
布鲁菌104M∶Omp19过表达株的构建与免疫保护性评价
2018-03-03 08:30杨巧玲宰晓东殷瑛李汭桦钱梦樱赵陶然李耀辉付玲徐俊杰
生物技术通讯 2018年1期
杨巧玲,宰晓东,殷瑛,李汭桦,钱梦樱,赵陶然,李耀辉,付玲,徐俊杰
军事科学院 军事医学研究院 生物工程研究所,北京 100071
布鲁菌是一种兼性胞内寄生的革兰阴性球杆菌,由其引起的布鲁菌病是一种人畜共患病,给畜牧业带来巨大损失并严重威胁人类健康[1]。根据宿主和抗原的变异,可将布鲁菌分为9个种型,在我国流行的主要为牛种、羊种和猪种[2]。布鲁菌的感染能力很强,可以通过呼吸道、消化道、皮肤黏膜等途径侵入机体,而人体主要是通过接触患病动物或食用未消毒的奶制品患病[3]。
疫苗是控制布鲁病疫情的重要手段[4]。常见的兽用布鲁菌疫苗为减毒活疫苗,如牛种S19、牛种RB51、羊种Rev.1等[5-7]。目前国际上其他国家尚未出现获批的人用布鲁菌疫苗,我国正在使用的人用布鲁菌疫苗为牛种弱毒株104M,分离自前苏联地区感染的胎牛,经连续传代减毒获得[8]。该菌株具有一定的免疫原性和保护性,但因采用皮上划痕的接种方式,而且多次接种会使机体致敏,因此在布鲁菌病流行地区无法实现大面积接种[9]。近年来,国内外研究者鉴定到多个布鲁菌保护性抗原,并对其中部分抗原如Omp16、Omp19、Omp31、核糖体蛋白L7/L12、Cu-Zn超氧化物歧化酶、胞质蛋白P39等的免疫保护性进行了深入研究[10]。然而,目前报道的重组亚单位疫苗的保护效果与减毒疫苗相比还存在一定差距,并不能对机体起到完全的保护作用[10]。
Omp19抗原是存在于细菌表面的外膜脂蛋白,在布鲁菌中的同源性达90%以上,在布鲁菌外膜蛋白与多粘菌素之间起桥梁作用,并在脂多糖固定结构的修正、细菌胞内生存的维持方面发挥重要功能[11]。Omp19在体内不仅能够活化树突状细胞,而且在无佐剂的情况下可有效对抗多种布鲁菌的感染[12]。据报道,Omp19联合佐剂免疫后机体可以产生较高水平的特异性抗体[13-14]。然而,104M中Omp19的表达水平以及免疫104M后机体可产生的针对Omp19的抗体水平尚不清楚。此外,增加104M中Omp19的表达量是否有助于提升针对Omp19的特异性抗体水平也尚不明确。因此,我们在本研究中将Omp19基因插入表达载体,转化104M株构建104M∶Omp19过表达株,比较了104M与104M∶Omp19中Omp19蛋白的表达水平;通过免疫小鼠,评价机体产生的针对Omp19的特异性抗体水平与细胞因子水平;利用菌株攻毒,评价104M与104M∶Omp19免疫后机体产生的保护作用。期望能通过增加104M中Omp19抗原的表达量来增强104M株的免疫保护效果,为新型布鲁菌疫苗的研制提供实验依据。
1 材料与方法
1.1 材料
6~8周雌性BALB/c小鼠购自北京维通利华实验动物技术有限公司;布鲁菌104M株、A19株为本实验室保存;大肠杆菌TransT1、BL21(DE3)株,EasyPfu聚合酶购自北京全式金生物技术有限公司;表达载体pNSGroE2His购自BioVector公司;限制性内切酶BamHⅠ、XbaⅠ,T4DNA连接酶购自New England Biolabs公司;基因组提取试剂盒购自Promega公司;胶回收试剂盒和小量质粒提取试剂盒购自Omega公司;CBA细胞因子检测试剂盒购自BD公司;胰蛋白胨大豆肉汤(TSB)培养基购自Oxoid公司;HRP标记的羊抗鼠IgG、鼠源An⁃ti-His抗体购自Abcam公司;Omp19蛋白由本室重组表达纯化制备;Anti-Omp19小鼠多克隆抗体由Omp19蛋白免疫小鼠后的血清制备。
1.2 构建Omp19-pNSGroE2His过表达质粒
接种104M单菌落于TSB培养基中,培养至对数期(D600nm约1.0),收集菌体,用细菌基因组提取试剂盒提取104M基因组DNA。根据GenBank公布的布鲁菌Omp19基因序列,去除其中的跨膜区和信号肽,同时结合表达载体pNSGroE2His的序列特点,采用Primer Premier 5.0软件设计引物,在上、下游分别添加BamHⅠ、XbaⅠ酶切位点,构建引物Omp19-F、Omp19-R;在pNSGroE2His载体酶切位点上下游选取30 bp左右序列构建鉴定引物对A-F、A-R。本研究所用引物均在上海生工生物技术服务有限公司合成,序列见表1。
以布鲁菌104M株基因组为模板,用Omp19-F、Omp19-R引物进行PCR扩增Omp19基因(PCR反应条件:预热94℃ 5 min;解链94℃ 20 s,退火58℃ 20 s,延伸 72℃ 1 min,30个循环)。所得PCR产物用1%琼脂糖凝胶电泳检测回收,回收产物用BamHⅠ和XbaⅠ双酶切,并同时双酶切载体pNSGroE2His。酶切产物经1%琼脂糖凝胶电泳鉴定并回收,用T4DNA连接酶连接过夜。取5 μL连接产物转化大肠杆菌Trans-T1感受态细胞,涂板培养,随机挑取阳性克隆进行PCR验证,送公司测序,选取成功插入载体的菌液样品提取质粒,即获得Omp19-pNSGroE2His过表达质粒。
1.3 构建104M∶Omp19过表达株
接种104M单菌落于TSB培养基中,37℃培养至D600nm约为 0.6,冰浴 30 min,4℃、5000 r/min离心20 min,收集所有菌体;用10 mL预冷的去离子水重悬菌体,离心,如此洗涤3次,最后用100 μL 10%甘油重悬菌体,制备104M感受态细胞。在感受态细胞中加入适量Omp19-pNSGroE2His过表达质粒混匀,冰浴30 min,2 kV、200 Ω、25 μF进行电转,立即加入1 mL SOC-B培养基,37℃、220 r/min振荡培养24 h,取500 μL菌体涂于氯霉素抗性TSA,37℃培养。
1.4 鉴定104M∶Omp19过表达株
挑取氯霉素抗性平板上的单菌落,以Omp19扩增引物和鉴定引物分别进行菌落PCR鉴定,阳性克隆测序进一步确认。为了验证Omp19在菌株中的过表达,利用Anti-His抗体、Anti-Omp19抗体对测序正确的菌株进行Omp19蛋白的Western印迹鉴定。
1.5 104M∶Omp19过表达株的免疫评价方案
50只6~8周的雌性BALB/c小鼠随机分为5组,通过皮下注射方式分别免疫104M∶Omp19过表达株及104M株,设高、低剂量组,PBS组作为阴性对照组,具体免疫方式见表2。
免疫后第0、35 d分别采血,1000 r/min离心收集血清,用间接ELISA法检测血清中针对Omp19的特异性抗体IgG。以大于阴性血清D值2.1倍的最大稀释度作为该样品的抗体滴度。
免疫后第49 d,各组的4只小鼠取脾细胞,用30 μg/mL Omp19蛋白体外刺激培养免疫组和对照组小鼠的脾细胞,72 h后收集上清,用CBA细胞因子检测试剂盒检测IFN-γ、IL-2、TNF-α、IL-17A和IL-10的分泌水平。所有小鼠脾细胞样品分别重复3次。
免疫后第35 d,各组的6只小鼠通过腹腔注射布鲁菌A19菌株(1×106CFU/只)的方式进行攻毒评价,1周后取出脾脏,用PBS溶解研磨,稀释后涂布平板,计算菌载量。结果以菌载量取对数后的平均值的形式(mean lgCFU)来表示,同时将免疫组小鼠脾脏中布鲁菌菌载量相对于对照组小鼠脾脏中菌载量下降的数值定义为保护指数,也就是对照组与实验组的平均lgCFU的差值。
1.6 统计学方法
实验结果以x±s表示,应用 Student′st检验对2组样本间均值差异进行统计学分析,以P<0.05表示差异具有统计学意义。
2 结果
2.1 过表达质粒Omp19-pNSGroE2His的构建
以布鲁菌104M基因组为模板扩增Omp19基因序列,1%琼脂糖凝胶电泳鉴定获得约500 bp的条带,与理论值相符(图1)。切取胶块用胶回收试剂盒回收,用BamHⅠ和XbaⅠ对片段和载体pNSGroE2His进行双酶切,回收后用T4DNA连接酶连接,转化大肠杆菌Trans-T1,涂板培养后进行PCR菌落鉴定,将菌落鉴定为阳性的菌液提交公司测序,结果显示插入片段的序列与Omp19基因的编码序列完全一致。
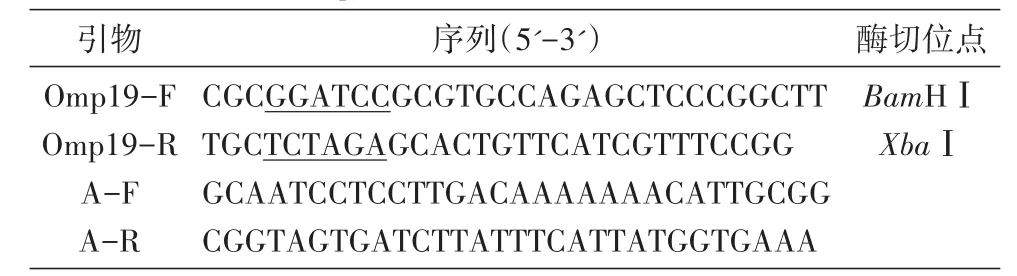
表1 Omp19过表达菌株构建及鉴定的引物
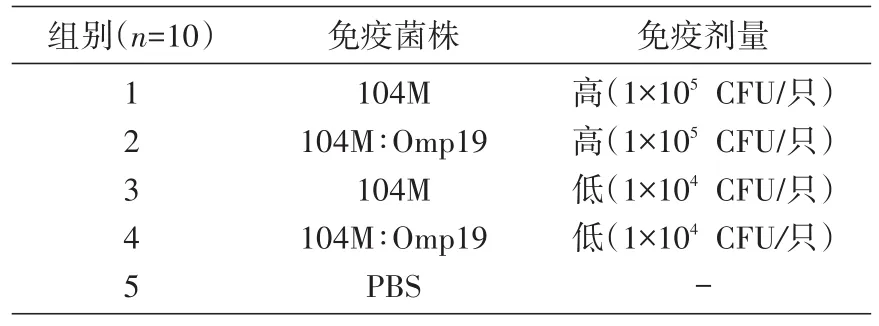
表2 Omp19过表达株的免疫评价
2.2 104M∶Omp19过表达株的构建与鉴定
将构建的过表达质粒Omp19-pNSGroE2His转入布鲁菌104M,培养后涂布在氯霉素抗性平板上,挑选转化子进行基因水平鉴定,分别用扩增引物和鉴定引物进行菌落PCR鉴定,均可鉴定出阳性克隆(图2)。
对基因水平鉴定正确的阳性克隆进一步进行蛋白水平的鉴定。培养48 h后,用超声波细胞破碎仪对菌体沉淀进行破碎,离心取上清进行SDS-PAGE分析。利用Anti-Omp19抗体对蛋白的表达进行Western印迹鉴定,结果如图3,104M中Omp19蛋白本身表达水平较低,而104M∶Omp19中Omp19蛋白的表达量明显增加。随后利用An⁃ti-His抗体对Omp19-pNSGroE2His质粒在104M株中的表达情况进行Western印迹鉴定,结果显示在104M的泳道中检测不到特异性条带的存在,而104M∶Omp19对应的泳道在相对分子质量约19 000处有特异性条带,大小与Omp19蛋白一致,提示转入的Omp19-pNSGroE2His质粒在104M中进行了表达。用Anti-Omp19抗体检测到的Omp19表达量明显增加是由于外源导入Omp19-pNSGroE2His质粒所致,这也进一步提示成功构建获得了104M∶Omp19过表达株。
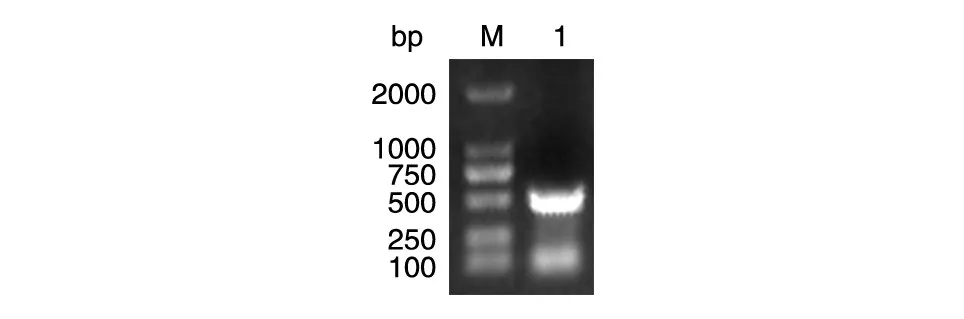
图1 布鲁菌Omp19基因的PCR扩增

图2 Omp19过表达株的基因鉴定
2.3 104M∶Omp19过表达株激发的抗体水平
小鼠免疫后第35 d取血,ELISA法检测针对Omp19的特异性抗体水平,结果如图4。104M高、低剂量免疫组小鼠血清中针对Omp19的特异性抗体滴度分别为267、637,表明104M疫苗免疫后小鼠产生的针对Omp19的抗体水平均较低;104M∶Omp19高、低剂量免疫组小鼠血清中针对Omp19的特异性抗体滴度分别为576、1905,显著高于104M组(P<0.05),表明通过提高Omp19的表达量,104M∶Omp19免疫组小鼠血清中针对Omp19的特异性抗体水平明显增强。
2.4 104M∶Omp19过表达株激发的细胞因子水平
小鼠初次免疫后第49 d,用CBA细胞因子检测试剂盒检测Omp19抗原体外刺激后小鼠脾细胞分泌的细胞因子水平,结果如图5。Omp19抗原刺激后,104M与104M∶Omp19高、低剂量免疫组小鼠的淋巴细胞培养上清中均能检测到细胞因子 TNF-α、IFN-γ、IL-2、IL-10和IL-17A 的表达,104M∶Omp19高剂量免疫组的TNF-α、IFN-γ、IL-2表达水平均显著高于104M组(P<0.05)。Th1细胞主要分泌的细胞因子包括TNF-α、IFN-γ和IL-2等,可以介导细胞毒性免疫反应以及和局部炎症有关的免疫反应。Th2细胞主要分泌的细胞因子包括IL-4和IL-10等,对清除胞外病原微生物有重要作用。结果表明,与104M组相比,在体外Omp19蛋白的刺激下,104M∶Omp19免疫组小鼠的淋巴细胞能产生更高水平的以分泌TNF-α、IFN-γ、IL-2为特征的Th1型细胞介导的免疫反应,以及以分泌IL-10为特征的Th2型细胞介导的免疫反应。

图3 Omp19过表达株的Western印迹鉴定
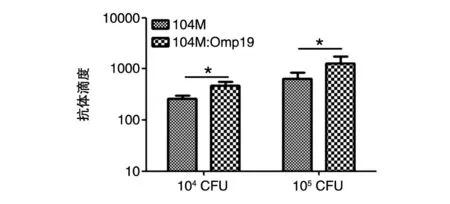
图4 免疫小鼠血清中抗原特异性抗体水平检测
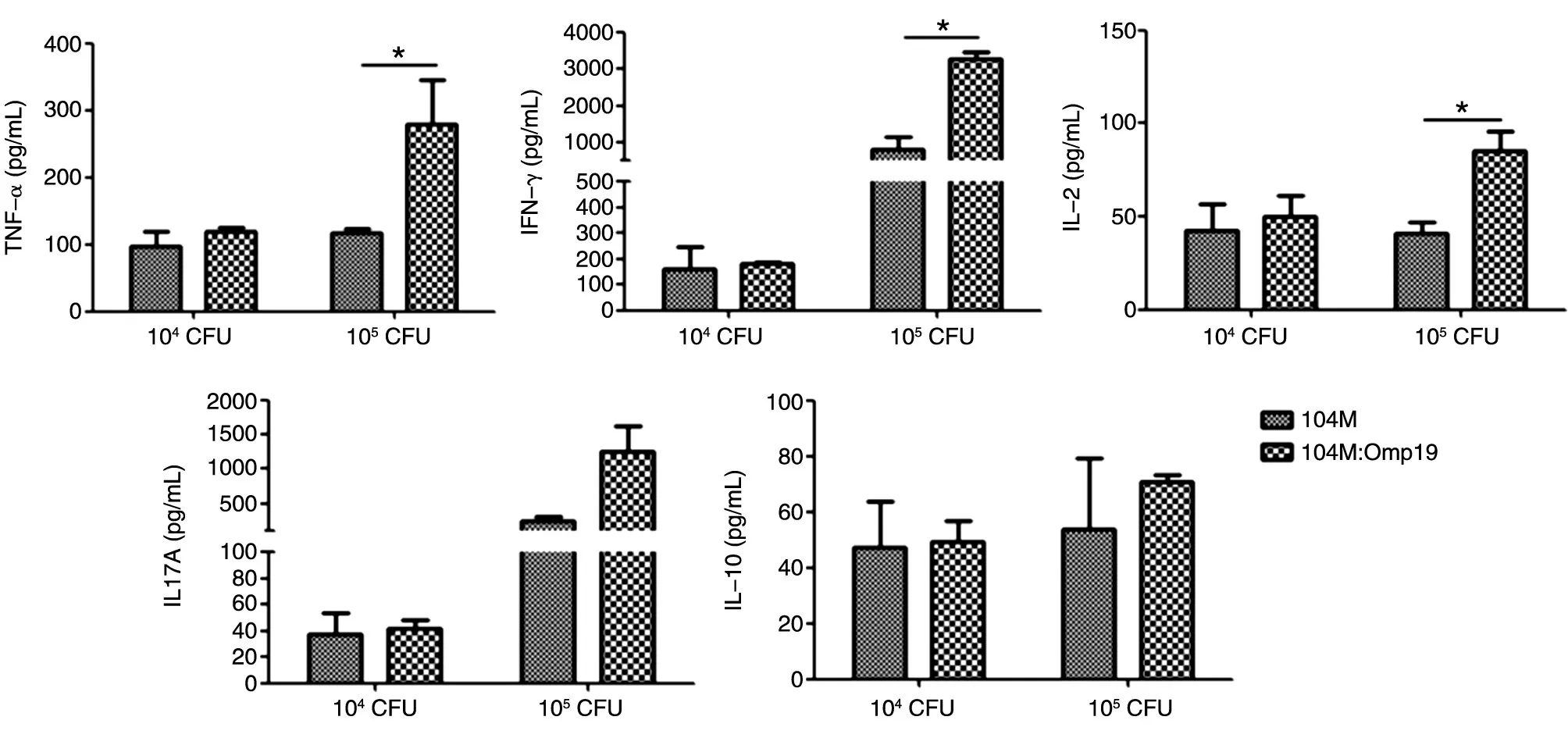
图5 免疫小鼠脾细胞抗原刺激后细胞因子分泌水平
2.5 104M∶Omp19过表达株的免疫保护效果
小鼠初次免疫后第35 d,用布鲁菌A19(1×106CFU)对免疫的小鼠进行腹腔攻击,利用保护指数评价104M∶Omp19过表达株的保护效力,结果见表3。与PBS对照组相比,攻毒后104M和104M∶Omp19高、低剂量免疫组小鼠脾脏中的菌载量均显著降低(P<0.01),显示出保护效果。104M高剂量免疫组脾脏菌载量降低了2.02个数量级,低剂量组降低了1.56个数量级;104M∶Omp19高剂量免疫组脾脏菌载量降低了2.56个数量级,低剂量组降低了2.07个数量级。在同等免疫剂量条件下,104M∶Omp19高、低剂量组的保护指数均大于相应的104M组,且在低剂量免疫组中这种保护作用的差异具有统计学意义(P<0.05)。在达到同等保护指数(约2.0)时,104M∶Omp19的免疫剂量(1×104CFU/只)只需104M(1×105CFU/只)的1/10。这提示,104M∶Omp19免疫小鼠可以显著降低小鼠攻毒后脾脏中的菌载量,与104M相比,保护效果显著增强。

表3 小鼠攻毒实验保护性效果统计表
3 讨论
疫苗在防控布鲁菌病疫情方面发挥重要作用,虽然减毒活疫苗在控制动物疫情方面作用显著,但是其对人体致病,因而限制了使用。目前国际上其他国家尚未出现人用布鲁菌疫苗,我国正在使用的人用布鲁菌疫苗为牛种弱毒株104M。该菌株虽然具有一定的免疫原性和保护性,但仍存在较大的副作用。而且由于皮上划痕的接种方式难以控制接种量,导致人们的接受性普遍较低。Omp19抗原是一种布鲁菌外膜脂蛋白,在细菌中发挥重要功能。研究表明Omp19作为保护性抗原可有效刺激小鼠产生特异性抗体和细胞因子,同时保护小鼠抵抗布鲁菌的侵袭[14]。
本研究通过将Omp19基因与过表达质粒连接后转入104M株的方式构建了104M∶Omp19过表达株,评价了104M与104M∶Omp19中Omp19的表达水平,发现104M中Omp19的表达水平较低,而104M∶Omp19中Omp19的表达量明显增加;通过免疫小鼠,评价了104M与104M∶Omp19免疫后机体产生的Omp19的特异性抗体水平与细胞因子水平,发现104M疫苗免疫后小鼠产生的针对Omp19的抗体水平与细胞因子水平均较低;通过构建104M∶Omp19过表达株,提高Omp19的表达量,发现可显著增强Omp19的特异性抗体水平以及细胞因子水平;通过A19弱毒菌株攻毒,评价了104M与104M∶Omp19免疫后所诱导的免疫保护效果,发现104M∶Omp19免疫小鼠后,可以显著降低小鼠脾脏菌载量,与104M相比,保护效果显著增强。在达到同等保护指数时,104M∶Omp19的免疫剂量只需104M的1/10,有望实现减少疫苗接种剂量,降低104M自身的副作用。
布鲁菌作为胞内寄生菌,能够逃避宿主免疫反应,从而建立慢性感染。体液免疫和细胞免疫反应对于病原体的清除均发挥关键作用。小鼠淋巴细胞体外刺激实验结果表明,与104M相比,免疫104M∶Omp19组小鼠的淋巴细胞在接受抗原刺激后可以显著增强抗原特异的Th1偏向型细胞因子TNF-α、IFN-γ、IL-2和Th2偏向性细胞因子IL-10的产生,这也提示104M株中Omp19的过表达可以有效增强细胞介导的免疫反应。攻毒实验表明,104M∶Omp19免疫组的小鼠脾脏中的菌载量显著低于104M免疫组,推测这可能是由于过表达株中Omp19抗原的表达量提高,增强了小鼠体内布鲁菌特异的体液免疫和细胞免疫反应,从而提高了宿主对布鲁菌的清除能力。综上所述,我们成功构建了布鲁菌疫苗104M的过表达株104M∶Omp19,在小鼠模型中评价了相应的免疫原性和保护作用。初步研究结果表明,相对于104M疫苗,Omp19的过表达可以显著增强原有疫苗的免疫保护效力,104M∶Omp19可作为一种新的布鲁菌疫苗候选菌株用于后续研究。
[1] Franco M P,Mulder M,Gilman R H,et al.Human brucellosis[J].Lancet Infect Dis,2007,7(12):775-786.
[2] Atluri V L,Xavier M N,de Jong M F,et al.Interac⁃tions of the human pathogenic Brucella species with their hosts[J].Ann Rev Microbiol,2011,65(1):523-541.
[3] Li Y J,Li X L,Liang S,et al.Epidemiological fea⁃tures and risk factors associated with the spatial and temporal distribution of human brucellosis in China[J].BMC Infect Dis,2013,13(1):547.
[4] Nielsen K,Duncan J R.Animal brucellosis[M].CRC Press,1990.
[5] Hanson G W.Brucellosis in humans and animals[J].Ceylon Med J,2006,52(2):66.
[6] Schurig G G,Sriranganathan N,Corbel M J.Brucello⁃sis vaccines:past,present and future[J].Vet Microbiol,2002,90(1-4):479-496.
[7] Nicoletti P.Vaccination of cattle with Brucella abortus strain 19 administered by differing routes and doses[J].Vaccine,1984,2(2):133-135.
[8] Yu D,Hui Y,Zai X,et al.Comparative genomic anal⁃ysis of Brucella abortus vaccine strain 104M reveals a set of candidate genes associated with its virulence attenuation[J].Virulence,2015,6(8):745.
[9]孙养信,王丽.人用布鲁菌104M冻干活菌苗免疫效果[J].中华预防医学杂志,2001,35(1):54.
[10]Carvalho T F,Haddad J P,Paixão T A,et al.Metaanalysisand advancementofBrucellosisvaccinology[J].PLoS One,2016,11(11):e0166582.
[11]Tibor A,Decelle B,Letesson J J.Outer membrane proteins Omp10,Omp16,and Omp19 of Brucella spp.are lipoproteins[J]. Infect Immun, 1999,67(9):4960-4962.
[12]Pasquevich K A,Ibañez A E,Coria L M,et al.An oral vaccine based on U-Omp19 induces protection againstB.abortusmucosalchallengebyinducingan adaptiveIL-17 immune responsein mice[J].PLoS One,2011,6(1):e16203.
[13]Tadepalli G,Singh A K,Balakrishna K,et al.Immu⁃nogenicity and protective efficacy of Brucella abortus recombinant protein cocktail(rOmp19+rP39)against B.abortus 544 and B.melitensis 16M infection in murine model[J].Mol Immunol,2016,71:34.
[14]AbkarM,LotfiA S,AmaniJ,etal.Survey of Omp19 immunogenicityagainstBrucellaabortusand Brucella melitensis:influence of nanoparticulation ver⁃sus traditional immunization[J]. Vet Res Commun,2015,39(4):217-228.
猜你喜欢
灾害医学与救援(电子版)(2018年1期)2018-06-05
生命科学研究(2018年1期)2018-05-29
中国医药指南(2017年3期)2017-11-13
天津医科大学学报(2015年2期)2015-12-22
华南农业大学学报(2015年5期)2015-12-04
中国全科医学(2015年33期)2015-12-02
肿瘤预防与治疗(2015年1期)2015-09-26
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年11期)2015-06-10
中国当代医药(2015年9期)2015-03-01
