
太空诱变对紫色马铃薯种子及幼苗性状的影响研究
2018-03-06 03:48张桂芝刘淑娜于高波金光辉姜丽丽李德泽田丽美高幼华
中国林副特产 2018年1期
张桂芝,刘淑娜,于高波,金光辉,姜丽丽,李德泽,田丽美,高幼华
(黑龙江八一农垦大学农学院,黑龙江 大庆 163319)
彩色马铃薯属双子叶植物纲茄科多年生草本植物[1],原产南美秘鲁等地,国内近年已有种植,且随着人们对食品多样性和安全健康食品需求增加,彩色马铃薯深受大众喜爱,而紫色马铃薯是彩色马铃薯中重要一类,花青素含量高且营养丰富[2]。但目前生产上的紫色马铃薯栽培品种存在品种少、产量低的问题。利用返回式卫星搭载进行植物太空诱变育种已成为我国相对独立且有效的育种手段[3],水稻、大豆、棉花、青椒、黄瓜等几十种作物千余个品种应用太空诱变技术成功选育出513个作物品种和一些对农作物品质和产量有重要影响的罕见突变体[4]。马铃薯遗传较复杂,与小麦等主粮作物相比,辐射诱变育种进展较缓慢,获得成就少[5],如鲁马铃薯 2 号是经Co60的照射育成[6],黑美人是采用航天育种技术育成[7],国内对马铃薯辐射诱变研究多集中在薯块、愈伤组织、试管苗上[8-10],对于航天诱变马铃薯实生种子和幼苗的研究少有报道。本试验是利用“实践十号”进行了紫色马铃薯实生种子搭载,初步研究微重力和太空强辐射等因素对紫色马铃薯种子及幼苗期生理性状的影响,以期为紫色马铃薯种质改良和品种选育提供参考。
1 材料与方法
1.1 试验地点
试验于2016年12月至2017年6月期间在黑龙江八一农垦大学马铃薯种质资源实验室和育种基地日光温室内完成。
1.2 材料
试验材料为3个紫色马铃薯品系,分别为MD07-15、ZL02-3、ND13-4,由黑龙江八一农垦大学马铃薯研究所提供,3个品系实生种子经“实践十号”返回式科学实验卫星搭载12d返回地面,以未经搭载的地面种子为对照,进行发芽试验和幼苗期性状鉴定试验。
1.3 方法
1.3.1 种子发芽试验
每个品系搭载和未搭载种子分别取150粒,设三次重复,放入培养皿中浸种24h,放置在20℃无菌培养室中进行发芽培养。每天记录种子萌发数,将正常幼苗和不正常幼苗分别记录,死种子和霉烂种子及时检出并记录,直至无萌发种子出现为止,测定各诱变材料实生种子发芽势、发芽率、发芽指数等。
1.3.2 幼苗性状地面鉴定试验
取各诱变品系200粒,以未搭载种子为对照(CK),于2016年12月3日在育种基地日光温室内进行播种,设4次重复,每个品种播种4个育苗箱,育苗箱规格长1m×宽0.5m×高0.15m,以0.1m×0.1m距离点播,育苗土为细沙∶黑壤土∶腐熟菌渣肥=1∶5∶0.1,温室正常管理。出苗后及时观察并记录田间出苗率、成苗率,待幼苗长至30d测量并计算平均株高、茎粗,记录叶片数,观察诱变种质在株高、株型、茎色、叶色、叶缘形状等农艺性状,与对照进行比较,在田间表现有明显差异的植株,定为假定变异株。
茎粗与株高均采用门福义等的方法测定[11]。株高是用直尺测量生长点至茎基的高度,茎粗是用游标卡尺去测量茎基的最粗处。
1.3.3 幼苗叶片保护酶测定方法
待幼苗长至30d时随机取样并测定诱变品系搭载前后幼苗叶片相关保护酶活性,包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性。采用紫外吸收法进行幼苗叶片CAT活性的测定[12-13],采用愈创木酚法进行POD活性的测定[13],采用氮蓝四唑(NBT)光还原法进行SOD活性的测定[14]。
2 结果与分析
2.1 太空诱变对紫色马铃薯种子的影响
2.1.1 太空诱变对紫色马铃薯种子发芽势的影响
由表1可以看出,在经过航天诱变处理之后,不同的紫色马铃薯品系发芽势比对照低,且降低幅度各品系间有较大差异,达9.3%。其中MD07-15经航天诱变后发芽势变化最大,比对照低20.1%,其次是ZL02-3和ND13-4,分别是14.8%和10.8%。诱变材料种子发芽势下降的原因可能是太空微重力和强幅射对种子造成一定的生理损伤,致使诱变后的种子发芽较普通种子缓慢。
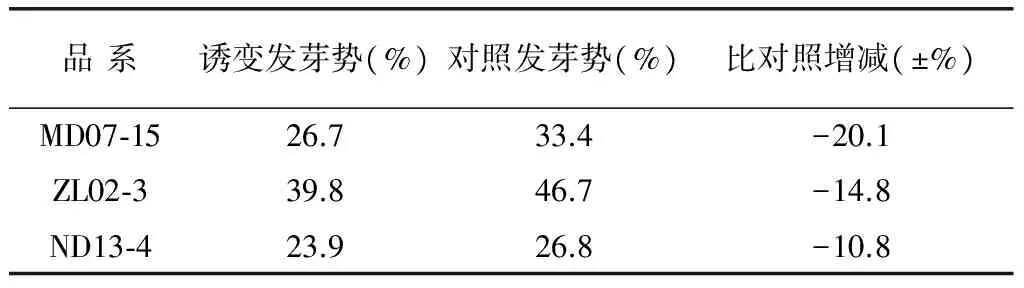
表1 诱变品系种子发芽势比较
2.1.2 太空诱变对紫色马铃薯种子发芽率的影响
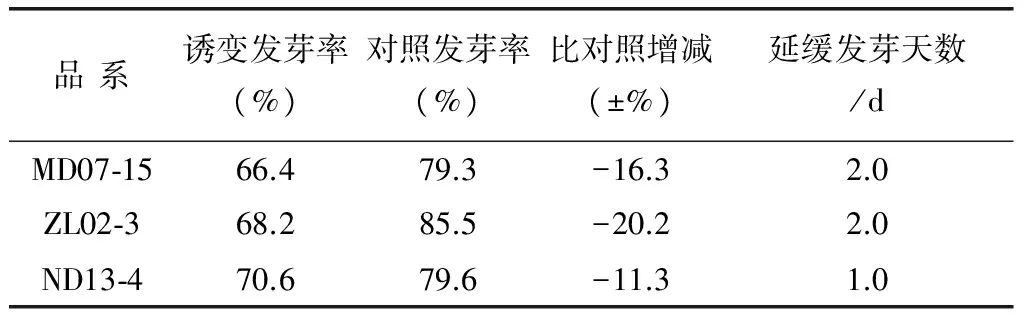
表2 诱变品系种子发芽率比较
各诱变材料种子发芽率变化情况如表2所示,经太空诱变处理之后,各品系发芽天数延迟,且发芽率均比对照低10%以上,且不同品系间存在差异。其中ZL02-3的下降幅度最大,ND13-4降低幅度最小。经太空强幅射后,不同紫色马铃薯品系之间发芽率的差异表现不同的,可能原因有不同品系自身的遗传物质不同,还有不同品系对环境的反应敏感性和应激反应不同,诱变效率就会有差异。
2.1.3 太空诱变对紫色马铃薯种子成苗能力的影响
从表3可以看出,3个诱变品系的出苗率和成苗率均较对照有所下降,各品系之间的下降幅度存在很大差异,差异程度达27%。其中ZL02-3下降幅度最大,达27.0%以上,而ND13-4下降幅度最小,其成苗率仅下降0.9%,MD07-15则居中。在成苗能力方面,各品系之间也存在差异性,ZL02-3CK的出苗率和成苗率都高于其它两品种,但诱变处理后其出苗率和成苗率都降为最低,而ND13-4的出苗率和成苗率则最高,这说明不同品系对诱变环境效应有较大差异。
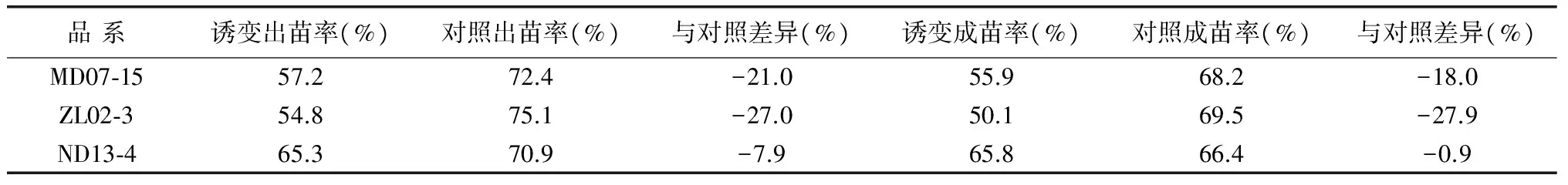
表3 诱变品系种子成苗能力分析
2.2 太空诱变对紫色马铃薯幼苗农艺性状的影响
通过搭载前后各诱变材料群体观察取样测定(表4),经过航天诱变处理后,各品系幼苗期生长势受到抑制,生长迟缓,矮化,平均株高和平均叶片数都低于对照,平均株高降幅在2.33%~8.62%, MD07-15和ZL02-3诱变后生长迟缓,平均叶片数都少于对照。本试验结果表明,航天诱变后3个品系苗期性状表现存在很大差异,ND13-4苗期的平均叶片数与对照相近,没有明显差异,而在平均茎粗方面是下降的,其它两个品系MD07-15和ZL02-3则增加,增加幅度在8.33%~11.11%。由此可知,3个品系对航天诱变环境的敏感性和细胞损伤程度不同,ND13-4对太空诱变环境不敏感,诱变后苗期性状变化幅度不大,而其它2个品系表现敏感,致畸率也高。

表4 诱变品系幼苗农艺性状差异
2.3 太空诱变对紫色马铃薯幼苗叶片保护酶的影响
由图1可以看出,各品系经太空诱变处理后,诱变品系的幼苗叶片中各保护酶SOD、CAT、POD活性均高于对照,但增强幅度不同。其中SOD活性较对照增强幅度最小,为11.88%~22.93%, POD活性增强幅度为23.53%~54.35%,CAT活性增强幅度最大,为19.67%~57.35%。各品系幼苗叶片保护酶活性对诱变环境的反应存在差异,品系MD07-15幼苗叶片SOD、CAT、POD活性平均增幅最大,而品系ZL02-3幼苗叶片中各酶活性增幅最小。
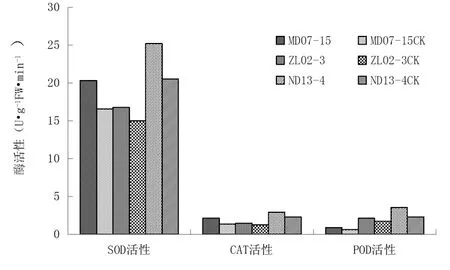
图1 诱变品系幼苗叶片SOD、CAT、POD的活性
2.4 变异株与对照幼苗性状差异分析
由表5可以看出,经太空诱变处理后,各品系的变异株与对照的差异主要表现在株高、茎和叶片的颜色变化方面,株高与对照差异较大,茎和叶片的颜色呈加深趋势,叶片边缘波状变强。究其原因,可能是高空的强幅射激发了种子形成色素的物质积累。

表5 变异株与对照幼苗性状差异分析
3 结论与讨论
与对照相比,经返回式科学实验卫星“实践十号”搭载的紫色马铃薯种子经过地面发芽试验和幼苗性状鉴定试验表明,总体上诱变后种子的发芽势和成苗能力大幅度下降,幼苗矮化,生长迟缓,茎叶颜色变深,幼苗叶片SOD、CAT、POD的活性增强,但也会出现个别较对照生长快的高变异株。
太空环境对植物种子的诱变效应差异很大[15],卫星搭载的番茄其SP1代发芽率比对照低[16-17],航天搭载黄芩种子发芽和成苗能力和SP1代植株生长受到抑制[3],这与本研究结果一致。另有研究表明小麦种子经航天处理却明显促进其发芽、出苗及生长[18-19],豌豆、红花、石刁柏等植物的种子经太空诱变处理后其活力也明显比地面对照种子高[20],这与本试验研究结果存在差异。作物遇到低温、盐、辐射等逆境后体内的多种酶活性会产生很大变化,酶活性增强和下降与植物的抗逆性密切相关[21],马铃薯幼苗遇低温胁迫时SOD、POD活性增加[22],小麦幼苗盐胁迫时POD活性增强[23],UV-B辐射使彩色马铃薯叶片中保护酶活性增强[24]。本试验结果表明,太空强辐射环境也使紫色马铃薯幼苗叶片SOD、CAT、POD的活性增强,且增幅较大,这与前人研究结果相似。
彩色马铃薯种子后代遗传复杂[25-26],本试验着重研究了太空诱变紫色马铃薯实生种子的发芽和成苗能力及幼苗群体的农艺性状、叶片酶活性变化情况,还需利用分子生物学手段进一步明确太空诱变材料变异体遗传物质的的真实性,并利用多代无性系繁殖技术检测诱变获得变异的可持续遗传稳定性等。
[1]刘珍珍.赤霉素对马铃薯实生种子萌发的影响[J].青海农林科技,2015(01):1-3.
[2]马娜,陈彦杞.紫色马铃薯的保健功效及栽培技术[J].河南农业,2016(21):51-52.
[3]单成钢,王志芬,苏学合,等.航天诱变黄芩种子对其SP1代的影响[J].核农学报,2008(02):188-191.
[4]杨明飞,姚红军,吴苏霓,等.我国航天诱变技术在育种上的应用进展[J].北方水稻,2014,44(06):78-80.
[5]李欣.不同生态条件对马铃薯主要性状的影响及遗传分析[D].哈尔滨:东北农业大学,2013.
[6]金光辉.我国马铃薯育种方法的研究应用现状及其展望[J].中国马铃薯,2000,3(14):184-186.
[7]范宏伟.马铃薯新品种黑美人及高产栽培技术[J].中国农技推广,2011,27(06):17-18.
[8]杨先泉,王翠,赵勤,等.60Coγ射线辐照马铃薯适宜剂量与效应研究[J].核农学报,2010,24(05):917-921.
[9]杨宇,李东,张乾.60Coγ辐射对马铃薯耐盐愈伤组织诱导的影响[J].安徽农业科学,2010,38(12):6140-6141.
[10]张洪亮,许庆芬,张荣华,等.紫外线辐射对马铃薯脱毒试管苗生长的影响[J].现代化农业,2014(02):33-35.
[11]马铃薯栽培生理[M].北京:中国农业出版社,1995:45.
[12]张新永,郭华春,戴华峰.增强UV-B辐射对彩色马铃薯叶片中相关保护酶活性的影响[J]. 西北植物学报,2009,29(05):968-974.
[13]陈建勋,王晓峰.植物生理实验学实验指导[M].广州:华南理工大学出版社,2006:72-73.
[14]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2004:211-221.
[15]贺鹏.航天诱变烤烟品种的发芽特性及酶活性变化研究[D].长沙:湖南农业大学,2008.
[16]李金国,刘敏,王培生,等,空间条件对番茄诱变作用及遗传的影响[J].航天医学与医学工程,2000,13(02):114-115.
[17]周秀艳,金晓霞,秦智伟,等.航天诱变育种及其在蔬菜中的应用[J].中国农学通报,2008(06):291-295.
[18]王广金,闰文义,孙岩,等.空间诱变选育小麦新品系的研究[J],黑龙江农业科学,2004(04):l-4.
[19]王广金,闰文义,孙岩,等.春小麦航天育种效果的研究[J].核农学报,2004,18(04):257-260.
[20]薛淮,刘敏.植物空间诱变的生物效应及其育种研空进展[J].生物学通报,2002,37(11):7-9.
[21]艾希珍,于贤昌,王绍辉,等.低温胁迫下黄瓜嫁接苗与自根苗某些物质含量的变化[J].植物生理学通讯,1999,35( 01) : 26-29.
[22]杨慧菊,郭华春.马铃薯幼苗低温胁迫的生理响应及品种耐寒性综合评价[J]. 西南农业学报,2016,29(11):2560-2566.
[23]宋旸,裴冬丽. 盐胁迫对小麦幼苗POD活性和同工酶的影响[J]. 湖北农业科学,2011,50(09):1759-1761.
[24]张新永,郭华春,戴华峰. 增强UV-B辐射对彩色马铃薯叶片中相关保护酶活性的影响[J]. 西北植物学报,2009,29(05):968-974.
[25]李先平,包丽仙,李山云,等.彩色马铃薯块茎色素研究进展[J].作物杂志,2009(01):4-8.
[26]刘思泱. 6个彩色马铃薯品种的核型及ISSR分析[D].呼和浩特:内蒙古农业大学,2010.
猜你喜欢
当代水产(2022年6期)2022-06-29
西北林学院学报(2022年3期)2022-06-09
今日农业(2020年14期)2020-12-14
新疆农业科学(2020年9期)2020-10-14
现代农村科技(2020年9期)2020-09-08
陕西农业科学(2019年3期)2019-04-09
云南农业科技(2019年2期)2019-04-04
科技知识动漫(2017年7期)2017-08-09
cookie world(2010年2期)2010-03-04
现代农业科技(2009年19期)2009-03-20
