
克氏原螯虾雌性生殖系统发育及组织结构观察
2018-03-02 08:11宋光同何吉祥吴本丽
江西农业学报 2018年2期
宋光同,何吉祥,吴本丽,陈 静,黄 龙,汪 翔,武 松
克氏原螯虾(Procambarus clarkii)隶属于节肢动物门(Arthropada)、甲壳纲(Crustacea)、十足目(Decapoda)、爬行亚目(Reptantia)、螯虾科(cambaridae)、原螯虾属(Procambarus),俗称淡水小龙虾。克氏原螯虾原产于北美洲,在1929年由日本引入我国江苏南京地区。该虾具有适应性广、繁殖力强、生长快、肉质鲜美、营养丰富、抗病力强、适于稻田养殖等特点,广受国内外养殖者和消费者青睐,为我国主要经济虾类之一。
2016年,全国小龙虾养殖总规模超60万hm2,总产量达85.23万t,且发展势头不减,养殖户扩展养殖热情依然高涨。现在对克氏原螯虾优质虾苗需求量与日俱增,而我国至今尚无规模化苗种繁育场,苗种主要来自野外捕捞、商贩收集和自繁自养等,优质苗种生产严重滞后于养殖需求,弱苗、次苗、伤苗充斥市场,直接导致50%以上的养殖户养殖失败,严重影响了龙虾产业可持续发展。
国内外关于克氏原螯虾繁殖生物学的研究报道较多,该虾性腺发育、交配、产卵皆不同步[1-8]。 多数学者研究认为:克氏原螯虾的交配期为5~9月份,7、8月为交配高峰期,交配后,精子储存在纳精囊中,等待排卵受精;一年产卵1次,存在4、5月和9、10月两个产卵高峰期,以 9、10 月份为主[6-8]。 也有学者认为极少数克氏原螯虾个体在产卵后经3~4个月恢复后再次成熟产卵[9]。关于克氏原螯虾生殖系统发育的分期,主要根据其卵巢外部特征的周年变化,结合卵子发生过程特征,将克氏原螯虾卵巢发育分为5~6期[9-13]。但是对克氏原螯虾性腺发育过程中卵巢、输卵管的显微及超微结构缺乏详细研究和介绍。我们采用雌性生殖系统外部特征、组织显微、超微结构相结合的方法,对克氏原螯虾的卵巢颜色和大小、卵子发生、卵巢壁、输卵管以及内质网、线粒体等结构的变化特征进行了系统的观察研究,以期更好地掌握克氏原螯虾雌性生殖系统发育的内在客观规律,为该虾人工繁育提供理论基础资料。
1 材料与方法
1.1 试验材料
实验用克氏原螯虾均从东至县大联圩农业开发有限公司养殖池塘采集,自2015年8月至2016年7月采用投放地笼的方式随机捕获样本虾,每月采集1~2次,其中繁殖盛期(8~10月份)每月采集2次,其他月份每月采集1次。选取活力强、附肢完整、体质量17.52~44.68 g的雌性克氏原螯虾样本,每次采集18~25尾,选择3尾用于石蜡组织切片实验;并对剩下的样本虾(15~22尾)进行称重和解剖观察,观察不同季节克氏原螯虾卵巢、输卵管的形态、大小和颜色,并对解剖出的卵巢称重,计算卵巢的性腺指数,性腺指数(GI)=100%×卵巢湿重/活体体质量。分别于2015年9月26日、2016年1月16日和2016年6月29日,采集雌性样本虾各1尾,用于卵巢及左右输卵管超微结构的观察实验。
1.2 实验方法
1.2.1 石蜡切片 将雌虾头胸甲从背面拨开,迅速取出卵巢及左、右输卵管组织,使用波恩试液固定20 min;再将卵巢、左输卵管、右输卵管分开,分别固定于波恩试液中24 h以上;利用脱水机(JJ-12J),按照75%(4 h)、85%(2 h)、90%(2 h)、95%(1 h)、100%(30 min)、100%(30 min)的酒精梯度做梯度脱水;用体积比为 1∶1的 100%酒精+二甲苯(5~10 min)和二甲苯Ⅰ(5~10 min)、二甲苯Ⅱ(5~10 min)给样品做透明;用石蜡Ⅰ(1 h)、石蜡Ⅱ(1 h)、石蜡Ⅲ(1 h)做浸蜡,用包埋机(JBP5)做石蜡包埋,用切片机(RM2016)切片,切片厚度为4μm;最后用 H.E染色,用中性树脂封片;用科研级显微镜(Olympus bx53)观察拍照。
1.2.2 透视电镜超薄切片 将采集的活体克氏原螯虾雌虾带回实验室,在4℃左右的低温环境中迅速解剖其生殖系统组织,置入预冷的4%戊二醛固定液中;在4℃下预固定20 min后,将卵巢、左输卵管、右输卵管分开,移入盛有预冷的4%戊二醛固定液的2 mL离心管中,在4℃下固定过夜。将固定的样品用缓冲液冲洗数次,再用1%锇酸再次固定,脱水,用环氧树脂包埋,用超薄切片机切片,用醋酸铀柠檬酸铅进行双重染色,用日立HT-7700型透射电镜观察拍照。
2 结果与分析
2.1 克氏原螯虾卵巢成熟系数的周年变化
从图1~图2可以看出:在4~6月,克氏原螯虾的性腺指数为0.10%~0.13%,为较低水平期;在7~9月,其性腺指数为0.53%~4.71%,为增长期;在10、11月份,随着部分虾产卵,性腺指数急速下降,仅为0.81%~0.83%;之后,性腺指数又逐步升高,至次年2月份,形成另一个峰值,达3.97%;在3~4月份,性腺指数又急速下降。克氏原螯虾性腺指数分别在9月份和2月份出现峰值。
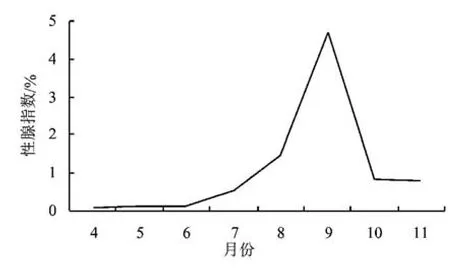
图1 克氏原螯虾性腺指数在4~11月的变化

图2 克氏原螯虾性腺指数在10月~翌年5月的变化
2.2 解剖结构及生殖腺组织结构
2.2.1 解剖结构 克氏原螯虾雌性生殖系统由卵巢和输卵管组成,卵巢呈“Y”型结构(图3),位于头胸甲背侧中央,其前端为胃,上端为心脏,下端为肝胰脏。成熟的卵巢后端伸向头胸甲与腹节交界处,前端伸至额剑后缘。在卵巢发育早期,卵巢呈白色透明状,体积较小,卵粒小而不易分离;随着卵巢的发育,卵巢体积逐渐变大,卵巢不断向前、后、左右延伸膨胀,卵粒大而易分离;卵巢颜色变化依次为白色透明、白色、黄色、咖啡色、深棕色、黑色。输卵管分为左右输卵管,从卵巢两侧发出,沿围心腔外壁向腹侧延伸,最后开口于第3对胸足基部,形成雌性产卵孔。
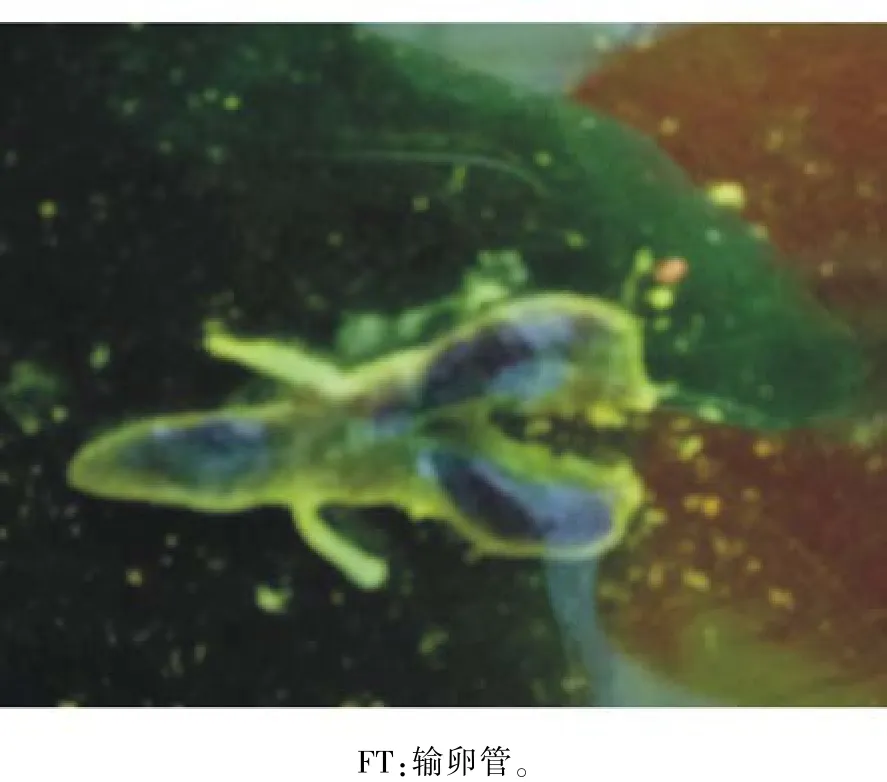
图3 克氏原螯虾雌性生殖系统的解剖结构
2.2.2 卵巢壁 卵巢壁为卵巢外膜,为疏松结缔组织。在卵巢发育早期,疏松结缔组织较厚,内含血管、血窦,分化滤泡细胞;随着卵巢发育,疏松结缔组织逐渐变薄,到卵巢成熟期,仅为薄薄一层;卵巢排卵后,疏松结缔组织又开始增厚(图4-1~图4-7)。
2.2.3 卵室 在卵巢发育早期,在卵巢中央布满卵室,卵室由外膜和内生殖上皮组成,外膜主要由肌层组织组成,内生殖上皮分化增殖卵原细胞。随着卵原细胞生长发育成卵母细胞,卵室消失,卵母细胞被滤泡细胞包围;排卵后,卵室再次逐渐增多(图4-1、图4-7)。
2.2.4 输卵管 左右输卵管的外观解剖结构基本一致,在生殖腺发育初期,输卵管管径小,呈灰白色;随着生殖腺发育,管径逐渐增大,进入成熟期,管径达到最大,呈乳白色。切片显示,输卵管由外膜、基膜和管壁上皮组成。外膜为一层疏松结缔组织,内含血管;基膜紧贴管壁上皮外面;管壁上皮为单层柱状上皮,在发育初期,外膜较厚,随着性腺发育,逐渐变薄。输卵管内含物从无到有,逐渐增多,在发育中期内含物嗜碱性,在发育后期转为嗜酸性物质,同时输卵管中出现丰富的白色液泡状物质(图5-8~图5-14)。
2.3 卵子发生
根据克氏原螯虾卵母细胞的大小、形状、染色、核仁数目、卵黄积累情况,以及卵母细胞与滤泡细胞的关系,将克氏原螯虾卵子的发生划分为卵原细胞、卵黄合成前卵母细胞、卵黄合成期卵母细胞及成熟期卵母细胞4个时相。
2.3.1 卵原细胞(增殖期) 卵原细胞附着于卵室内边缘,多数近圆形或卵圆形,长径2.89~6.47μm,短径1.54~4.06μm;细胞核大,核质着色浅,染色质呈絮状聚集,长径1.99~4.80μm,短径 0.88~3.06 μm;核仁较多,多数沿核膜边缘分布;少量卵原细胞较大,染色质呈细线状。细胞质少,围绕在细胞核周围形成很薄的一层,嗜碱性。卵巢壁疏松结缔组织内分化增殖滤泡细胞(图4-1)。
2.3.2 卵黄合成前卵母细胞(小生长期) 卵母细胞迅速增大,细胞质增多,卵母细胞近圆形或不规则形,长径 59.85~134.32μm,短径 35.05~93.74μm。细胞核圆形或卵圆形,长径20.75~37.22μm,短径17.42~25.66μm,核质内充满絮状颗粒;核仁8~10个,呈颗粒状,染色较深,沿核膜边缘分布。细胞质呈嗜碱性,布满蓝色颗粒,无卵黄积累。滤泡细胞开始分割包围卵母细胞,滤泡细胞多呈扁平、卵圆形(图 4-2)。
2.3.3 卵黄合成期卵母细胞(大生长期) 根据卵黄积累、大小、分布情况,将此发育期分为初期、中期、后期。
初期(大生长期1):卵母细胞继续增大,卵母细胞呈不规则圆形,长径 115.73~261.88μm,短径90.02~178.44μm;核卵圆形,长径22.05~35.27μm,短径20.54~28.63μm,核质内充满絮状颗粒,核仁8~10个,沿核膜边缘分布。细胞膜边缘开始聚集细小颗粒状卵黄,呈嗜酸性,被染成浅红色,中央出现少量大颗粒卵黄颗粒;细胞质充满白色空泡;卵母细胞四周的滤泡细胞增多,滤泡细胞呈扁平形、卵圆形、圆形(图 4-3)。
中期(大生长期2):卵母细胞呈不规则圆形,长径 187.20~196.22μm,短径 149.18~183.27 μm;核卵圆形,被染成蓝色,长径19.48~42.74μm,短径16.75~35.38μm,核仁6~8个,沿核膜边缘分布。细胞质嗜酸性,被染成红色,胞质中卵黄颗粒由细胞膜边缘向中央扩散,粒径逐渐增大,最大达2.80μm;细胞质充满白色空泡,在细胞核膜周边可见蓝色核物质;滤泡细胞呈近圆形(图4-4)。
后期(大生长期3):卵母细胞呈卵圆形,长径287.54μm,短径207.9μm,细胞核开始皱缩,细胞质嗜酸性,在细胞质中充满大颗粒状卵黄颗粒,粒径最大达3.5μm;细胞核外围周边染色较浅,细胞质充满白色空泡。滤泡细胞呈圆形(图4-5)。

图4 克氏原螯虾卵巢、卵巢壁和卵室的组织结构
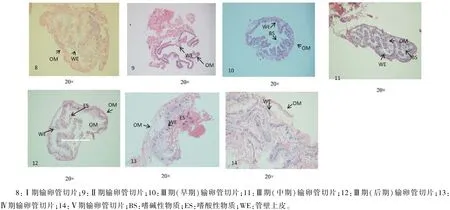
图5 克氏原螯虾输卵管的组织结构
2.3.4 成熟期卵母细胞 卵细胞近圆形,直径252.6 μm。细胞核消失,细胞质嗜酸性,细胞质中充满大量大颗粒卵黄,粒径最大达6.70μm。卵母细胞与滤泡细胞已发生分离(图4-6)。
2.4 卵巢发育分期
根据克氏原螯虾卵巢的外形、大小与颜色,卵子发生时相及卵巢壁、输卵管的结构特征,可将克氏原螯虾卵巢发育期分为5个发育期。
2.4.1 Ⅰ期 以卵原细胞为主要时相。解剖结构观察,卵巢呈白色透明状,卵粒细小,隐约可见,不可分离,附着在卵巢膜上。切片观察,卵巢内布满卵室,边缘上分化增殖众多卵原细胞,为卵原细胞增殖中心;卵巢壁厚度10.62~31.2μm;输卵管的外膜疏松结缔组织较厚,内含血管;管壁上皮为单层柱状上皮,上皮细胞呈卵圆形,较粗壮,向输卵管内陷弯曲(图4-1、图4-8)。 每年12月至翌年3月份,以Ⅰ期卵巢为主,部分小规格红壳虾卵巢处于Ⅳ期、Ⅴ期。
2.4.2 Ⅱ期 以卵黄合成前卵母细胞为主要时相。解剖结构观察,卵巢呈白色,卵巢明显变大,卵粒细小可见,不可分离,平均卵径0.18~0.48 mm。切片观察,卵巢内布满卵黄合成前卵母细胞,滤泡细胞开始分割包围卵母细胞;卵巢中有少量卵原细胞;存在退化的卵母细胞。卵巢壁厚8.20~25.52μm;输卵管管壁单层柱状上皮变细,弯曲度变小(图4-2、图4-9)。每年4、5月以Ⅱ期卵巢为主,部分小规格个体卵巢处于Ⅰ、Ⅴ期。
2.4.3 Ⅲ期 为卵黄合成期(大生长期)。根据卵巢的颜色、大小及卵黄颗粒的积累、大小、分布等情况,将该发育期分为3个时期。
Ⅲ期早期:解剖结构观察,卵巢呈黄色,卵巢卵粒可分辨,但仍不可分离;卵径0.5~1.2 mm。切片观察,卵巢内以大生长期1卵母细胞为主,存在1~2个小生长期卵母细胞;卵巢壁厚4.36~19.36μm;输卵管管壁单层柱状上皮进一步变细,弯曲度进一步变小,输卵管内开始分泌少量嗜碱性物质(图4-3、图4-10)。每年6、7月以Ⅲ期早期卵巢为主,部分个体卵巢处于Ⅱ期。
Ⅲ期中期:解剖结构观察,卵巢呈咖啡色,卵巢卵粒可分辨,可分离,卵径1.2~1.5 mm。卵巢内以大生长期2卵母细胞为主,卵母细胞周围滤泡细胞增多,滤泡细胞核由扁平状变为卵圆形,存在1~2个大生长期1卵母细胞;卵巢壁厚4.17~13.77μm;输卵管外膜变薄,管壁单层柱状上皮粗壮,弯曲度增大,输卵管内分泌嗜碱性物质(图4-4、图4-11)。每年8月以Ⅲ期中期卵巢为主,部分规格较小的个体卵巢处于Ⅱ期、Ⅲ期早期。
Ⅲ期晚期:解剖结构观察,卵巢呈深褐色,卵粒可分辨,可分离,卵径1.5~1.8 mm。卵巢内以大生长期3卵母细胞为主。卵巢壁厚4.92~27.83μm;输卵管的外膜疏松结缔组织进一步变薄,管壁单层柱状上皮粗壮,输卵管内开始分泌少量嗜酸性物质(图4-5、图4-12)。该发育期主要出现在9月上中旬,但在9月上中旬部分规格较小的个体卵巢处于Ⅲ期早、中期,也有部分较大个体卵巢发育达Ⅳ期。
2.4.4 Ⅳ期 以成熟期卵母细胞为时相。卵巢几乎充满整个头胸甲,呈黑色,卵粒饱满均匀,挑破卵膜后卵粒即可分离,卵径1.8~2.0 mm。切片观察,以成熟期卵母细胞为主,滤泡细胞和卵母细胞已分离。卵巢壁极薄,为2.57μm;输卵管内分泌大量嗜酸性物质,出现大量液泡状物质,准备排卵(图4-7、图4-13);9月下旬~10月上中旬以Ⅳ期卵巢为主,也有部分个体处于Ⅲ期、Ⅴ期卵巢。
2.4.5 Ⅴ期 为产后恢复期。此期卵巢呈血红色,卵巢体积缩小,卵巢内只有残存的少量黄色退化卵粒。切片观察,卵巢仅剩排空的滤泡腔空泡和正在退化的卵母细胞,卵巢壁厚度5.18~10.45μm;输卵管内分泌的酸性物质开始分解扩散(图4-8、图4-14)。在11月份以Ⅴ期卵巢为主,部分规格较小个体处于Ⅳ期卵巢。
2.5 克氏原螯虾雌性生殖系统的超微结构
2016年1月26日:样本卵巢呈浑白色,超微结构观察,卵质中含有丰富的块状肌层组织(图6-15),还含有少量管状滑面内质网、环状粗面内质网,内含线粒体(图6-16、图6-17)。左输卵管含有丰富的管状、环形滑面内质网,少量线粒体和环状粗面内质网,发现部分退化的线粒体(图 6-18、图 6-19)。右输卵管含有环状、管状粗面内质网,也发现退化的线粒体(图 6-20)。
2016年6月26日:样本卵巢呈咖啡色,超微结构观察,卵质中积累了大量呈囊泡状滑面内质网,分布较多致密小颗粒物质,并逐渐向囊泡状滑面内质网内积聚,并越积越多,使整个滑面内质网演变成均匀致密、近球形的具膜大卵黄粒,卵质充满卵黄颗粒(图6-21);卵质中多数线粒体脊断裂、脱落,部分线粒体与囊泡状滑面内质网相互融合(图6-22)。滤泡细胞质中含有丰富的线粒体和少量囊泡状粗面内质网,在靠近卵母细胞一边,可见排列整齐的长泡状结构(图6-23)。左输卵管含有丰富的纹理状结构、不规则泡状结构及粗面内质网(图6-24、图6-25);右输卵管中含有软管结构,软管间含有丰富的线粒体、短管状粗面内质网(图6-26)。
2015年9月26日:样本卵巢呈黑色,超微结构观察,卵质中含量丰富的线粒体、管状粗面内质网、空泡状脂滴以及大的卵黄颗粒,可见高尔基体(图6-27~图6-29);左输卵管含有丰富的肌层组织(图6-30);右输卵管含有丰富的管状粗面内质网(图6-31)。
3 讨论
3.1 关于卵巢、输卵管的结构特点
多数虾类卵巢分为左右两叶,如日本新糠虾卵巢呈“H” 型[14];罗氏沼虾卵巢左右两叶呈桑椹状[15];哈氏仿对虾卵巢分为左右两叶,在前端第一腹节处愈合[16];脊尾对虾卵巢呈菱形[17];粗糙对虾卵巢分成对称的左右两叶,相互贴近,部分愈合[18];黑斑口虾蛄卵巢分为两叶,左右对称,基本愈合[19]。克氏原螯虾卵巢分为3叶,呈“Y”型结构。克氏原螯虾卵巢壁疏松结缔组织内含血管、血窦,与营养物质和氧气运输有关,与中华绒螯蟹[20]、Penaeus setiferus[21]等相同;而粗糙对虾[19]、罗氏沼虾[15]的疏松结缔组织中不含血管、血窦。克氏原螯虾的疏松结缔组织分化滤泡细胞,卵室内生殖上皮分化增殖卵原细胞;而中华绒螯蟹[20]、Penaeus setiferus[21]、南美白对虾[22]等卵巢壁上的内生殖上皮同时分化生殖卵原细胞和滤泡细胞。
在克氏原螯虾卵巢成熟期,输卵管内分泌物逐渐由嗜碱性变为嗜酸性,并在输卵管内含有大量白色液泡状物质。其嗜酸性物质和白色液泡状物质是否与排卵有关,需要进一步研究。 邓道贵[18]、King[21]皆提到输卵管中分泌一种物质,有利于卵子排出。

图6 克氏原螯虾卵巢和输卵管的超微结构
3.2 关于卵子发生问题
目前,多数学者根据虾类卵细胞的大小、生长、迁移以及染色、卵黄积累等情况,进行卵子发生期划分,例如栗志国等[17]将脊尾白虾卵子的发生划分为卵原细胞、卵黄合成前期卵母细胞、内源性卵黄合成期卵母细胞、外源性卵黄合成期卵母细胞及成熟期卵母细胞5个时期;王春琳等[19]将黑斑口虾蛄的卵细胞的发育分为卵原细胞期、卵黄形成前期卵母细胞、卵黄形成期卵母细胞、成熟期卵细胞四个时期;上官步敏等[23]将锯缘青蟹的卵子发生分为卵原细胞、卵黄发生前期卵母细胞、卵黄发生期卵母细胞三个时期;洪水根等[24]将长毛对虾的卵子发生划分为卵原细胞、卵黄发生前卵母细胞和卵黄发生期卵母细胞三个时期;薛鲁征等[25]将中华绒螯蟹的卵子发生分为卵原细胞期、初级卵母细胞期、成熟前期和成熟期。本文结合上述的卵子发生划分方法,根据克氏原螯虾卵母细胞大小、形状、染色、核仁数目、卵黄的积累情况,以及卵母细胞与滤泡细胞的关系,将克氏原螯虾卵子的发生划分为卵原细胞、卵黄合成前卵母细胞、卵黄合成期卵母细胞及成熟期卵母细胞4个时相。其中,根据卵黄积累、大小、分布情况,将卵黄合成期卵母细胞分为初期、中期、后期。
3.3 关于卵巢发育分期问题
虾类卵巢分期,主要依据卵巢大小、颜色、成熟系数以及内部组织学特征。如郑忠明等[16]将哈氏仿对虾的卵巢发育分为形成期、小生长期、大生长期前期、大生长期后期、成熟期和恢复期6个时期;栗志国等[17]将脊尾白虾的卵巢发育分为增殖期、小生长期、大生长期、成熟期及产后恢复期;上官步敏等[23]将锯缘青蟹的卵巢发育分为未发育期、发育早期、发育期、将成熟期、成熟期和排卵后期6个时期;颜素芬等[22]将南美白对虾的卵巢发育分为增殖期、小生长期、大生长期早期、将成熟期、成熟期和恢复期6个时期;薛鲁征[25]将中华绒螯蟹的卵巢发育分为5期。本文根据克氏原螯虾卵巢的外形、大小与颜色,卵子发生时相以及卵巢壁、输卵管的结构特征,将克氏原螯虾的卵巢发育期分为5个时期:Ⅰ期、Ⅱ期(小生长期)、Ⅲ期(大生长期)、Ⅳ期(成熟期)和Ⅴ期(产后恢复期),其中根据卵黄的积累情况,将Ⅲ期(大生长期)又分为早期(大生长期1)、中期(大生长期2)、后期(大生长期3)。在7~9月份,随着克氏原螯虾交配,性腺发育加快,卵巢内卵黄合成旺盛,卵黄逐渐增多,交配行为是否是促进性腺加速发育的一个刺激因素,需要进一步研究。结合卵巢的颜色及卵母细胞的特征,本研究认为,卵巢呈黑色,卵黄粒径大,细胞核消失,卵母细胞与滤泡细胞分离可作为克氏原螯虾成熟的标志。
3.4 关于克氏原螯虾雌性生殖系统的超微结构
本文对克氏原螯虾雌性生殖系统的超微结构观察研究表明:在1月份,卵质布满肌层组织结构,含有少量滑面、粗面内质网和线粒体,属于卵巢发育缓慢期;在6月份,克氏原螯虾卵巢含有大量囊泡状滑面内质网和致密小颗粒物质,线粒体正在相互融合、结合,合成大量小颗粒卵黄,属于卵巢快速发育阶段;在9月底,卵质中含量丰富的线粒体、管状粗面内质网、空泡状脂滴以及大的卵黄颗粒,可见高尔基体,卵巢发育接近成熟,卵黄合成基本完成。6月份,在滤泡细胞周边可见排列整齐的长泡状结构,可能与滤泡细胞向卵母细胞输送营养物质有关;在1月份、9月份没有观察到此结构。
3.5 关于克氏原螯虾的产卵问题
克氏原螯虾卵巢成熟系数周年变化表明,克氏原螯虾的性腺指数最高出现在9月份和2月份,这与安徽地区克氏原螯虾分别在10月和4月大量抱卵、抱仔虾基本相符。10月份克氏原螯虾产卵后,进入冬季低温期,卵巢性腺发育缓慢,至4、5月份个体再次产卵的可能性不大。而克氏原螯虾的性腺指数在2月份再次出现峰值,可能与采集样本体质量较小,性腺发育延后,又恰逢低温期,性腺发育缓慢有关。对卵子发生时相的研究表明,每个个体卵巢中卵子的卵母细胞的发育基本一致,没有发现大量处于不同发育阶段的卵母细胞,仅有少量发育之后的卵母细胞,也在卵巢发育过程中逐渐退化。因此,本研究认为,克氏原螯虾个体一年仅产卵1次,一年形成2个产卵高峰,主要由个体性腺发育不同步导致。这与河蟹[20]、南美白对虾[22]、青虾[26]等多次产卵类型不同。
[1]魏青山.武汉地区克氏原螯虾的生物学研究[J].华中农学院学报,1985,4(1):16-24.
[2]方春林,邓勇辉,余智杰,等.克氏原螯虾生物学特性的研究[J].江西水产科技,2010(3):18-20.
[3]谢文星,董方勇,谢山,等.克氏原螯虾的食性、繁殖和栖息习性研究[J].水利渔业,2008,28(4):63-65.
[4]江河,汪留全.克氏原螯虾生物学特性和人工养殖技术[J].齐鲁渔业,2009,19(12):13-15.
[5]雷晓青,肖鸣鹤,胡火庚,等.鄱阳湖克氏原螯虾生物学特性及生长繁殖习性的研究[J].江西水产科技,2009(1):20-21.
[6]龚世园,吕建林,孙瑞杰,等.克氏原螯虾繁殖生物学研究[J].淡水渔业,2008,38(6):23-30.
[7]董卫军,李铭,徐加元.克氏原螯虾繁殖生物学研究[J].水利渔业,2007,27(6):27,104.
[8] Oluoch A O.Breeding biology of the Louisiana red swamp crayfish Procambarus clarkii Girard in Lake Naivasha [J].Kenya Hydrobiologia, 1990, 208: 85-92.
[9]徐增洪,邴旭文,赵朝阳,等.克氏原螯虾卵巢发育与产卵周期变化的观察[J].江苏农业学报,2010,26(6):1304-1307.
[10]吕建林.克氏原螯虾繁殖生物学及胚胎和幼体发育研究[D].武汉:华中农业大学,2006.
[11]戴颖,巩雪洁,李兵,等.武汉地区克氏原螯虾繁殖期的研究[J].动物学杂志,2008,43(2):21-27.
[12]郭风英,周鑫,赵朝阳,等.2种不同体色的克氏原螯虾卵巢发育及生长特性研究[J].海洋湖沼通报,2012(2):89-96.
[13]刘智俊,李建忠,陆锦天,等.上海崇明地区克氏原螯虾卵巢发育规律研究[J].水产科技情报,2017,44(1):1-5.
[14]杨筱珍,王金峰,杨丽娜,等.日本新糠虾(Neomysis japonica)卵巢发育与卵子发生[J].海洋与湖沼,2009,40(3):338-346.
[15]赵云龙,彭欣,夏李祥.罗氏沼虾雌性生殖系统的组织学研究[J].华东师范大学学报:自然科学版,1998(3):81-85.
[16]郑忠明,李明云.哈氏仿对虾卵巢发育的形态学与组织学观察[J].水产学报,2006,26(2):105-110.
[17]栗治国,张成松,李富花,等.脊尾白虾的性腺发育及组织结构观察[J].水产学报,2014,38(3):362-370.
[18]邓道贵.粗糙沼虾雌性生殖系统的组织学研究[J].淮北农师院学报,2000,21(1):49-53.
[19]王春琳,蒋霞敏,赵青松,等.黑斑口虾蛄的卵巢组织学研究[J].动物学杂志,2001,36(4):6-9.
[20]顾志敏,何林岗.中华绒螯蟹卵巢发育周期的组织学细胞学观察[J].海洋与湖沼,1997,28(2):138-145.
[21] King JE.A study of the reproductive organs of common marine shrimp, Penaeussetiferus[J].Biol Bull, 1948, 94: 244-262.
[22]颜素芬,姜永华.南美白对虾卵巢结构及发育的组织学研究[J].海洋湖沼通报,2004(2):52-58.
[23]上官步敏,刘正琮,李少菁,等.锯缘青蟹卵巢发育的组织学观察[J].水产学报,1991,15(2):96-103.
[24]洪水根,林加涵,陈细法,等.长毛对虾卵子发生的研究Ⅰ:卵子发生过程[J].海洋与湖沼,1988,19(4):301-309.
[25] 薛鲁征,堵南山,赖伟.中华绒鳌蟹(Eriocheir sinensis)雌性生殖系统的组织学研究[J].华东师范大学学报:自然科学版,1987(3):88-97.
[26]何緖刚,张训蒲,龚世园,等.武湖日本沼虾卵巢发育研究[J].华东师范大学学报:自然科学版,2002,21(4):148-151.
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21
新医学(2022年4期)2022-04-23
中国畜牧杂志(2022年4期)2022-04-15
中国畜牧杂志(2021年5期)2021-12-05
临床与实验病理学杂志(2021年7期)2021-09-06
湖泊科学(2020年4期)2020-07-17
中国畜牧杂志(2020年6期)2020-07-11
放射学实践(2020年5期)2020-06-01
特产研究(2019年1期)2019-01-13
中西医结合心血管病杂志(电子版)(2018年28期)2018-01-12
