
重金属Zn2+2种暴露方式对三角褐指藻生长繁殖及光系统II的影响
2018-04-11 09:00:50吴常文张建设
浙江海洋大学学报(自然科学版) 2018年6期
陈 晋,吴常文,张建设,周 超
(浙江海洋大学海洋科学与技术学院,国家海洋设施养殖工程技术研究中心,浙江舟山 316022)
随着沿海地区经济的迅速发展和海洋资源的开发利用,海洋污染日益严重,其中Cu2+、Zn2+、Cd2+等重金属离子在海水中的含量不断上升,重金属成为海洋的主要污染物。重金属污染物会对海洋生物造成持续不可逆的伤害,积累到一定限度更会危害到海洋生态系统的结构和功能,或经过食物链间接危害到人类健康。海洋微藻作为海洋主要的初级生产者,是海洋食物链的基础和起点。相较于海洋食物链中高营养级的生物,海洋微藻对重金属污染应激反应更为敏感。因此开展重金属对海洋微藻的毒害作用研究,对于建立相应的污染物监测标准具有重要重要现实意义。
近年来,有关重金属对海洋微藻毒害作用的研究已初见报道[1-2]。传统重金属毒害作用研究,通常集中于重金属半致死浓度(LC50)或者半抑制浓度(EC50)[3]以及该浓度胁迫下藻类相应的生理生化反应研究[4-5]。由于重金属在海洋中的浓度是常低于国标浓度,并保持长期低浓度刺激海洋生态系统,因此从低限值持续刺激角度切入,探究重金属污染对海洋生物的影响,对于完善海洋重金属污染的毒性胁迫机理具有重要作用。锌在生物体中起着双重作用,既是生物代谢必需的微量营养元素,又是一种一定浓度下具高毒性的重金属。有关锌对藻类生长的影响已有不少研究[1-4],而关于重金属低限值持续刺激下海洋微藻的毒性研究还鲜见报道。硅藻是一类重要的真核浮游生物,其光合产量占据海洋初级生产力的40%[6],具有重要的生态学地位。三角褐指藻Phaeodactylum tricornutum Bohlin作为硅藻门中1个重要种类,在水产养殖业中常被用作生物饵料,同时也是一种常用的毒性监测受试生物[3]。本文以硅藻-三角褐指藻为测试目标,探究Zn2+不同胁迫方式对三角褐指藻生长繁殖以及光合作用的影响,旨在掌握不同的刺激方式对海洋浮游植物初级生产力的影响情况。
1 材料与方法
1.1 藻类来源
三角褐指藻藻种购买自上海光语生物科技有限公司。
1.2 培养条件及试剂
培养用的锥形瓶经10%稀HCl浸泡过夜后,用蒸馏水反复冲洗后灭菌备用;培养用海水为自然海水,经自然沉淀后,用0.45 μm微孔滤膜抽滤经充分曝气消毒后使用。实验过程中,藻类采用F/2培养基,于GXZ-208B型恒温光照培养箱培养(宁波江南仪器厂),其中光照强度为6 000 lx,温度为20±2℃,光暗周期为 12 h:12 h。
实验过程中添加的ZnCl2(AR分析纯)为国药生产,配制ZnCl2溶液的水采用超纯水。实验过程中添加的ZnCl2溶液浓度分别为5.000 mg/mL及0.500 mg/mL。盐酸(HCl,AR分析纯)为沪试生产,超纯水稀释到10%备用。
1.3 实验方法
将10 mL对数生长期的三角褐指藻接种到装有90 mL新鲜的F/2培养基的250 mL锥形瓶中,起始藻液浓度为8×105cell/mL。实验组分为2组:一组为强刺激组(high-dosage single exposure,HSE),另一组为持续弱刺激组(low-dosage repeated exposure,LRE),HSE组:加入20 μL浓度为5.000 mg/mL的ZnCl2溶液(体积比为0.02%)到培养基,使Zn2+最终浓度为1 mg/L,接近三角褐指藻的半抑制浓度(EC50)[3-4];LRE组:每日加入20 μL浓度为0.500 mg/mL的ZnCl2溶液(体积比为0.02%),每天2次持续5 d,5 d后最终使培养基中Zn2+的浓度达到与高浓度组相同的1 mg/L;空白对照组(CK)无ZnCl2添加,所有的试验都设置3组平行试验组,在恒温光照培养箱中培养7 d。在暴露试验过程中每日摇晃3~4次,防止藻细胞附壁沉降。
1.3.1 三角褐指藻种群增长曲线及生长速率的测定
三角褐指藻种群数量采用血细胞计数板法对三角褐指藻藻细胞进行计数,其中生长速率(μ)采用以下公式[7]计算得到:

式中μ为生长速率,x1和x2分别代表培养时间为t1和t2时藻液的细胞浓度。
1.3.2 叶绿素a含量的测定
叶绿素a含量测定采用热乙醇法[8],取2 mL藻液,过GF/F滤膜后,-20℃冷冻过夜后取出迅速放入盛有95%的热乙醇的离心管,盖紧后80℃水浴2 min提取叶绿素。提取液经超声波振荡10 min,暗处理4 h后,于15 000 r/min、4℃下离心5 min。取上清液于紫外分光光度计(UV-2700,岛津)下,测定665 nm及750 nm处的吸光值,经1 mol/L的盐酸酸化后,再测得665 nm及750 nm处的吸光值。提取液中的叶绿素a浓度根据PORRA[9]的公式计算获得:

其中:E665和E750为酸化前的吸光值;A665和A750为酸化后的吸光值;27.9为叶绿素a在乙醇相中吸光常数;V乙醇为乙醇相的体积;V样品为样品溶液的体积。
1.3.3 快速叶绿素荧光动力学测定
叶绿素荧光参数测量采用浮游植物荧光仪(Phyto-PAM,Walz,德国)测定。每日取2 mL的三角褐指藻样品经暗适应 5 min后,在最大饱和脉冲光照为 4 000 μmol/(m2·s1)下,测得 F0,Fm,Fv/Fm以及 rETR 的数值。
其中:F0为初始叶绿素荧光产量(minimal fluorescence);Fm为暗适应后最大荧光产量(maximal fluorescence);Fv=Fm-F0;Fv/Fm为PSII的光能转换效率;rETR为相对光合电子传递率 (relative electron transport rate)。
1.4 数据处理
实验结果以平均值±标准误差(mean±S.E.)表示,差异显著性设置P<0.05。锌胁迫浓度差异采用t检验进行统计学处理。实验数据采用SPSS 18.0.0(Chicago,USA)收集和统计原数据,进行单因素方差分析及Duncan多重比较,α=0.05。
2 结果
2.1 不同暴露方式下重金属Zn2+对三角褐指藻种群生长状况的影响
三角褐指藻在重金属Zn2+不同暴露方式下其种群生长情况如图1所示。研究表明,对照组的三角褐指藻的生长曲线呈现典型的“S”型生长曲线,而HSE组和LRE组的三角褐指藻种群生长曲线出现明显的差异(P<0.05)。相较于对照组,在实验早期(1~3 d)HSE组三角褐指藻种群生长速率显著低于对照组和LRE组(P<0.05),从第4天开始进入补偿生长,生长速率逐渐恢复到对照组水平,第7天观测结束时仍维持较高的生长速率。LRE组三角褐指藻种群在1~4 d里的生长状况与对照组相近,第5天增速放缓,在第7天观测结束时其种群密度显著的低于HSE组(P<0.05)。
重金属Zn2+不同暴露方式下三角褐指藻种群增长速率如图2所示,LRE组与空白对照组的种群增长趋势一致,在第3天达到最大增长速率,并随着时间的变化速率逐渐下降。而HSE组最大增长速率出现在第4天,此后迅速下降;观测结束时,HSE组三角褐指藻种群增长速率与空白对照组及LRE组相差不大。

图1 三角褐指藻种群数量随时间变化图Fig.1 The population of P.tricornutum changed over time

图2 三角褐指藻种群增长速率随时间变化图Fig.2 The growth ratio of P.tricornutum changed over time
2.2 不同暴露方式下重金属Zn2+对三角褐指藻叶绿素a含量的影响
三角褐指藻在重金属Zn2+不同暴露方式下叶绿素a含量随时间变化情况如图3所示。空白对照组的叶绿素a含量随时间变化不断增加;HSE组的三角褐指藻叶绿素a含量随着时间变化先缓慢下降并显著低于空白对照组,第6天开始增长至正常水平,与空白对照组含量无显著差别;LRE组的三角褐指藻叶绿素a含量随着时间变化不断下降,第5天开始显著低于空白对照组。
2.3 不同暴露方式下重金属Zn2+对三角褐指藻光系统II的影响
重金属Zn2+不同暴露方式下对三角褐指藻PSII反应中心最大光化学效率(Fv/Fm)变化情况如图4A所示。在整个暴露实验过程中,空白对照组的Fv/Fm自第1天到第3天从52.8%增长到71.6%,此后持续的3 d时间内在70%上下波动;而HSE组和LRE组之间并无显著性差异,第3天开始两个实验组的Fv/Fm均要显著低于对照组(P<0.05)。从时间变化上来看,HSE组及LRE组并未呈现相关规律,二者在暴露实验过程中其Fv/Fm均在62%之间上下波动。
重金属Zn2+不同暴露方式下对三角褐指藻光合作用相对电子传递效率(rETR)如图4B所示。相较于对照组,HSE组在受到大剂量重金属Zn2+刺激下rETR抑制明显,并随着时间逐渐恢复到初期水平,最后第7天HSE组rETR要显著高于LRE组(P<0.05)。LRE组在暴露实验初期rETR值缓慢上升并略高于对照组,但在暴露实验第4天其rETR较前一天显著下降并低于低于对照组的值(P<0.05),随着重金属Zn2+浓度不断积累其rETR值也随之不断下降,并在第7天其rETR值显著低于对照组及HSE组(P<0.05)。
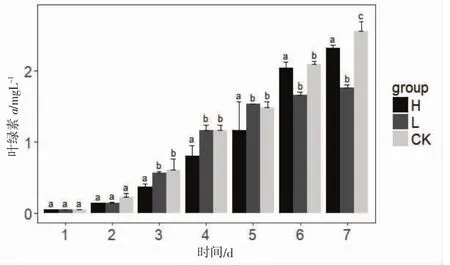
图3 三角褐指藻叶绿素a含量随时间变化图Fig.3 The Chlorophyll a of P.tricornutum changed over time

图4 三角褐指藻光系统II各参数(Fv/Fm、rETR)随时间变化图Fig.4 The Chlorophyll fluorescence parameters(Fv/Fm,rETR)of P.tricornutum changed over time
3 讨论
3.1 HSE组重金属Zn2+刺激对三角褐指藻的生物学效应
HSE组所设置的Zn2+浓度为三角褐指藻的EC50[3-4],高浓度的Zn不仅抑制相对光合电子传递效率而且使线粒体膜的功能性降低,叶绿素含量下降,最终影响藻类的生长[10]。在5 mg/L的重金属Zn2+刺激下,暴露初期(1~3 d)三角褐指藻种群生长受到了显著抑制,其叶绿素a含量及rERT也受到了不同程度的抑制作用。但在暴露实验中后期(4~7 d),HSE组三角褐指藻的种群生长和叶绿素a含量出现了类似“补偿效应”的增长,Zn2+对其抑制作用明显弱化,种群数量及叶绿素a含量恢复到正常水平。该现象与三角褐指藻在N、P限制[11]以及UV胁迫[12]解除时出现的种群生增长状况相似。推测有可能是微藻的吸附作用[13-14]或者代谢作用[5]降低了高浓度的重金属Zn2+对三角褐指藻的毒害作用,进而出现补偿生长的现象,试验结果需要进一步验证。另一方面根据HSE组的rETR虽然在暴露后期有所恢复但是其值仍显著低于对照组这一现象,推测有可能高浓度的Zn2+已经损伤细胞器,进而影响光合作用。
3.2 LRE组重金属Zn2+持续刺激对三角褐指藻的生物学效应
LRE组设置的Zn2+浓度为0.5 mg/L为国标[15]设置的水污染1.0 mg/L的一半,在重金属Zn2+的5 d持续刺激下,在暴露早期(1~3 d)三角褐指藻种群生长与空白对照组无异叶绿素a含量及叶绿素荧光参数也随之正常增长。这可能是由于Zn是微藻必需的微量元素之一,它对维持生命体的生长、代谢和酶活性等起着非常重要的作用。但随着封闭的培养液体系中重金属Zn2+的浓度不断上升,最终达到HSE组设置的浓度5 mg/L,LRE组三角褐指藻种群提前进入平台期,其叶绿素a含量及叶绿素荧光参数在第4天前后开始下降。这与PAULSSON,et al[16]在Göta Älv流域研究藻类重金属锌的长期积累性毒害作用,亦发现类似结果。研究发现,7 d时LRE组rETR值远低于对照组以及HSE组。研究表明,低浓度Zn2+污染(低于国标水污染浓度)在海水中对三角褐指藻亦具有积累性毒害作用。
4 结论
本文比较了HSE和LRE两种暴露方式下重金属Zn2+对三角褐指藻的生态毒性效应,结果表明HSE的胁迫方式在暴露起始阶段对三角褐指藻的毒性表现最为明显,并具有时效性;而LRE的胁迫方式在整个暴露实验中在叶绿素a含量和PSII电子传递方面表现出显著的积累性变化。研究发现,长期低浓度的重金属Zn2+在水体中能造成三角褐指藻持续的毒害作用甚至造成细胞损伤,进而影响光合作用。
猜你喜欢
今日农业(2022年15期)2022-09-20 06:54:16
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
看世界·学术下半月(2020年7期)2020-09-10 07:22:44
中国果业信息(2019年1期)2019-01-05 17:41:42
红土地(2018年7期)2018-09-26 03:07:38
广东农业科学(2017年10期)2018-01-25 03:22:15
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
生物学教学(2017年9期)2017-08-20 13:22:32
甘肃农业科技(2015年12期)2015-04-22 02:57:24
食品工业科技(2014年6期)2014-05-10 06:04:50
