
藏东南兰科植物多样性及其沿海拔梯度的分布格局
2018-04-08 05:07王喜龙土艳丽文雪梅朱荣杰段元文
中南林业科技大学学报 2018年12期
王喜龙,土艳丽,文雪梅,朱荣杰,段元文
(1.西藏自治区高原生物研究所,西藏 拉萨 850001;2.西藏自治区农牧科学院蔬菜研究所,西藏 拉萨 850032;3.中国科学院 昆明植物研究所,云南 昆明 650201)
兰科是单子叶植物最大的科和被子植物中的第二大科,全世界约有800属25 000种[1],该科植物在植物界的系统演化上属于最进化、最高级的类群,主要表现在丰富的生活型和高度多样性的花特征[2]。根据Flora of China的记录,中国的兰科植物有194属1 388种[3],主要分布于热带和亚热带地区,少数种类分布在温带地区。虽然我国典型热带地区所占比例面积不大,也不是兰科植物种类最丰富的地区,但由于中国跨越热带、亚热带和温带3个气候带以及具有复杂的地理环境,使得中国不仅具有各个气候带的兰科植物区系和生态类型,并且还保留了许多原始类群[4-6]。
青藏高原是全世界面积最大的高原,较大的纬度跨度使得该地区具有较多的低纬度带,高原多样的地形条件和气候类型孕育了这一地区低纬度带丰富的生物多样性,是全球34个生物多样性热点地区的重要组成部分[7]。由于跨越多个不同的气候和植被带,使得该地区的气候类型复杂而且垂直变化大。特殊的地理条件和气候类型使得藏东南地区拥有世界上独一无二的兰科植物区系[8]。1987年出版的《西藏植物志》第五卷记录了西藏兰科植物有64属191种及2变种[9],这些基础资料对于了解西藏兰科植物资源、分布和生存环境为我们提供了最为宝贵的资料。近年来,随着西藏经济社会的快速发展,一些较为偏远的地区,如墨脱等地逐渐通车,为植物学工作者提供了便利的研究条件,并且陆续有一些新种和新纪录种发现和报道[10-20]。
目前,生物多样性沿海拔梯度的变化规律是生物多样性研究热点之一,生物多样性随着海拔的升高会出现较大的变化[21]。研究物种多样性的梯度格局以及控制这些格局的生态因子也是保护生物学研究的重要基础,群落中物种多样性对环境因子的响应不同,不同区系成分的垂直分布格局也有所不同[22]。例如,王国宏(2002)[23]对祁连山北坡中段植物群落物种多样性垂直分布格局进行了初步研究,结果表明植物群落草本层和灌木层物种丰富度和多样性在海拔梯度上呈单峰曲线变化趋势,而乔木层的物种丰富度和多样性的变化不大。而王志恒等(2004)[24]对高黎贡山植物沿海拔梯度的研究,发现全部种子植物和不同生活型植物物种丰富度随着海拔的升高呈现先增加后减小的趋势,最大值出现在海拔1 500 ~2 000 m的范围;物种密度与海拔也呈现单峰曲线关系。物种丰富度和物种密度分布格局的形成主要受不同海拔梯度的水、热状况组合差异以及物种分布的边界影响。
喜马拉雅山脉阻挡了夏季印度洋水汽进入到青藏高原,使得喜马拉雅山的南坡和北坡的气候条件(特别是水热条件)出现了巨大的差异。已有的研究表明,北侧兰科植物随海拔的升高物种多样性降低[26-27]。而喜马拉雅南侧兰科植物是否存在同样的变化趋势仍然是空白。因此,本研究通过野外实地调查和查阅相关资料基础上,分析了藏东南兰科植物多样性、生活型及其沿海拔梯度分布格局,以其为西藏野生兰科植物资源的保护和研究提供基础数据。通过对藏东南兰科植物多样性随海拔梯度变化格局的研究对于揭示兰科植物多样性的环境梯度变化规律具有重要意义。
1 研究方法
1.1 研究区域
藏东南地区位于西藏东南部雅鲁藏布江中下游,复杂的地形和巨大的海拔高差使得该地区拥有多样的植被类型和丰富的区系成分类型。因此,藏东南地区成为生物多样性和植物区系研究的理想区域。本研究“藏东南地区”特指即林芝市辖区的工布江达县、米林县、墨脱县、波密县、察隅县、朗县和新设立的巴宜区等7县区。尤其是墨脱县域内地势北高南低,海拔在200 ~7 787 m,平均海拔1 200 m,南迦巴瓦峰和加拉白垒峰是东喜马拉雅山脉最高的两座山峰,地势由北向南急剧下降,北部高达7 756 m的南迦巴瓦峰,南缘巴昔卡海拔仅155 m,短距离内相对高差7 000 m,形成了世界上最深最长最险峻的峡谷—雅鲁藏布大峡谷。受印度洋暖湿气流的影响,该地区植物种类繁多、植被结构复杂,垂直带谱明显,在40 km的水平距离内,从高山寒带植物到热带植物几乎都能生长,原始森林类型众多,有常绿阔叶林、针阔叶混交林、暗针叶林等。本研究重点对墨脱地区开展了实地科学考察,详细记录了以上各县兰科植物分布等信息,并采集相关标本。标本信息的获取主要以墨脱、察隅、米林、波密和巴宜区为主。
1.2 数据获取及整理
本研究以在2014年—2017年先后开展了3次野外调查并采集了大量的标本,同时结合《西藏植物志》、中国植物志(西藏部分)、《中国高等植物彩色图鉴》第9卷以及标本信息(主要来自中国数字植物标本馆(CVH)(http://www.cvh.ac.cn),以及新发表的新种、新纪录种等对资料进行整理。对所有记录的兰科植物分布按照500 m的间隔划分了9个不同海拔梯度带,分别为0~1 100 m、1 100~1 500 m、1 500~2 000 m、2 000~2 500 m、2 500~3 000 m、3 000~3 500 m、3 500~4 000 m、4 000~4 600 m、>4 600 m,统计各海拔段内兰科植物种数、生活型、相似性和区系成分等。兰科植物区系成分分析主要依据中国种子植物属的分布区类型进行划分[28]。
1.3 系统发育树构建
在确定物种名录后,以种子植物系统发育关系[29]为主体,利用Phylomatic在线软件构建基于APGⅢ分类系统的西藏兰科植物的系统发育树[30]。
1.4 物种多样性和系统发育多样性分析
不同海拔梯度带的物种多样性用该海拔带兰科植物的物种数量表示,各海拔梯度带间物种相似性分析采用Sørenson系数法[31]表示,具体计算公式为Cs=2c/(a+b),其中a、b分别为任意两个海拔带的兰科植物种数,而c为两个海拔带之间共有的兰科植物种数。
系统发育多样性分析用Faith’s PD系数[32]和系统发育beta多样性PhyloSor系数表示[33]。PD表示各海拔段所有物种的进化历史长度,PD值越大表明进化历史越长。具体的计算公式为PhyloSorij=2BLij/(BLi+BLj),其中PhyloSorij表示不同海拔带i和j的系统发育的β多样性,BLij表示两个海拔段共有物种的枝长和,BLi和BLj分别表示不同海拔段i和j中所含的物种的枝长和,PhyloSor比值越大说明两个海拔段物种相似度越高。
净相关系数(Net relatedness index,NRI)来衡量不同海拔梯度带兰科物种系统发育结构[30],当NRI> 0时表示系统发育结构为聚集,NRI值越高则亲缘关系越近;当NRI< 0时表示系统发育结构为分散;NRI值接近0则表明物种在系统发育上是随机的。所有上述系数的计算均在R软件(R development Core Team, 2016)中对应的程序包完成。
2 结果与分析
2.1 野生兰科植物物种多样性
共记录了西藏野生兰科植物89属336种,分别占中国兰科植物的45.9%和26.4%,表明西藏野生兰科植物的种类多样性极为丰富。其中,达到或超过20个物种的属有3个,分别是石豆兰属Bulbophyllum有20种,占总种数的5.5%;角盘兰属Herminium有20种,占总种数的5.5%;羊耳蒜属Liparis有21种,占总种数的5.7%。舌唇兰属Platanthera和虾脊兰属Calanthe各有18种和17种;兰属Cymbidium、杓兰属Cypripedium、石斛属Dendrobium、斑叶兰属Goodyera、玉凤花属Habenaria各有12种;贝母兰属Coelogyne有10种。以上超过10个种的有12属,共计178种,占总种数的48.6%。矮柱兰属Thelasis、白及属Bletilla等39属仅记录到1种,占总属数的43.8%。
2.2 兰科植物沿海拔梯度分布格局
对兰科植物海拔梯度的分析结果表明,西藏兰科植物的海拔分布呈“中间高度膨胀型”分布趋势。兰科植物主要分布在1 500~2 000 m、2 000~2 500 m、2 500~3 000 m、3 000~3 500 m等4个海拔段内,各有132种,159种,130种和95种(见表1),在海拔2 000 ~ 2 500 m海拔段分布最多。而分布最少的是4 600 m海拔段,由于海拔较高,仅有生长有2种地生兰科植物,分别为斑唇盔花兰Galearis wardii、剑唇兜蕊兰Herminium pugioniforme。

表1 兰科植物不同海拔梯度的物种多样性与系统发育多样性比较Table1 Comparison of species diversity and Phylogenetic diversity of different elevational blets for Orhideceae in Tibet
不同海拔段兰科植物系统发育多样性与物种丰富度呈显著正相关关系(R2=0.995,P<0.001),物种丰富度越高物种系统发育多样性也随之升高。西藏兰科植物不同海拔系统发育相似性与物种相似性也呈显著正相关关系(R2=0.994,P< 0.001)。在第2 500~3 000 m海拔段系统发育多样性系数最高,达到10 480.09,与物种丰富度系数一致,都随着海拔的升高而逐步升高,在2 000~2 500 m海拔段达到一个峰值,然后逐渐降低。各海拔梯度兰科植物的系统发育结构除了在4 600 m以上呈离散性外其余均呈聚集型分布。在3 500~4 000 m海拔段NRI系数最高(4.85),说明亲缘关系越近。在4 600 m海拔以上NRI< 0,呈谱系离散状态,说明在4 600 m海拔以上,兰科植物的亲缘关系越远。
2.3 兰科植物生活型构成的垂直分布格局
西藏野生兰科植物生活型有地生兰187种、附生兰136种、菌类寄生兰13种,分别占总种数的55.7%、40.5和3.6%(图1)。除4 000 m以上海拔外,其余各种生活型兰科植物在各海拔都有分布。地生兰主要分布在2 000~2 500 m、2 500~3 000 m、3 000~3 500 m的海拔段内,种数分别为85、85和81,且在各海拔段都有分布;附生兰除了在海拔4 600 m以上无分布外,其余都有分布,且在第1 500~2 000 m海拔段最高(82种);腐生兰除了在4 000 m以上海拔无分布外,在其余各海拔段都有分布,且在2 000~2 500 m海拔段分布最高(6种)。地生兰的物种数都随着海拔的上升而升高然后下降,呈中间高两边低的趋势。附生兰主要分布在低海拔,随着海拔段升高逐步减少。腐生兰在高于海拔4 000 m没有分布。

图1 各生活型野生兰科植物在不同海拔段分布数量Fig.1 Species number of orchid life forms in different elevational belts of Tibet
2.4 兰科植物不同海拔段兰科植物物种相似性分析
由表2和表3可知,不同海拔段之间兰科植物物种和系统发育相似性系数有一定的差异。不同海拔段的兰科植物平均物种组成相似性(Sørenson系数)为0.224,其中在Ⅳ与Ⅴ和Ⅴ与Ⅵ最高分别为0.65。在不同海拔段(n= 36)的兰科植物平均系统发育相似性(PhyloSor系数)为0.280,其中在Ⅴ与Ⅵ两组海拔段的物种相似度最高,PhyloSor系数均为0.69。Ⅳ与Ⅴ和Ⅵ与Ⅶ两组海拔段一致,分别为0.68。而且,随着海拔差加大,相似性系数也随之降低,甚至在4 600 m以上海拔与其他海拔段相比较,相似系数几乎为0。Sørenson系数和PhyloSor系数在相邻海拔段基本一致,在统计学上无显著差异(P> 0.05),都在Ⅴ与Ⅵ海拔段达到最高,同时也是物种分布最为丰富的海拔段。
2.5 兰科植物不同海拔段兰科植物区系分析
在所有记录的336种兰科植物中,热带亚洲成分、中国-喜马拉雅成分和中国特有成分占绝对优势,三者之和占总种数的74.3%(表4)。其中热带亚洲成分物种数为49属120种,主要分布在Ⅰ-Ⅴ的海拔段内,且在第Ⅲ海拔段分布种数最多,如疏花石斛Dendrobium henryi、斑叶兰Goodyera schlechtendaliana、锥囊坛花兰Acanthephippium striatum;其次为中国-喜马拉雅成分有42属96种,如藏南角盘兰Herminium clavigerum、墨脱石豆兰Bulbophyllum eublepharum、通麦虾脊兰Calanthe griffithii等,主要分布在第Ⅳ和Ⅴ海拔段,种数分别为49和40种;中国特有的物种有25属56种,如:西藏玉凤花Habenaria tibetica、波密杓兰Cypripedium ludlowii、暖地杓兰Cypripedium subtropicum、清涌兜兰Paphiopedilum qingyongii等物种,主要分布在第Ⅵ海拔段,种数达30种。其中,属于旧世界热带成分、热带印度至华南成分、温带亚洲成分、中亚成分和西亚至喜马拉雅和西藏等区系成分各仅有1种分布,分别为丛生羊耳蒜Liparis cespitosa、鹤顶兰Phaius tancarvilleae、裂瓣角盘兰Herminium alaschanicum、掌裂兰Dactylorhiza hatagirea和卵叶火烧兰Epipactis royleana。而落地金钱Habenaria aitchisonii和小花舌唇兰Platanthera minutiflora等2种兰科植物属于中亚至喜马拉雅和我国西南成分。
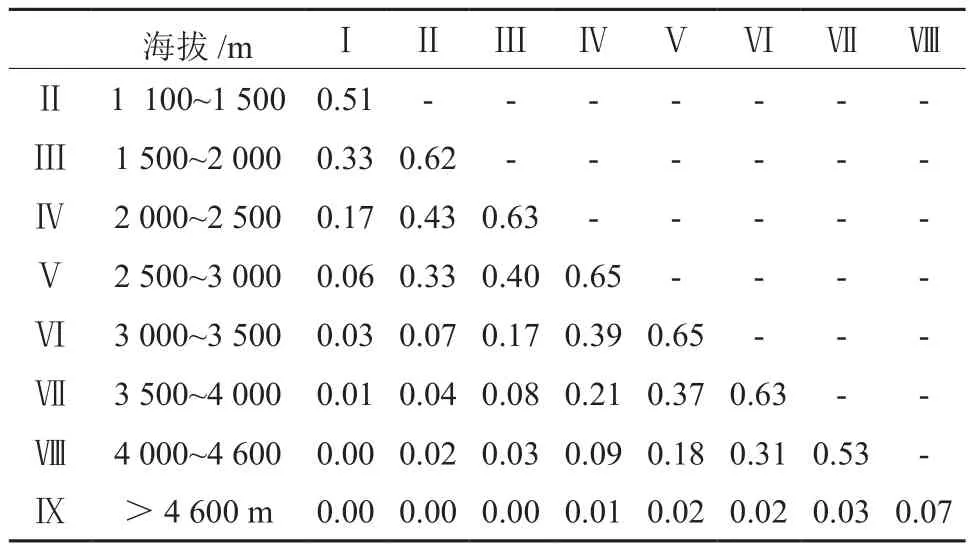
表2 藏东南不同海拔段兰科植物物种相似性系数Table2 Similarity coefficient of orchid plant species at nine elevation belts of Tibet
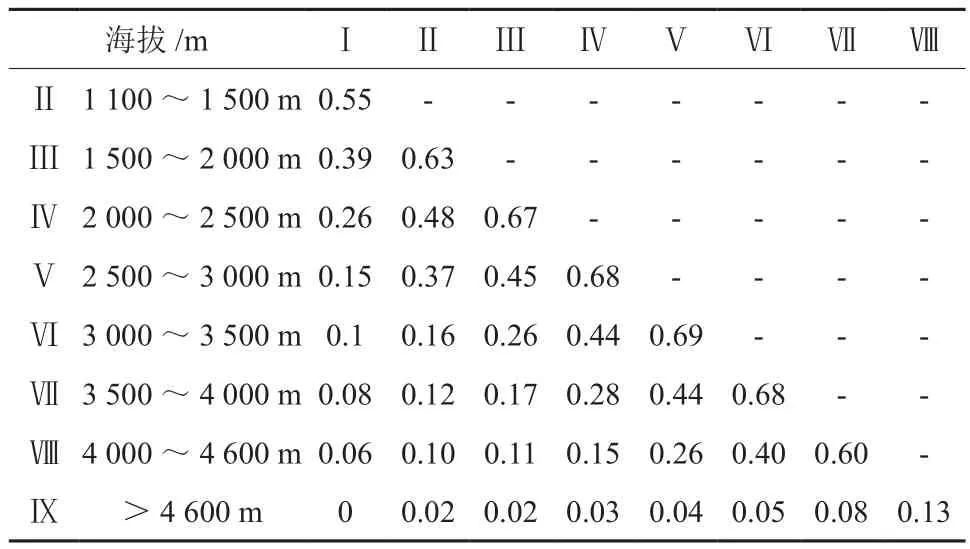
表3 藏东南不同海拔段兰科植物系统发育相似性系数Table3 Similarity coefficient of orchid plant species at nine elevation belts of Tibet
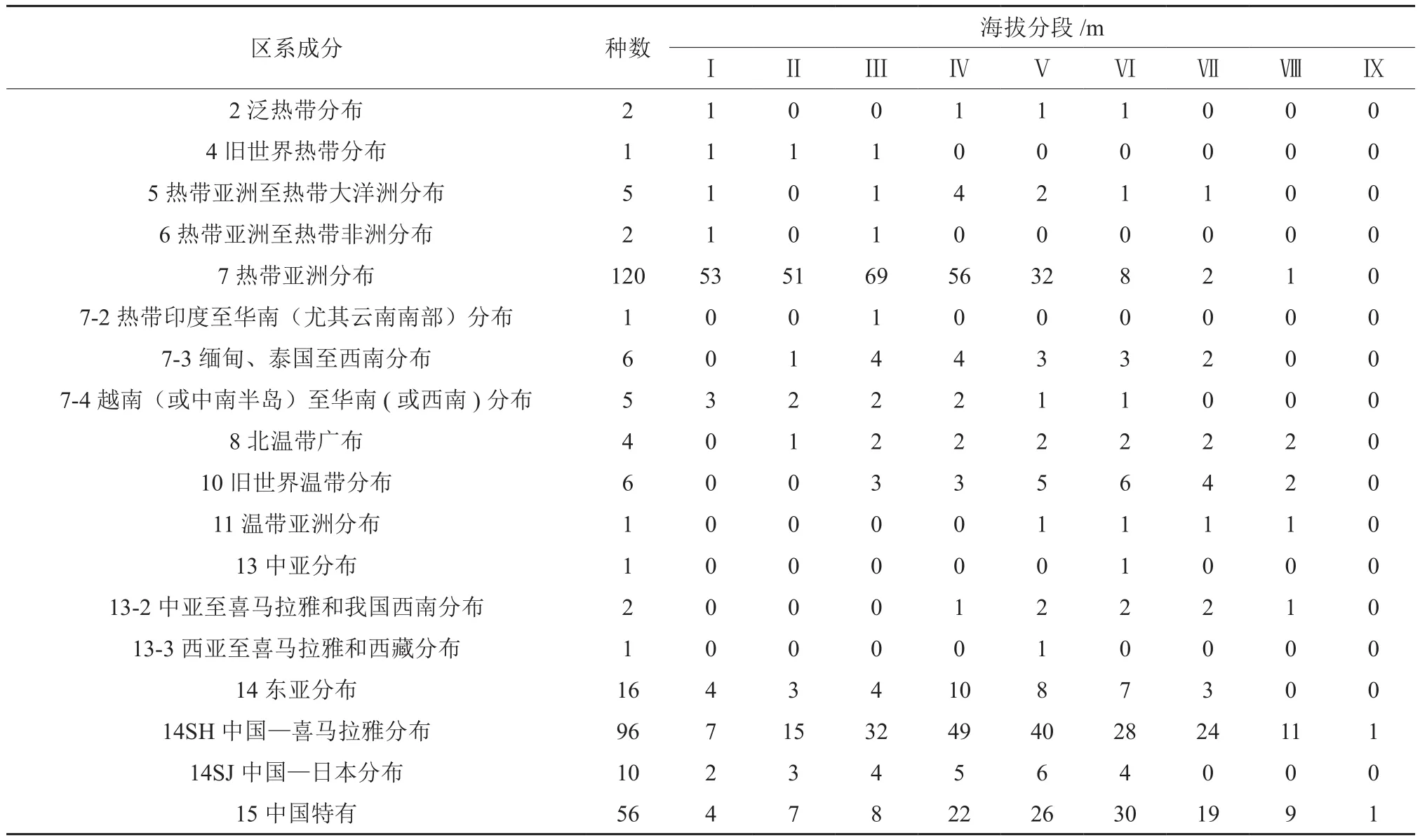
表4 不同海拔兰科植物区系分析Table4 Floristic components of orchids at different elevation belts in Tibet
3 结论与讨论
通过野外调查和相关文献资料整理,共记录了西藏野生兰科植物89属336种。西藏野生兰科植物随着海拔呈现“中间高度膨胀型”分布趋势。其生活型也呈多样化,不论是那种生活型的兰科植物,物种数都随着海拔的上升而升高然后下降,也呈“中间高度膨胀型”趋势。相邻海拔段兰科植物相似性系数较高,随着海拔差加大,相似性系数也随之降低。西藏的兰科植物总体分布主要集中在1 500~3 500 m海拔段,在海拔2 000~2 500 m分布最多,而分布最少的是4 600 m以上的海拔,仅有生长有2种兰科植物,分别为斑唇盔花兰Galearis wardii、剑唇兜蕊兰Herminium pugioniforme。通过群落系统发育的研究,结果也表明,分布在藏东南的野生兰科植物在不同海拔系统发育相似性与物种相似性呈显著正相关关系,群落多样性系数随海拔升高而逐渐降低的趋势。群落系统发育结构沿海拔梯度发生显著变化,说明非随机过程在生态群落的构建过程中起着非常重要的作用,同时揭示了藏东南野生兰科植物的系统发育结构随海拔的升高而呈现出由聚集(海拔4 600 m以下)走向发散(海拔4 600 m以上)的变化规律。参照吴征镒对中国种子植物属的分布区类型划分,将兰科植物划分为18个类型,其中分布最多的是热带亚洲分布,有120种兰科植物;其次为中国—喜马拉雅分布96种,中国特有4种。在东亚分布16种,中国—日本分布10种。兰科区系成分中热带亚洲成分、中国—喜马拉雅成分和中国特有成分占绝对优势,其中热带亚洲成分物种数>中国-喜马拉雅成分>中国特有成分。
兰科植物作为西藏植物区系的重要组成部分,也是濒危物种保护的“旗舰类群”[2]。西藏兰科植物多样性丰富度仅次于云南、广东、台湾和四川等省市[8]。尤其是位于西藏南部和东南部的墨脱、波密和察隅等地,受西南季风的影响,气候温暖湿润,为兰科植物的生长提供了得天独厚的自然环境,是兰科植物集中分布地方,本研究记录到的兰科植物在《西藏植物志》的基础上增加了25属143种(变种)。随着西藏道路等条件的不断改善,一些兰科新种和新纪录种的发现也在不断的增加,极大的丰富了西藏兰科植物的多样性,如无叶沼兰Crepidium aphyllu、矮生白点兰Thrixsperm pygmaeum、西藏槌柱兰Malleolla tibetica、小馥兰Phreatia elegans、冷兰Herminium humidicola、无叶美冠兰Eulophia zollingeri和大理铠兰Corybas taliensis、杉林溪铠兰Corybas himalaicus、山珊瑚Galeola faberi。后来也有研究人员对西藏部分区域开展了兰科植物调查,如:石松林等(2012)[34]对珠穆朗玛峰国家级自然保护区兰科植物多样性进行了研究,调查有兰科植物35属63种,主要以地生兰为主。林玲等(2013)[35]对林芝色季拉山兰科植物的分布进行了研究,共记录了兰科植物35属67种。
植物物种多样性随海拔梯度的变化主要有5种模式[25]:(1)负相关,即随海拔的升高物种多样性随之降低;(2)中间高度膨胀,即物种多样性在中等海拔最高;(3)在中等海拔物种多样性最低;(4)正相关,即随着海拔的升高物种多样性增加;(5)无相关性,即物种与海拔变化无规律。藏东南的野生兰科植物随着海拔呈现“中间高度膨胀型”分布趋势,这与郎楷永(1980)[8]以及田怀珍等(2008)[36]、杨正斌等(2014)[37]、韩宇等(2015)[38]的研究结果一致。西藏的兰科植物主要分布在中海拔阶段,因为中海拔的环境梯度最宽,从而植物物种多样性最多,中海拔段有着适宜各种生活型兰科植物的生长环境,所以不同生活型的兰科物种多样性均在此达到最大值[37]。而分布最少的是4 600 m以上的海拔,这可能与高原特殊的地理环境有很大的关系,在4 600 m以上的海拔主要为高山冰缘植被为主,位于高山草甸之上,永久雪线之下,强烈的太阳辐射和大的昼夜温差使得兰科植物的生长繁育受到限制。我们也通过群落系统发育的研究也均说明群落系统发育结构沿海拔梯度发生显著变化,说明非随机过程在生态群落的构建过程中起着非常重要的作用,这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据[39]。利用群落系统发育方法来理解和探讨群落装配的过程,对全球气候变化下生物多样性的保护与维持有着非常重要的意义。
郎楷永先生最早对西藏兰科植物地理和区系进行了系统的研究,兰科植物主要分布于藏东南,尤以墨脱和察隅两地区最多[8],而且中国特有的兰科植物达61种。尤其是位于藏东南的墨脱地区正处于横断山区系向东喜马拉雅区系,是生物多样性热点地区,孕育了丰富的附生兰科植物,有“植被类型天然博物馆”之称。虽然近年来藏东南的交通条件大为改善,西藏自治区高原生物研究所也组织开展了多次动植物考察,但有些比较偏僻的地区任然受到人力、物力和自然条件的限制,在考察时没有覆盖到,尤其在雅鲁藏布江大峡谷国家级自然保护区覆盖范围内道路极为艰险,考察物资全靠人背马拖,使得考察无法深入完全覆盖整个研究区域,有些兰科植物可能被遗漏而未被记录到。作为生物多样性热点地区,还需要和保护区管理局以及相关研究单位联合开展综合科学考察,对保护区本地的资源进行深入考察,为保护区的管理提供坚实的基础数据[40],特别是需要对一些濒危、特有植物进一步深入联合开展综合科学考察,为将来西藏生物多样性保护提供支撑。
猜你喜欢
数学物理学报(2022年5期)2022-10-09
河北画报(2020年8期)2020-10-27
陕西农业科学(2019年3期)2019-04-09
浙江林业(2018年8期)2018-05-14
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
环球市场信息导报(2017年1期)2017-04-08
现代农业科技(2016年22期)2017-03-24
学周刊·下旬刊(2015年10期)2015-07-22
俄罗斯问题研究(2013年1期)2013-03-11
