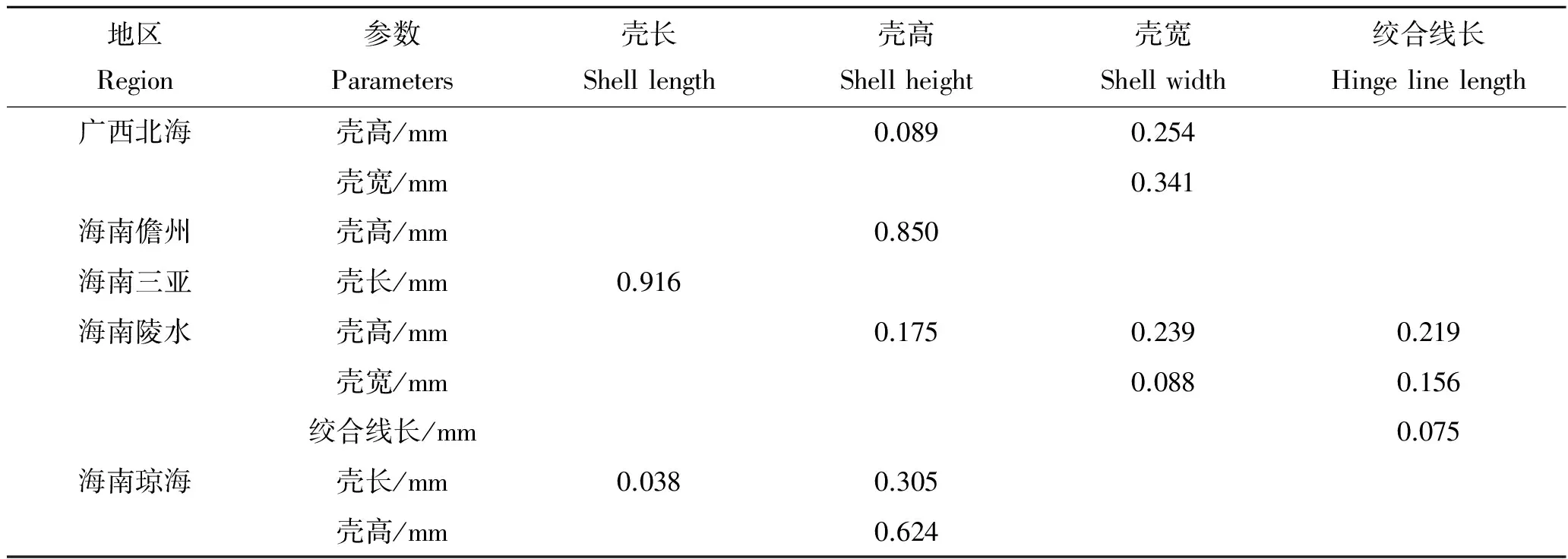
表10 各形态性状对闭壳肌重的决定系数Tab.10 The determinant coefficients of morphometric traits to the adductor muscle weight of A.pleuronectes
2.5 多元回归方程建立及显著性检验
5个地区建立以壳长(x1/mm)、壳高(x2/mm)、壳宽(x3/mm)和绞合线长(x4/mm)分别对湿重(y1/g)、软体重(y2/g)、性腺重(y3/g)和闭壳肌重(y4/g)的多元回归方程,如下所示。
广西北海,对湿重:y1= 0.763x1+ 1.212x3- 51.759,R2=0.848;对软体重:y2= 0.163x2+ 1.091x3+ 0.430x4- 28.146,R2=0.734;对性腺重:y3= 0.213x3-1.848,R2=0.226;对闭壳肌重:y4= 0.090x2+ 0.514x3- 10.182,R2= 0.684。
海南儋州,对湿重:y1= 0.440x1+ 0.507x2- 0.559x3- 36.169,R2=0.904;对软体重:y2= 0.259x1+ 0.298x2- 0.408x3- 21.504,R2=0.876;对性腺重:y3= 0.073x2- 3.433,R2=0.257;对闭壳肌重:y4= 0.213x2- 9.348,R2=0.850。
海南三亚,对湿重:y1= 0.499x2+ 2.198x3- 38.880,R2= 0.981;对软体重:y2= 0.344x1+ 0.913x3- 21.936,R2=0.975;对性腺重:y3= 0.109x2- 5.726,R2=0.682;对闭壳肌重:y4= 0.229x1- 9.768,R2=0.915。
海南陵水,对湿重:y1= 0.511x2+ 1.030x3+ 0.832x4- 44.019,R2=0.963;对软体重:y2= 0.365x2+ 0.634x4- 27.314,R2=0.931;对性腺重:y3= 0.250x4- 4.096,R2=0.572;对闭壳肌重:y4= 0.106x2+ 0.265x3+ 0.208x4- 10.123,R2=0.950。
海南琼海,对湿重:y1= 0.196x1+ 0.774x2- 44.497,R2=0.973;对软体重:y2= 0.111x1+ 0.397x2- 23.710,R2=0.962;对性腺重:y3= 0.048x1- 0.094x3+ 0.101x4- 3.096,R2=0.750;对闭壳肌重:y4= 0.047x1+ 0.193x2- 11.185,R2=0.968。
多元线性回归方差分析表明,广西北海、海南儋州、海南三亚、海南陵水和海南琼海的长肋日月贝湿重与形态性状之间的多元回归,其中,F值分别为248.824、300.835、1 465.035、839.704和1 745.942;软体重与形态性状之间的多元回归,其中,F值分别为80.932、225.202、1101.639、652.302和1 224.722;性腺重与形态性状之间的多元回归,其中,F值分别为26.318、33.811、124.353、130.892和96.194;闭壳肌重与形态性状之间的多元回归,其中,F值分别为96.352、555.906、624.906、614.189和1 459.065。经济性状与形态性状之间的回归均达到极显著水平(P<0.01),方差分析结果有显著差异,说明方差分析是有意义的,可以应用于实际生产中。
5个地区中被保留下来的形态性状与经济性状间的复相关系数如表11,从所得R2表明,5个地区被保留的形态性状与湿重、软体重和闭壳肌重间的关系极为密切,这些性状可以认为是影响相应群体湿重、软体重和闭壳肌重的主要性状。
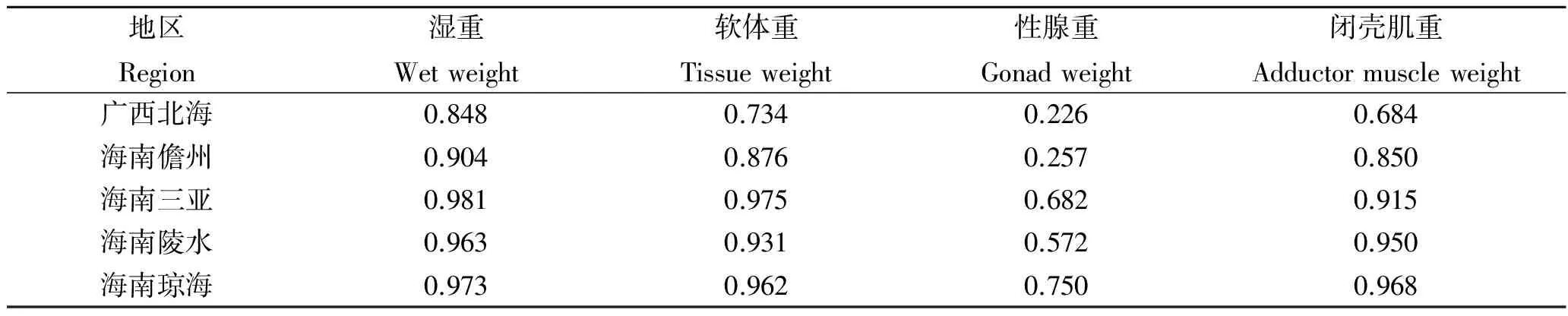
表11 各形态性状与主要经济性状的复相关系数R2Tab.11 The multiple-correlation coefficients of morphometric traits on the main economic traits
2.6 不同地区长肋日月贝形态分类
基于形态特征,将长肋日月贝5个地区进行欧式最短距离聚类分析,结果如图2,其中海南三亚、海南琼海和海南陵水的长肋日月贝形态特征最为接近,其次是与海南儋州的形态距离次之,与广西北海的长肋日月贝形态距离最远。
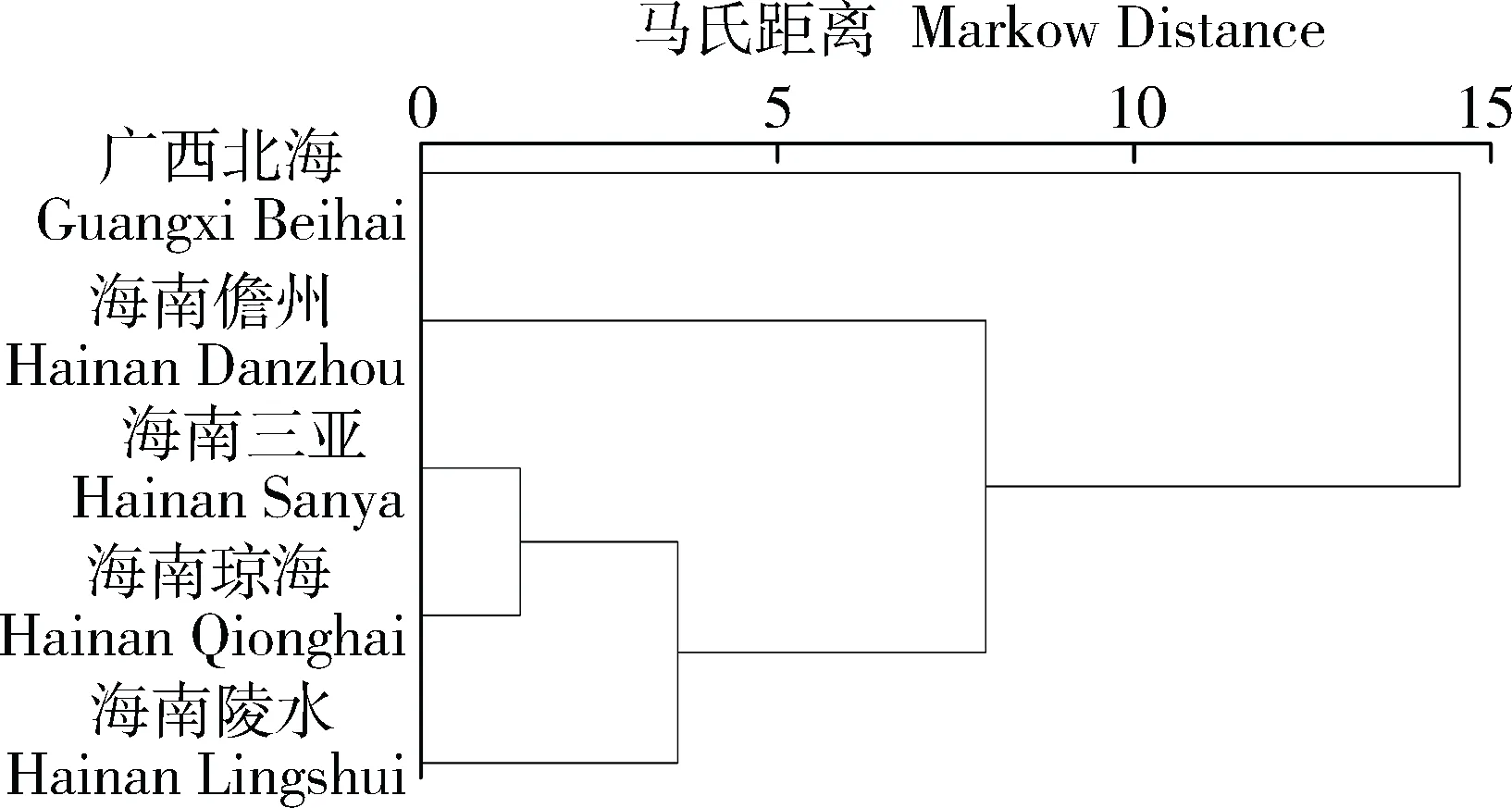
图2 长肋日月贝5个地区形态聚类分析Fig.2 Cluster dendrogram of A. pleuronectes from 5 regions
3 讨论
长肋日月贝经济性状的变异系数均大于其形态性状,结果与其他贝类表现一致[26-29];经济性状中性腺重变异系数大于其他性状,这与符玉等[17]和王雨等[30]对长肋日月贝的研究结果相同。刘志刚等[31]曾对马氏珠母贝(Pinctadamartensi)同一个体繁殖前后的质量称量发现其差异可达10%;对大小相同的同一批次,不同繁殖期及生理状况下检测到其质量差异可达20 %以上。长肋日月贝与其他大部分海洋双壳贝类一样,成体贝一年四季均可繁殖[30],同一个体在排卵(精)前后性腺重差异较大,由此性腺重变异系数较大。性腺重的改变在一定程度上也影响着湿重和软体重,因此湿重和软体重的变异系数也相应增大。闭壳肌是长肋日月贝良种选育中极其重要的经济性状,相同个体在不同生理状况下对闭壳肌重的影响较小,闭壳肌重变异系数比形态性状大,这在长肋日月贝大型闭壳肌良种选育中有着巨大的潜力。
本研究中,不同地区的野生长肋日月贝,表现出不同的体型特征,影响其主要经济性状的形态性状有所不同[30, 32]。该研究中,选取了长肋日月贝的壳长、壳高、壳宽和绞合线长对经济性状进行分析。结果表明,不同地区的长肋日月贝对各经济性状相关系数最大的形态性状不尽相同,呈现出地区差异,这与日本沼虾[15]和海湾扇贝(Argopectenirradians)[26]不同群体的形态性状对经济性状相关系数所呈现的结果类似。相关系数是变量间相互关系的体现,由于未剔除其他因素的影响,可导致自变量对因变量的直接作用和通过其他自变量对因变量的正负作用可以抵消,因此不能全面地解释变量间的相互关系。然而通径系数是变量标准化后的偏回归系数,能有效地区分直接影响和间接影响[3]。在相关分析的基础上再进行通径分析和决定系数的分析,刘小林等[33]认为只有复相关系数或各自变量对因变量的单独决定系数及两两共同决定系数的总R2大于或等于0.85时,表明影响因变量的主要自变量已经找到。本研究中海南儋州、海南三亚、海南陵水和海南琼海保留的形态性状对湿重、软体重和闭壳肌重的复相关系数均大于0.85,表明保留的形态性状是影响其相应经济性状的主要性状。广西北海被保留的形态性状对湿重、软体重和闭壳肌重的复相关系数略小于0.85,表明影响其相应经济性状的形态性状已基本找到。5个地区被保留的形态性状对性腺重的复相关系数皆小于0.85,表明形态性状与性腺重之间可能没有直接或必然的联系。王雨等[30]研究认为长肋日月贝成贝周年均可繁殖,排卵(精)受潮水周期影响,因此认为性腺重与个体性腺所处发育时期关系更为密切。
5个地区长肋日月贝形态特征经欧式最短距离聚类分析,其中海南三亚、海南琼海和海南陵水的长肋日月贝在形态特征上较为接近,其次是与海南儋州,与广西北海距离最远。形态特征距离与朱彩艳等[25]研究的海南4个地理群体长肋日月贝遗传间距的远近基本相符。由此揭示了不同地区的同种生物在形态特征上的差异,聚类分析结果与地理位置距离的远近有关,这与黄有辉等[15]研究的日本沼虾在形态特征上的差异与地理位置上的远近所表现出的结果相同。
4 结论
不同地区长肋日月贝,影响其主要经济性状的形态性状不尽相同,其中,直接影响其湿重作用最大的性状是海南儋州、海南陵水和海南琼海的壳高,广西北海的壳长以及海南三亚的壳宽;直接影响其软体重作用最大的是海南儋州、海南陵水和海南琼海的壳高,广西北海的壳宽以及海南三亚的壳长;直接影响其性腺重作用最大的是海南儋州和海南三亚的壳高,广西北海的壳宽,海南陵水的绞合线长以及海南琼海的壳长;直接影响其闭壳肌重作用最大的是海南儋州、海南陵水和海南琼海的壳高,广西北海的壳宽以及海南三亚的壳长。在对目标经济性状进行选育时,建议应当针对不同地区做相应的统计分析,以便选出最佳的形态性状。随着地理位置间距的增大,形态特征差异越大,由此,在进行选择育种中应尽量避免不同地区间的杂交;然而,在杂交育种中,为了体现杂种优势,应该选择距离较远的两个种群进行杂交。
参考文献:
[1]杜美荣, 方建光, 高亚平, 等. 不同贝龄栉孔扇贝数量性状的相关性和通径分析 [J]. 水产学报, 2017, 41(4): 580-587.
[2]董世瑞, 孔杰, 万初坤, 等. 中国对虾形态性状对体重影响的通径分析 [J]. 海洋水产研究,2007, 28(3): 15-22
[3]郭华阳, 陈明强, 王雨, 等. 黄边糙鸟蛤野生群体主要经济性状间的相关性及通径分析[J]. 南方水产科学, 2013, 9(2): 1-8.
[4]张伟杰, 常亚青, 丁君, 等. 日本镜蛤(DosiniajaponicaReeve)壳尺寸与质量性状的相关与回归分析 [J]. 海洋与湖沼, 2013, 44(3): 796-800.
[5]张根芳, 张文府, 方爱萍, 等. 养殖条件下不同年龄背瘤丽蚌(Lamprotulaleai)数量性状的相关与通径分析 [J]. 海洋与湖沼, 2014, 45(5): 1115-1121.
[6]王志铮, 吴一挺, 杨磊, 等. 日本沼虾(Macrobrachiumnipponensis)形态性状对体重的影响效应 [J]. 海洋与湖沼, 2011, 42(4):612-618.
[7]李鸿鹏, 富裕, 任夙艺, 等. 舟山近海日本囊对虾野生群体与越冬养殖群体形态性状对体重和肉重影响的比较 [J]. 海洋与湖沼, 2015, 46(5): 1218-1227.
[8]张嘉丽, 王庆恒, 邓岳文, 等. 斧文蛤(Meretrixlamarchii)形态性状对体质量的影响效果分析 [J]. 渔业科学进展, 2014, 35(6): 110-113.
[10]白临建, 杨爱国, 周丽青, 等. 栉江珧形态性状对重量性状的影响 [J]. 渔业科学进展, 2012, 33(6): 87-92.
[11]Ma H Y, Ma C Y, Ma L B, et al. Correlation of growth-related traits and their effects on body weight of the mud crab (Scyllaparamamosain)[J]. Genet Mol Res, 2013, 12(4):4127-4136.
[12]Zhao L, He Y, Yang F, et al. Correlation and path analysis of morphological and weight traits in marine gastropodGlossaulaxreiniana[J]. Chin J Oceanol Limn, 2014, 32(4):821-827.
[13]Luo X, Ke C, You W. Estimates of correlations for shell morphological traits on body weight of interspecific hybrid abalone (HaliotisdiscushannaiandHaliotisgigantea) [J]. J Shellfish Res, 2013,32(1): 115-118.
[14]肖述, 符政君, 喻子牛. 香港巨牡蛎雌雄群体的数量性状通径分析 [J]. 南方水产科学, 2011, 7(4): 1-9.
[15]黄有辉, 范斌, 李一鸣, 等. 日本沼虾五群体形态性状对体质量的通径分析[J]. 水产学报, 2016, 40(8): 1173-1185.
[16]杨文, 蔡英亚, 邝雪梅. 中国南海经济贝类原色图谱 [M]. 北京: 中国农业出版社, 2013: 168.
[17]付玉, 颜云榕, 卢伙胜, 等. 北部湾长肋日月贝的生物学性状与资源时空分布 [J]. 水产学报, 2012, 36(11): 1694-1705.
[18]王祯瑞. 中国动物志 软体动物门 双壳纲 贻贝目 [M]. 北京:科学出版社, 2002.
[19]朱彩艳, 王雨, 张殿昌, 等. 长肋日月贝的营养成分分析及评价 [J]. 海洋科学, 2011, 35(3): 87-91.
[20]郭华阳, 王雨, 陈明强, 等. 盐度、饵料密度对长肋日月贝滤水率的影响 [J]. 广东农业科学, 2012, 15: 144-146.
[21]叶乐, 赵旺, 王雨, 等. 盐度与pH对长肋日月贝幼虫存活及生长的影响 [J]. 南方农业学报, 2015, 46(9): 1698-1703.
[22]王雨, 叶乐, 杨其彬, 等. 长肋日月贝个体发生观察及人工育苗初步试验 [J]. 南方水产, 2009, 5(1): 36-41.
[23]蒲利云, 陈傅晓, 曾关琼, 等. 长肋日月贝亲本促熟培育与催产技术研究 [J]. 福建水产, 2012, 34(5): 405-409.
[24]Belda C A, Del Norte A G C. Notes on the induced spawning and larval rearing of the Asian moon scallop,Amusiumpleuronectes(Linné), in the laboratory [J]. Aquaculture, 1988, 72(1/2): 173-179.
[25]朱彩艳, 江世贵, 杨其彬, 等. 海南4个野生地理群体长肋日月贝AFLP分析 [J]. 琼州学院学报, 2016, 23(2): 99-104.
[26]孙俭, 王国栋, 周玮, 等. 三个海湾扇贝群体可量性状对闭壳肌重的通径分析[J]. 河北渔业, 2017(3):31-36.
[27]韩自强, 李琪. 长牡蛎壳橙品系形态性状与体质量的相关性及通径分析[J]. 中国海洋大学学报, 2017, 47(12): 46-52.
[28]杨蕊, 赵旺, 张欣, 等. 华贵栉孔扇贝两个群体的养殖效果评价及通径分析[J]. 水产科技情报, 2017, 44(1): 20-24.
[29]方军, 肖国强, 张炯明, 等. 两种壳色文蛤壳形态性状对活体质量重的影响[J]. 大连海洋大学学报, 2017, 32(3):310-314.
[30]王雨, 叶乐, 陈旭, 等. 海南野生长肋日月贝形态性状与重量性状的通径分析[J]. 安徽农业科学, 2009, 37(8): 3570-3572.
[31]刘志刚, 王辉, 孙小真, 等. 马氏珠母贝经济性状对体重决定效应分析[J]. 广东海洋大学学报, 2007, 27(4):15-21.
[32]杨彬, 谢恩义, 贾柽. 北部湾长肋日月贝形态性状对活体质量的影响分析[J]. 水产养殖, 2013, 34(7): 14-18.
[33]刘小林, 吴长功, 张志怀, 等. 凡纳滨对虾形态性状对体质量的影响效果分析[J]. 生态学报, 2004, 24(4): 857-862.
猜你喜欢
法人(2021年12期)2021-05-09
水产养殖(2021年2期)2021-02-24
今日农业(2020年15期)2020-09-25
今日农业(2020年13期)2020-08-24
南方农业学报(2020年4期)2020-06-04
南方农业学报(2020年10期)2020-01-21
中学生英语·教师版(2019年10期)2019-11-05
科学与财富(2018年12期)2018-06-11
人民论坛(2016年15期)2016-06-24
人民论坛(2016年15期)2016-06-24
