
重组水疱性口炎病毒载体病毒包装体系的建立及优化
2018-04-09 03:37卢建博郑文铝余云舟叶建强戴秋云刘珠果
生物技术通讯 2018年2期
卢建博,郑文铝,,余云舟,叶建强,戴秋云,刘珠果
1.军事医学研究院 生物工程研究所,北京 100071;2.扬州大学 兽医学院,江苏 扬州 225100
水疱性口炎病毒(vesicular stomatitis virus,VSV)属弹状病毒科,为不分节段的单股负链RNA病毒。负链RNA病毒的包装需要RNA与核衣壳蛋白形成有活性的核糖核蛋白,以此作为RNA依赖的RNA聚合酶的模板,所以单纯的负链RNA无感染性,且病毒复制周期中没有出现DNA环节,病毒基因组不会整合到宿主细胞基因组中,因此来源于改构的VSV的重组VSV(rVSV)载体疫苗具有较高的安全性。近年来,随着流感、埃博拉病毒病和寨卡病毒病等大规模暴发,其疫苗的研制已被世界各国广泛关注。以rVSV为载体的疫苗已研究多年,具有免疫效率高、无预存免疫等优点,已用于人类免疫缺陷病毒、流感病毒[1]、尼帕病毒[1]、埃博拉病毒[2-5]、马尔堡病毒[6-7]和中东呼吸综合征冠状病毒[8]等的疫苗研究,其中以rVSV为载体的埃博拉疫苗已经完成Ⅲ期临床试验并提交美国FDA审批[9]。
此外,VSV能在多种细胞中高滴度快速生长,可建立VSV反向遗传操作系统[10-12],特别适合于丝状病毒及黄热病毒疫苗的设计及制备。表达外源基因的rVSV的稳定高效包装是制备rVSV载体疫苗的关键环节。提高病毒包装的成功率,除须准确合成病毒的基因组cDNA外,病毒包装细胞系的选择、rVSV骨架质粒和辅助质粒的用量、重组痘病毒(vTF7-3)的感染时间、转染试剂作用时间等均影响病毒的稳定高效包装。本实验室根据文献报道的条件进行了rVSV的包装,发现其包装效率较低,稳定性不高[13-16]。本研究中,我们对该系统包装条件进行探索及优化,为建立表达外源基因的rVSV载体疫苗奠定了坚实基础。
1 材料和方法
1.1 材料
BHK21-WI2细胞购自Kerafast公司;293T细胞购自Clontech公司;重组痘病毒vTF7-3购自ATCC公司;prVSVΔG-GFP骨架质粒、辅助质粒(pBS-N、pBS-P、pBS-G、pBS-L)、pCAGGS-G由MichaelA.Whitt教授馈赠;转染试剂 Lipo⁃fectAMINE 2000购自Invitrogen公司;Opti-MEM培养基、DMEM高糖培养基、胰蛋白酶(0.25%)、胎牛血清(FBS)购自Gibco公司;96孔细胞培养板购自Corning公司;6孔细胞培养板购自NEST公司;0.22 μm针头滤器购自Millipore公司;TH4-200型倒置荧光显微镜为Olympus公司产品。
1.2 rVSVΔG-GFP包装及扩增
1.2.1 细胞接种 以3.5×105~5×105/孔接种至6孔细胞培养板,于37℃、5%CO2细胞培养箱中培育14~16 h,使细胞密度达到85%~95%,同一细胞系后续操作中培育条件不变。
1.2.2 vTF7-3感染细胞 自-80℃取出冻存的vTF7-3至37℃快速解冻,加入37℃预热的无血清DMEM培养基,混匀病毒液,移除1.2.1项中的培养基,用37℃预热的无血清DMEM漂洗细胞,将病毒液接种至6孔细胞培养板(MOI=5),在培养箱中孵育1 h。
1.2.3 质粒体系配制 将prVSVΔG-GFP骨架质粒、辅助质粒(pBS-N、pBS-P、pBS-G、pBS-L)分别以5、3、5、8、1 μg/孔移取。
1.2.4 转染试剂配制及准备 将1.2.3项中的质粒移至含500 μL Opti-MEM培养基的1.5 mL离心管中混匀,记为A管;LipofectAMINE 2000与质粒以2.5~5 μL∶1 μg的比例添加到含500 μL Opti-MEM培养基的1.5 mL离心管中混匀,记为B管,室温静置5 min;将B管移入A管并混匀,室温静置20 min。
1.2.5 转染细胞并观察 弃除1.2.2项中的培养液,移入1.2.4项中转染试剂,轻摇混匀,培养6 h后更换为含5%FBS的DMEM培养基,继续培养48 h,荧光显微镜观察细胞病变(CPE)及荧光。
1.2.6 病毒收获 从6孔细胞培养板上刮下细胞,反复吹打,经0.22 μm针头滤器过滤,收集滤液,冻存于-80℃备用。
1.2.7 pCAGGS-G质粒转染BHK21-WI2细胞 按1.2.1项接种细胞,pCAGGS-G质粒为2 μg/孔,用LipofectAMINE 2000转染细胞,培养至细胞出现明显的合胞体。
1.2.8 病毒扩增 取1.2.6项中初次包装病毒至37℃快速解冻,按500 μL/孔移至1.2.7项中已弃除培养液的细胞中,于细胞培养箱中感染1 h,每隔15 min轻微摇晃,然后以1.5 mL/孔添加预热的含5%FBS的DMEM培养基,继续培养48 h,观察CPE及荧光变化。
1.3 rVSVΔG-GFP包装条件探索及优化
1.3.1 辅助质粒用量比较 选取293T细胞用于病毒包装,实验方法同1.2,辅助质粒的比例与方法1.2.3项中辅助质粒比例相同,用量在原基础上减至1/2(8.50 μg)、1/4(4.25 μg)、1/6(2.83 μg)、1/8(2.16 μg)、1/10(1.70 μg)。
1.3.2 质粒总用量比较 选取293T细胞用于病毒包装,实验方法同1.2,质粒的比例与方法与
1.2.3项相同,骨架质粒及辅助质粒总量均减为原用量的1/2(11.00 μg)、1/4(8.50 μg)、1/6(3.67 μg)、1/8(2.75 μg)、1/10(2.20 μg)。
1.3.3 包装细胞系比较 以1.3.1与1.3.2项的结果确定优化的质粒用量,实验方法同1.2,将BHK21-WI2作为病毒包装细胞系,BHK21-WI2细胞培养需7.5%CO2。
1.3.4 重组痘病毒vTF7-3作用时间比较 以
1.3.3的优化结果确定质粒用量及细胞系,实验方法同1.2。将重组痘病毒vTF7-3在作用1 h后移弃,更换为500 μL Opti-MEM培养基继续作用1.5和3 h。
1.3.5 转染试剂作用时间组 以1.3.3项优化结果确定质粒用量及细胞系,实验方法同1.2,将包装质粒转染时间在6 h基础上增加至8、10、12、14 h。
1.4 TCID50法测定包装病毒滴度
收集BHK21-WI2细胞,用含5%FBS的DMEM配制成2×105/mL的细胞悬液,以100 μL/孔接种至96孔细胞培养板,然后于37℃快速解冻1.3项各批次病毒,按10-1~10-10梯度稀释至无血清DMEM中,再添加至96孔细胞培养板,各梯度设置5个复孔,培养4~7 d。观察细胞形态及荧光变化,统计CPE孔数,按Karber法计算病毒滴度。
1.5 表达寨卡病毒E蛋白的rVSV(rVSVΔG-ZE)包装
在prVSVΔG-GFP骨架质粒基础上,构建缺失VSV的包膜糖蛋白(GP)基因而带有寨卡病毒包膜蛋白(E)基因的VSV重组质粒prVSVΔG-ZE,按1.2方法及1.3项优化条件进行病毒包装,并测定病毒滴度。
2 结果
2.1 细胞感染病毒的CPE及荧光观察
在病毒扩增阶段,BHK21-WI2细胞先转染pCAGGS-G质粒至逐渐融合、形成大小不等的合胞体(图1C),随后,细胞在感染rVSVΔG-GFP后,逐渐出现皱缩、变圆、脱落、漂浮等现象(图1A),在荧光显微镜下观察到明亮的绿色荧光(图1B)。由于BHK21-WI2细胞CPE变化较293T细胞显著,因此选择BHK21-WI2细胞用于观察。
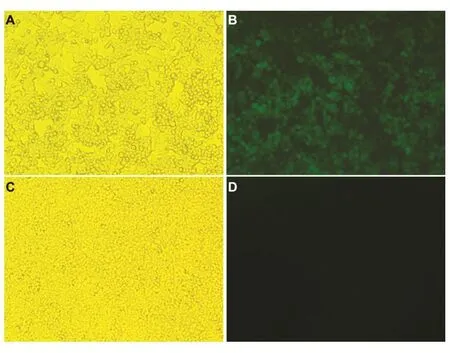
图1 病毒感染BHK21-WI2细胞后的CPE变化(100×)A:rVSVΔG-GFP感染细胞(光镜);B:rVSVΔG-GFP感染细胞(荧光);C:转染pCAGGS-G质粒细胞(光镜);D:转染pCAGGSG质粒细胞(荧光)
2.2 rVSVΔG-GFP滴度测定结果
不同条件包装病毒TCID50结果见表1。在辅助质粒单独递减时,病毒滴度降低;在骨架质粒及辅助质粒同时递减时,滴度降低,但趋势较小。不同细胞系包装rVSV时,293T细胞组包装病毒滴度远高于BHK21-WI2细胞组,延长vTF7-3作用时间或转染时间后,BHK21-WI2细胞系包装病毒滴度有一定提高。

表1 TCID50法测定rVSVΔG-GFP滴度
2.3 骨架质粒及辅助质粒用量的影响
本实验中rVSVΔG-GFP为复制缺陷型重组病毒,包装过程中还用到辅助质粒pBS-G、pBS-N、pBS-P、pBS-L,其比例为8∶3∶5∶1[17]。荧光观察结果显示,总质粒用量递减对荧光细胞数量影响不明显(图2A),但骨架质粒用量不变时,荧光细胞数量随辅助质粒用量递减而减少(图2B)。滴度测定结果显示总质粒用量为3.67~22 μg时可高效包装rVSVΔG-GFP,各组滴度变化差异不明显,总质粒用量低于3.67 μg时包装效率明显降低。骨架质粒为5 μg时,辅助质粒(pBS-N、pBS-P、pBS-L)的合适用量为3~9 μg,用量低于3 μg时包装效率极低。骨架质粒与辅助质粒用量较低时,或造成病毒基因组RNA及N、P、L蛋白表达量过低,而骨架质粒与辅助质粒比例相差过大时难以形成持续复制单位。这与已报道的rVSV载体非复制缺陷型疫苗包装条件较为一致[14],各文献中骨架质粒用量为2~6 μg,辅助质粒pBS-N、pBS-P、pBS-L比例近似为3∶5∶1,用量2~9 μg。

图2 不同质粒用量病毒感染BHK21-WI2细胞后CPE变化及荧光结果(100×)A1~A4:总质粒用量依次为1/2(11.00 μg)、1/4(8.50 μg)、1/6(3.67 μg)、1/8(2.75 μg);B1~B4:辅助质粒用量依次为(1/2(8.50 μg)、1/4(4.25 μg)、1/6(2.83 μg)、1/8(2.16 μg)
2.4 包装细胞系的影响
以2.3项结果确定优化的质粒用量为原骨架质粒和辅助质粒总量的1/2(11.00 μg),分别考察293T、BHK21-WI2细胞的包装效率。实验发现293T细胞系荧光细胞数量及包装稳定性显著优于BHK21-WI2细胞系(图3),且293T细胞包装能获得高滴度的病毒。293T细胞感染病毒时CPE不明显,生长较慢,而BHK21-WI2细胞包装效率低于同条件包装的293T细胞,生长快速,感染病毒后CPE明显[14,17]。另外,我们还尝试了BHK21和293T/Vero E6混合细胞进行病毒包装,293T/ Vero E6混合细胞体系包装效率高于BHK21-WI2细胞,BHK21-WI2细胞与BHK21细胞包装效率差异不明显。
2.5 重组痘病毒vTF7-3作用时间的影响
采用BHK21-WI2细胞包装时,荧光细胞数目较少且稳定性较差,我们期望通过延长vTF7-3的作用时间来提高其病毒包装效率。将vTF7-3的作用时间在1 h基础上分别延至1.5和3 h,发现均可成功包装病毒,且荧光细胞数量增多,滴度略有提高。结合前期实验发现vTF7-3以5~15 MOI感染细胞1 h或以5 MOI感染细胞1~4 h可成功包装病毒。当vTF7-3低于1 MOI感染1 h时,产生的T7RNA聚合酶量过少,导致后续包装效率低。当vTF7-3用量大于15 MOI或感染时间超过4 h时,包装效率变化不明显,甚至由于vTF7-3大量扩增抑制VSV包装。该结果与国外报道有部分差异,文献报道vTF7-3用量为5~10 MOI、感染时间45~60 min[18]。最近也有采用重组真核载体表达T7RNA聚合酶,而不用vTF7-3的报道[18]。
2.6 转染试剂作用时间的影响
同样为提高BHK21-WI2细胞包装效率,将转染试剂作用时间延长至8、10、12、14 h,其他条件不变。本实验中质粒转染细胞时间为6~14 h时包装效果较好,可见适当提高转染时间有利于提高包装效率。当转染时间小于6 h时,质粒进入细胞量过少,包装效率低。转染时间长于14 h时包装效率未能提高甚至出现降低,这可能是转染试剂作用时间过长导致细胞损伤,该结果比国外报道的转染细胞时间(4~7 h)长[18]。

图3 不同包装细胞系包装病毒感染BHK21-WI2细胞后的荧光观察结果(100×)A:293T包装细胞系组;B:BHK21-WI2包装细胞系组;C:转染pCAGGS-G质粒的对照细胞组
2.7 rVSVΔG-ZE组的包装结果
在rVSVΔG-GFP包装条件优化基础上,对rVSVΔG-ZE进行了病毒包装尝试,条件见表2。结果显示,在3种条件下实验组均出现明显的CPE,如图4所示,病毒均能包装且滴度较高。

表2 TCID50法测定rVSVΔG-ZE滴度
3 讨论
rVSV的包装包括以下几个主要方面:重组痘病毒vTF7-3感染细胞表达T7RNA聚合酶、转染的辅助质粒分别表达相应蛋白(核蛋白N、磷蛋白P、基质蛋白M、包膜蛋白G及大聚合酶蛋白L),骨架质粒转录生成病毒基因组RNA,核蛋白N与基因组RNA组装形成持续复制单位[17,19]。我们基于这些方面考察重组痘病毒vTF7-3的作用时间、总质粒用量与辅助质粒用量、质粒转染时间以及细胞系对rVSV包装效率的影响。此外,表达的外源蛋白也是影响rVSV包装的重要因素,如表达的外源蛋白有类似VSV野生型包膜GP蛋白的介导病毒与膜融合的性质,则包装效率高、扩增能力强,而类似GFP自身无包膜蛋白性质,则需要借助辅助质粒pBS-G包装形成复制缺陷性重组病毒,包装效率、扩增能力较低。VSV病毒包装是一个多条件影响的过程,单一适宜条件或许能得到病毒,但综合优化各个条件才能获得稳定高效的高滴度病毒株。

图4 rVSVΔG-ZE组感染BHK21-WI2细胞结果(100×)A:1/2(11.00 μg)全质粒293T细胞包装组;B:1/2(8.50 μg)辅助质粒293T细胞包装组;C:1/2(11.00 μg)质粒vTF7-3作用4 h转染试剂作用14 h于BHK21-WI2细胞包装组;D:转染pCAGGS-G质粒的对照细胞组
[1] van den Pol A N,Mao G,Chattopadhyay A,et al. Chikungunya,influenza,Nipah,and Semliki forest chi⁃meric viruses with vesicular stomatitis virus:actions in the brain[J].J Vriol,2017,91(6):pii:e02154-16.
[2] Dolzhikova I V,Zubkova O V,Tukhvatulin A I,et al.Safety and immunogenicity of Gam Evac-Combi,a heterologous VSV- and Ad5-vectored Ebola vaccine: an open phase I/II trial in healthy adults in Russia [J].Hum Vaccin Immunother,2017,13(3):13-620.
[3] Huttner A,escure C,Grillet S,et al.A dose-depen⁃dent plasma signature of the safety and immunogenici⁃ty of the rVSV-Ebola vaccine in Europe and Africa [J].Sci Transl Med,2017,9(385):pii:eaaj1701.
[4] Wang Y,Liu Z,Dai Q.A highly immunogenic frag⁃ment derived from Zaire Ebola virus glycoprotein elic⁃its effective neutralizing antibody[J].Virus Res,2014, 189:254-261.
[5] Lee S S,Phy K,Peden K,et al.Development of a mireplication-competent vesicular stomatitis hybrid vi⁃rus and a quantitative PCR readout[J].Vaccine,2017, 35(41):5481-5486.
[6]李拓,刘珠果,张跃,等.马尔堡病毒GP蛋白抗原性片段筛选及免疫性研究[J].军事医学,2017,41(6):440-444.
[7] 李拓,刘珠果,戴秋云.马尔堡病毒疫苗研究进展[J].军事医学,2016,40(3):261-264.
[8] Liu R,Wang J,Shao Y,et al.A recombinant VSV-vectored MERS-CoV vaccine induces neutralizing anti⁃body and T cell responses in rhesus monkeys after single dose immunization[J].Antiviral Res,2018,150: 30-38.
[9] Gsell P S,Camacho A,Kucharski A J,et al.Ring vaccination with rVSV-ZEBOV underexpanded ac⁃cess in response to an outbreak of Ebola virus dis⁃ease in Guinea,2016:an operational and vaccine safe⁃ty report[J].Lancet Infect Dis,2017,17(12):1276-1284.
[10]Lawson N D,Stillman E A,Whitt M A,et al.Recom⁃binant vesicular stomatitis viruses from DNA[J].Proc Natl Acad Sci USA,1995,92(10):4477-4481.
[11]Pattnaik A K,Ball L A,Le Grone A W,et al.Infec⁃tious defective interfering particles of VSV from tran⁃scriptsofa cDNA clone[J].Cell,1992,69(6):1011-1020.
[12]Whelan S P,Ball L A,Barr J N,et al.Efficient re⁃covery of infectious vesicular stomatitis virus entirely from cDNA clones[J].Proc Natl Acad Sci USA,1995, 92(18):8388-8392.
[13]Takada A,Robison C,Goto H.A system for function⁃al analysis of Ebola virus glycoprotein[J].Proc Natl Acad Sci USA,1997,94(26):14764-14769.
[14]Garbutt M,Liebscher R,Wahl-Jensen V,et al.Proper⁃ties of replication-competent vesicular stomatitis virus vectors expressing glycoproteins of filoviruses and are⁃naviruses[J].J Virol,2004,78(10):5458-5465.
[15]Wong G,Qiu X.Designing efficacious vesicular stoma⁃titis virus-vectored vaccines againstEbola virus[J]. Methods Mol Biol,2016,1403:245-257.
[16]邵钰,王金良,刘任强,等.埃博拉和马尔堡病毒囊膜糖蛋白嵌合型重组水泡性口炎病毒构建及免疫原性研究[J].中国预防兽医学报,2016,38(6):438-442.
[17]Whitt M A,Geisbert T W,Mire C E.Single-vector, single-injection recombinant vesicular stomatitis virus vaccines against high-containment viruses[J].Methods Mol Biol,2016,1403:295-311.
[18]Wong G,Audet J,Fernando L,et al.Immunization with vesicular stomatitis virus vaccine expressing the Ebola glycoprotein provides sustained long-term protec⁃tion in rodents[J].Vaccine,2014,32(43):5722-5729.
[19]Whitt M A.Generation of VSV pseudotypes using re⁃combinant ΔG-VSV for studies on virus entry,identifi⁃cation of entry inhibitors,and immune responses to vaccines[J].J Virol Methods,2010,169(2):365-374.
猜你喜欢
电子乐园·上旬刊(2022年5期)2022-04-09
昆明医科大学学报(2022年2期)2022-03-29
华侨大学学报(自然科学版)(2021年4期)2021-07-30
中国新技术新产品(2020年5期)2020-05-06
医学研究杂志(2015年4期)2015-06-10
中国医药导报(2015年26期)2015-02-28
现代检验医学杂志(2015年2期)2015-02-06
中国煤层气(2014年3期)2014-08-07
中华介入放射学电子杂志(2014年1期)2014-02-02
中国中西医结合皮肤性病学杂志(2013年4期)2013-11-01
