
UV- B辐射增强和氮沉降对不同种源地乌桕叶绿素荧光参数的影响
2018-03-02 08:23:21江亮波邓邦良刘喜帅郑利亚郭晓敏刘苑秋
浙江农业学报 2018年2期
郑 翔,江亮波,邓邦良,刘 倩,刘喜帅,郑利亚,郭晓敏,刘苑秋,张 令
(江西省森林培育重点实验室,江西特色林木资源培育与利用协同创新中心,江西农业大学 林学院,江西 南昌330045)
外来植物入侵严重威胁着生态系统功能和稳定性,已经成为重要的全球性问题[1-2]。随着工业革命的发展,人类活动加剧了全球环境变化,其中就包括UV- B辐射增强[3]和氮沉降加剧[4]。UV- B辐射增强会导致植物DNA同一条链上相邻的两个嘧啶偶联成嘧啶二聚体,造成DNA损伤并抑制细胞分裂以及植株生长,影响资源分配;同时,植物通过“光修复”来分解UV- B辐射形成的嘧啶二聚体,并合成大量的次级代谢产物用于吸收并阻止大部分UV- B辐射进入核DNA、叶绿体、细胞膜[5-7],从而影响植物光合作用或者叶绿素荧光参数等的变化[8-10]。氮沉降加剧则会显著增加植物叶绿素含量,从而影响植物光合作用[11]或者叶绿素荧光参数[12]等的变化。因此,UV- B辐射增强和氮沉降加剧可能会共同影响植物的生长发育,并可能直接影响到入侵植物的建群、扩散和成功入侵等过程。
乌桕(Triadicasebifera)是我国重要的工业油料树种,其种子油脂含量高达40%左右。1772年,为了发展蜡烛和肥皂产业,首次引种进入美国,如今在美国东南部迅速蔓延并呈现出严重的入侵现象[13-15]。乌桕之所以能够成功入侵美国,主要与入侵地乌桕采取“地上优先”的资源分配策略和低抗性、高耐性的进化选择有关[16-17]。先前的研究表明,入侵乌桕种群对UV- B辐射(20%增强)具有显著负效应[18],但入侵乌桕种群相比于本地乌桕种群具有较高的比叶面积、氮吸收速率和相对生长速率等特征[13-14, 19]。先前针对入侵植物乌桕的相关研究,主要是UV- B辐射增强[18],氮沉降增强[14]等单因子研究。然而,UV- B辐射增强和氮沉降加剧二种环境因子对乌桕入侵性的交叉影响研究尚不透彻。本试验采用盆栽试验的方式,通过模拟全球变化因子UV- B辐射增强和氮沉降加剧研究其对不同种源地来源的乌桕叶绿素荧光参数和生长特征的影响,以期探明不同种源地乌桕光合生理对UV- B辐射增强和氮沉降加剧的响应差异及其适应性。
1 材料与方法
1.1 实验材料与实验设计
2014年,在中国本地和美国入侵地各采集6个种群的乌桕种子(每个种群的种子至少采集于3棵树上),共12个乌桕种群(表1)。2015年3月采集乌桕非根际土壤,采集后剔除可见石块、动植物残体等杂物,然后混合均匀,通过四分法选取部分土壤测定土壤理化背景值,结果表明有机碳含量为(4.89±0.15)g·kg-1,总氮含量为(0.47±0.01)g·kg-1,硝态氮含量为(3.39±0.06)mg·kg-1,铵态氮含量为(10.32±0.10)mg·kg-1,有效磷含量为(2.6±0.65)mg·kg-1,pH值5.39±0.02。2015年4月,根据种源分类,将12个种源的乌桕种子分别播种在灭菌的乌桕非根际土壤里,育苗期间通过添加蒸馏水使土壤维持一定湿度,待6月初期乌桕种子萌发并生长至适合移栽时(大约35 d),选择植株高度相近的乌桕幼苗进行移栽并在江西农业大学温室花房中开展相关试验(花盆填装土壤质量为1 kg,体积为1.2 L)。2015年9月初测定叶绿素荧光参数、相对叶绿素含量和株高。
试验采用三因素裂区设计,主区为UV- B辐射(对照 vs. 20% UV- B辐射增强),副区为氮沉降(对照 vs. 8 g N·m-2)×种源(本地乌桕 vs. 入侵乌桕,分别包含6个种群,其中种群作为种源的内部嵌套因子),4次重复,一共192盆乌桕。使用紫外线灯管模拟UV- B辐射[6],白天(07:00—19:00)开启,晚上关闭。使用NH4NO3溶液进行8 g N·m-2的氮沉降模拟(相当于0.03 g N·kg-1土壤,共分3次加入,每次5 mL液体)。
表1不同种群乌桕的地理分布信息
Table1Locations of differentTriadicasebiferapopulations used in the study
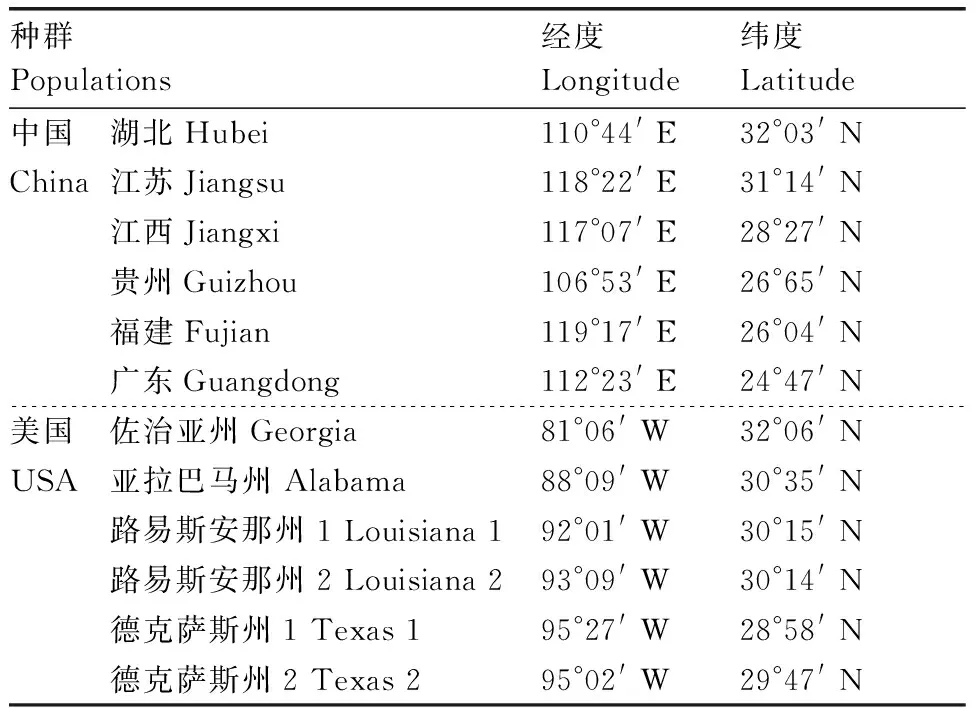
种群Populations经度Longitude纬度Latitude中国湖北Hubei110°44'E32°03'NChina江苏Jiangsu118°22'E31°14'N江西Jiangxi117°07'E28°27'N贵州Guizhou106°53'E26°65'N福建Fujian119°17'E26°04'N广东Guangdong112°23'E24°47'N美国佐治亚州Georgia81°06'W32°06'NUSA亚拉巴马州Alabama88°09'W30°35'N路易斯安那州1Louisiana192°01'W30°15'N路易斯安那州2Louisiana293°09'W30°14'N德克萨斯州1Texas195°27'W28°58'N德克萨斯州2Texas295°02'W29°47'N
1.2 试验方法
1.3 数据处理
采用JMP 9.0 (SAS Institute,Cary,NC,USA)软件进行统计分析,使用LSD(α= 0.05)法进行多重比较,其中种群作为种源的内部嵌套因子进行分析;采用Origin Pro 8.1软件绘图。
2 结果与分析
2.1 株高和SPAD
入侵乌桕较本地乌桕具有较高的株高(+11.26%)(图1- A)。在UV- B辐射条件下,氮沉降对株高具有显著正效应(+6.49%)(图1- B)。
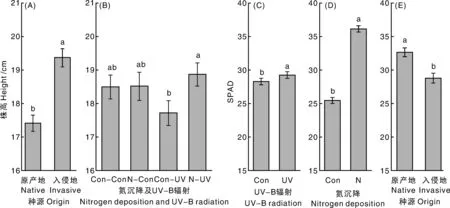
相同字母表示没有显著差异(P>0.05),下同。Data with the same letters were not significantly different in LSD test’s at α=0.05, the same as below.图1 不同处理对乌桕株高和相对叶绿素含量的影响Fig.1 Effect of different treatments on height and SPAD of Triadica sebifera
UV- B辐射对SPAD具有显著正效应(+3.47%)(图1- C)。氮沉降对SPAD具有显著正效应(+42.07%)(图1- D)。本地乌桕相比入侵乌桕具有较高的SPAD(+13.34%)(图1- E)。
2.2 Fv/Fm和Y(Ⅱ)
虽然UV- B辐射对本地乌桕Fv/Fm没有显著影响,但是对入侵乌桕Fv/Fm有显著负效应(-4.00%)(图2- A)。氮沉降对乌桕Y(Ⅱ)具有负效应(-6.4%)(图2- B)。入侵乌桕比本地乌桕具有较高的Y(Ⅱ) (+5.77%)(图2- C)。
2.3 快速光响应曲线(α、ETRmax和Ik)
氮沉降对ETRmax具有显著负效应(-12.32%)(图3),但UV- B辐射和氮沉降对α和Ik都没有显著影响(表2)。
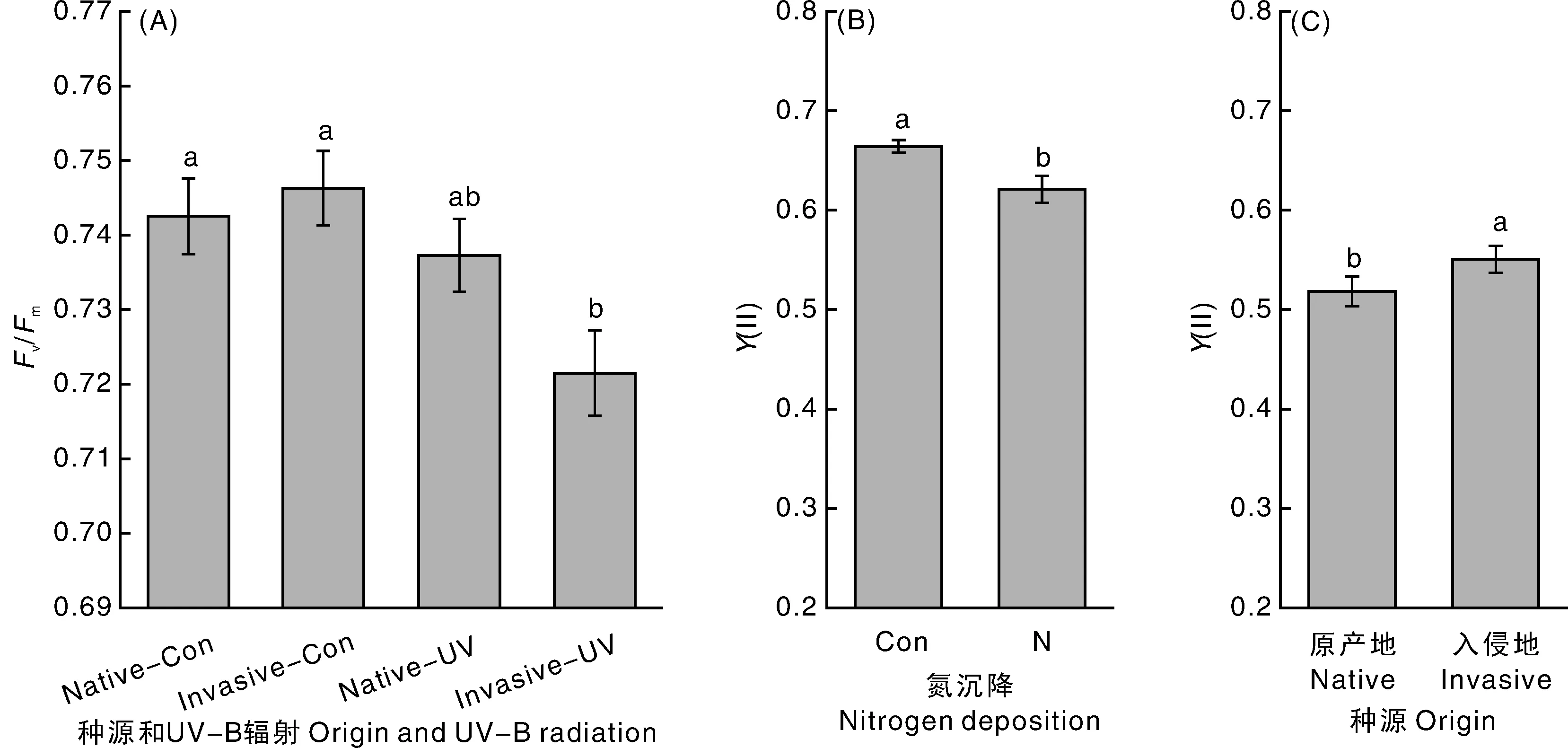
图2 不同处理对乌桕光系统Ⅱ最大量子产率和光系统Ⅱ实际光合量子产率的影响Fig.2 Effect of different treatments on Fv/Fm and Y(Ⅱ) of Triadica sebifera
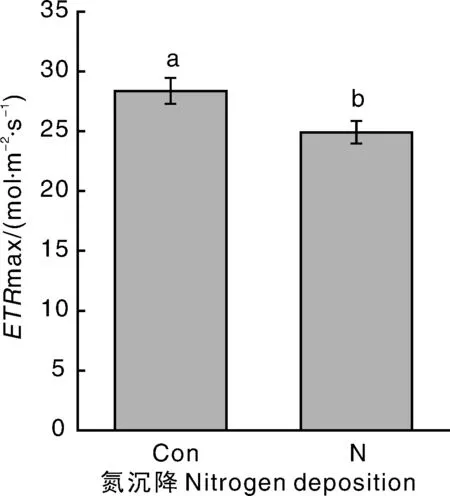
图3 不同处理对乌桕潜在最大相对电子传递速率的影响Fig.3 Effect of different treatments on ETRmax of Triadica sebifera
3 讨论
3.1 氮沉降能减缓UV- B辐射对乌桕株高生长的胁迫作用
株高受到UV- B辐射和氮沉降的交互作用影响(表2)。虽然UV- B辐射或者氮沉降处理与对照处理的乌桕株高无显著差异,但是UV- B辐射和氮沉降共同处理的株高显著高于UV- B辐射处理(+6.49%)(图1- B),这意味着氮沉降能减缓UV- B辐射对乌桕株高生长的胁迫作用。这可能是因为N的增加为乌桕合成用于吸收并阻止UV- B辐射的次级代谢产物提供了养分[5-7],由此我们推测UV- B辐射增强可能会影响植物对土壤养分的吸收或者利用能力,从而具有潜在影响植物入侵的能力。
3.2 UV- B辐射增加乌桕SPAD,但氮沉降对乌桕生长的影响具有多样性
UV- B增强降低了杨桐幼苗的SPAD(21个月和24个月后)[9];对乐东拟单性木兰、青冈和山核桃无影响,但相比于UV- B减弱处理,对照促进了乐东拟单性木兰的SPAD[10],而韩雯等[22]的研究结果则表明拟南芥幼苗叶绿素a、叶绿素以及叶绿素总含量(SPAD与叶绿素含量密切相关)
表2UV- B辐射和氮沉降对不同种源乌桕叶绿素荧光参数(Fv/Fm、Y(Ⅱ)、α、Ik、ETRmax)、相对叶绿素含量和株高的影响方差分析
Table2Effect of UV- B radiation (UV) and nitrogen deposition (N) on chlorophyll fluorescence parameters (Fv/Fm,Y(Ⅱ),α,Ik,ETRmax),SPADand height ofTriadicasebiferaorigin (O) and their interactions in ANOVAs

TreatmentdfFv/FmY(Ⅱ)α/(electrons·photons-1)ETRmax/(μmolelectrons·m-2·s-1)Ik/(μmol·m-2·s-1)SPADHeight/cmFvalueFvalueFvalueFvalueFvalueFvalueFvalueUV110.247**0.0041.3190.6792.9704.528*0.301N10.21025.151**0.0387.290*1.960182.132**1.724O10.8124.392*0.6670.0180.02926.143**40.229**UV-N10.6111.3410.0280.0062.7380.0324.490*UV-O16.058*2.6260.3581.5090.4630.1250.147N-O13.6330.0150.4480.0190.5770.3320.798UV-N-O10.4610.0381.7982.6720.1450.0120.083
UV, UV- B辐射; N, 氮沉降; O, 种源;*表示差异显著(0.01 UV, UV- B radiation; N, nitrogen deposition; O,Triadicasebiferaorigin; * , 0.01 对UV- B辐射胁迫响应具有时间效应,随着胁迫时间的延长其叶绿素a、叶绿素b以及叶绿素总含量呈现先上升后下降趋势。由此可见,植物叶片SPAD对UV- B胁迫响应既与植物种类相关,也取决于胁迫时间。在本实验中,UV- B辐射促进了乌桕叶片SPAD(图1- C),但在我们的研究中并未就不同胁迫时间叶片SPAD的变化进行测定,因此,在今后的研究中还需适当延长处理和测定时间,充分考虑SPAD对UV- B辐射的时间效应。 人为添加氮肥能显著增加小麦[23]和糜子[24]叶片SPAD,都与本试验结果相一致(图1- D),说明随着氮源的输入,植物叶片的叶绿素含量增加,叶片SPAD值上升。张绪成等[12]却发现施加氮肥能显著增加小麦叶片Y(Ⅱ),与本研究结果相反(图2- B),但是鲍海泳等[25]对入侵植物薇甘菊的研究表明,氮沉降对Y(Ⅱ)无影响,宫兆宁等[26]研究水氮含量对挺水植物(芦苇、香蒲等)Y(Ⅱ)的影响表明:随着水氮含量增加,Y(Ⅱ)先升高(<15 mg·L-1), 后持平(15~20 mg·L-1),因而,氮沉降对植物Y(Ⅱ)的影响可能与供试植物的归化程度以及土壤氮水平的共同作用有关。 虽然UV- B辐射对本地乌桕Fv/Fm没用显著影响,但是对入侵乌桕Fv/Fm有显著负效应(-4.00%)(图2- A),表明入侵乌桕比本地乌桕对UV- B辐射胁迫更敏感,换言之,本地乌桕相比于入侵乌桕具有较强的UV- B辐射抵御能力。本地和入侵植物对UV- B辐射胁迫的适应策略和响应机制有所不同,可能与本地和入侵植物对UV- B辐射胁迫的抵御和修复能力各异有关[5, 18, 27-28],因此,UV- B辐射增强可能直接影响入侵植物的建群、扩散和成功入侵等过程。本实验中,入侵乌桕相比于本地乌桕具有较高的Y(Ⅱ)(图2- C)和株高(图1- A),表明入侵乌桕相比本地乌桕具有较高的实际光能转化效率和株高生长速率,这可能是入侵乌桕在入侵地进化出了较高的植株生长性能导致的[29]。先前研究结果表明,入侵乌桕种群采取“地上优先”的分配策略和高耐性、低抗性的进化选择[16-17],使其相比于本地乌桕种群具有较高的比叶面积、氮吸收速率和相对生长速率等特征[13-14, 19],从而在植株生长等方面进化出了较为明显的优势。本研究结果同样表明,入侵乌桕在进化的过程中,资源分配发生了改变,将大部分的资源分配给了植物生长而不是抵御外界UV- B辐射逆环境,使得入侵乌桕具有较高的株高生长优势、光能转化效率和较低的UV- B辐射抗逆性,从而可能导致入侵乌桕对UV- B辐射增强和氮沉降加剧构成的复杂环境的响应及适应机制发生变化[29-33]。 全球变化因子UV- B辐射增强和氮沉降加剧组成的复杂环境可能会通过影响植物光合作用效率进而共同影响乌桕生长乃至成功入侵过程。氮沉降能减缓UV- B辐射对乌桕株高生长的胁迫作用,入侵乌桕种群相比于本地乌桕种群,在进化过程中对环境因子的响应和适应机制发生了改变,将更多的资源分配给了植物生长而不是抵御外界逆环境,导致入侵乌桕种群具有较高的实际光能转化效率和株高生长速度并对UV- B辐射胁迫更敏感,从而具有顺利生长、建群和扩散的潜力。 [1] 陈宝明, 彭少麟, 吴秀平, 等. 近20年外来生物入侵危害与风险评估研究的文献计量分析[J]. 生态学报, 2016, 36(20): 1-9. CHEN B M, PENG S L, WU X P, et al. A bibliometric analysis of researches on topics related to the ecological damage caused by and risk assessments of exotic invasive species from 1995 to 2014[J].ActaEcologicaSinica, 2016, 36(20): 1-9. (in Chinese with English abstract) [2] 吴昊. 陆生生境入侵植物空心莲子草群落数量分类与排序[J]. 浙江农业学报, 2017,29(3): 433-444. WU H. Numerical classification and ordination of invasive plantAlternantheraphiloxeroidescommunity in terrestrial habitats[J].ActaAgriculturaeZhejiangensis, 2017, 29 (3): 433-444. (in Chinese with English abstract) [3] WILLIAMSON C E, ZEPP R G, LUCAS R M, et al. Solar ultraviolet radiation in a changing climate[J].NatureClimateChange, 2014, 4(6): 434-441. [4] KANAKIDOU M, MYRIOKEFALITAKIS S, DASKALAKIS N, et al. Past, present, and future atmospheric nitrogen deposition[J].JournaloftheAtmosphericSciences, 2016, 73(5): 2039-2047. [5] 李良博, 唐天向, 海梅荣, 等. 植物对UV- B辐射增强的响应及其分子机制[J]. 中国农学通报, 2015, 31(13): 159-163. LI L B, TANG T X, HAI M R, et al. Response and molecular mechanisms of plants to enhanced UV- B radiation[J].ChineseAgriculturalScienceBulletin, 2015, 31(13): 159-163. (in Chinese with English abstract) [6] 倪郁, 宋超, 李加纳. UV- B辐射增强对拟南芥表皮蜡质的影响[J]. 生态学报, 2015, 35(5): 1505-1512. NI Y, SONG C, LI J N. Effect of enhanced ultraviolet- B radiation on epicuticular wax inArabidopsisthaliana[J].ActaEcologicaSinica, 2015, 35(5): 1505-1512. (in Chinese with English abstract) [7] SUCHAR V A, ROBBERECHT R. Integration and scaling of UV- B radiation effects on plants: from molecular interactions to whole plant responses[J].EcologyandEvolution, 2016,6(14):4866-4884. [8] 师生波, 尚艳霞, 朱鹏锦, 等. 短期UV- B辐射对青藏高原美丽风毛菊PSⅡ光化学效率的影响[J]. 应用生态学报, 2011, 22(5): 1147-1154. SHI S B, SHANG Y X, ZHU P J, et al. Effects of short term enhanced UV- B radiation on the PSⅡ photochemical efficiency of alpine plantSaussureasuperba[J].ChineseJournalofAppliedEcology, 2011, 22(5): 1147-1154. (in Chinese with English abstract) [9] 兰春剑, 江洪, 黄梅玲, 等. UV- B辐射胁迫对杨桐幼苗生长及光合生理的影响[J]. 生态学报, 2011, 31(24): 7516-7525. LAN C J, JIANG H, HUANG M L, et al. The growth and photosynthetic responses ofCleyerajaponicaThunb seedlings to UV- B radiation stress[J].ActaEcologicaSinica, 2011, 31(24): 7516-7525. (in Chinese with English abstract) [10] 黄梅玲, 江洪, 金清, 等. UV- B辐射胁迫下不同起源时期的3种木本植物幼苗的生长及光合特性[J]. 生态学报, 2010, 30(8): 1998-2009. HUANG M L, JIANG H, JIN Q, et al. The seeding growth and photosynthetic characteristic of three ligneous plants with different evolutionary time to UV- B radiation stress[J].ActaEcologicaSinica, 2010, 30(8): 1998-2009. (in Chinese with English abstract) [11] 黄长兵, 房伟民, 杨勇, 等. 不同水平和形态氮素供应对盆栽小菊外观品质和光合特性的影响[J]. 浙江农业学报, 2010,22(1): 45-50. HUANG C B, FANG W M, YANG Y, et al. Effects of different nitrogen levels and forms on the appearance quality and photosynthesis characters of potted chrysanthemum with small in florescences[J].ActaAgriculturaeZhejiangensis, 2010, 22(1): 45-50. (in Chinese with English abstract) [12] 张绪成, 于显枫, 马一凡. 施氮和大气CO2浓度升高对小麦旗叶光合电子传递和分配的影响[J]. 应用生态学报, 2011, 22(3): 673-680. ZHANG X C, YU X F, MA Y F. Effects of nitrogen application and elevated atmospheric CO2on electron transport and energy partitioning in flag leaf photosynthesis of wheat[J].ChineseJournalofAppliedEcology, 2011, 22(3): 673-680. (in Chinese with English abstract) [13] YANG Q, LI B, SIEMANN E. The effects of fertilization on plant- soil interactions and salinity tolerance of invasiveTriadicasebifera[J].PlantandSoil, 2015, 394(1/2): 99-107. [14] ZHANG L, ZHANG Y, WANG H, et al. Chinese tallow trees (Triadicasebifera) from the invasive range outperform those from the native range with an active soil community or phosphorus fertilization[J].PLoSOne, 2013, 8(9): e74233. [15] HUANG W, CARRILLO J L, DING J Q, et al. Interactive effects of herbivory and competition intensity determine invasive plant performance[J].Oecologia, 2012, 170(2): 373-382. [16] ZOU J W, SIEMANN E, ROGERS W E, et al. Decreased resistance and increased tolerance to native herbivores of the invasive plantSapiumsebiferum[J].Ecography, 2008, 31(5): 663-671. [17] 邓邦良, 刘倩, 刘喜帅, 等. UV- B辐射增强和氮沉降对入侵植物乌桕生长的影响[J]. 植物生态学报, 2017, 41(4): 471-479. DENG B L, LIU Q, LIU X S, et al. Effects of enhanced UV- B radiation and nitrogen deposition on the growth of invasive plantTriadicasebifera[J].ChineseJournalofPlantEcology, 2017, 41(4): 471-479. (in Chinese with English abstract) [18] WANG H, MA X C, ZHANG L, et al. UV- B has larger negative impacts on invasive populations ofTriadicasebiferabut ozone impacts do not vary[J].JournalofPlantEcology, 2016, 9(1): 61-68. [19] ZOU J, ROGER W E, SIEMANN E. Differences in morphological and physiological traits between native and invasive populations ofSapiumsebiferum[J].FunctionalEcology, 2007, 21(4): 721-730. [20] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000: 1-638. [21] 叶子飘, 胡文海, 肖宜安, 等. 光合电子流对光响应的机理模型及其应用[J]. 植物生态学报, 2014, 38(11): 1241-1249. YE Z P, HU W H, XIAO Y A, et al. A mechanistic model of light- response of photosynthetic electron flow and its application[J].ChineseJournalofPlantEcology, 2014, 38(11): 1241-1249. (in Chinese with English abstract) [22] 韩雯, 韩榕. 不同时间的UV- B辐射对拟南芥幼苗生长的影响[J]. 植物学报, 2015, 50(1): 40-46. HAN W, HAN R. Effect of different times of UV- B rRadiation on seedling growth ofArabidopsisthaliana[J].ChineseBulletinofBotany, 2015, 50(1): 40-46. (in Chinese with English abstract) [23] 赵犇, 姚霞, 田永超, 等. 基于上部叶片SPAD值估算小麦氮营养指数[J]. 生态学报, 2013, 33(3): 916-924. ZHAO B, YAO X, TIAN Y C, et al. Estimation of nitrogen nutrient index on SPAD value of top leaves in wheat[J].ActaEcologicaSinica, 2013, 33(3): 916-924. (in Chinese with English abstract) [24] 张盼盼, 周瑜, 宋慧, 等. 不同肥力水平下糜子生长状况及农田小气候特征比较[J]. 应用生态学报, 2015, 26(2): 473-480. ZHANG P P, ZHOU Y, SONG H, et al. Comparison of growth and field microclimate characteristics of broomcorn millet under different fertilization conditions[J].ChineseJournalofAppliedEcology, 2015, 26(2): 473-480.(in Chinese with English abstract) [25] 鲍海泳, 肖辉林, 陈红跃, 等. 模拟氮沉降对入侵植物薇甘菊叶绿素荧光参数的影响[J]. 生态环境学报, 2013, 22(12): 1867-1871. BAO H Y, XIAO H L, CHEN H Y, et al. Effects of simulated nitrogen deposition on the chlorophyll fluorescence parameters of the invasive plantMikaniamicrantha[J].EcologyandEnvironmentalSciences, 2013, 22(12): 1867-1871.(in Chinese with English abstract) [26] 宫兆宁, 范云豹, 刘辉, 等. 不同水氮梯度下典型挺水植物叶绿素荧光的响应特性[J]. 植物学报, 2016, 51(5): 631-638. GONG Z N, FAN Y B, LIU H, et al. Chlorophyll fluorescence response characteristics of typical emergent plants under different total nitrogen gradient[J].ChineseBulletinofBotany,2016, 51(5): 631-638.(in Chinese with English abstract) [27] HOCK M, BECKMANN M, HOFMANN R R, et al. Effects of UV- B radiation on germination characteristics in invasive plants in New Zealand[J].NeoBiota, 2015, 26: 21-37. [28] 李良博, 张连根, 唐天向, 等. UV- B辐射增强对紫茎泽兰和艾草抗性生理特性的影响[J]. 西北植物学报, 2016, 36(2): 343-352. LI L B, ZHANG L G, TANG T X , et al. Influence of enhanced UV- B on physiological characteristics ofEupatoriumadenophorumL. andArtemisiaargyiLevl[J].ActaBotanicaBoreali-OccidentaliaSinica, 2016, 36(2): 343-352.(in Chinese with English abstract) [29] BLOSSEY B, NOTZOLD R. Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis[J].JournalofEcology, 1995, 83(5): 887-889. [30] HENERY M L, BOWMAN G, MRZ P, et al. Evidence for a combination of pre- adapted traits and rapid adaptive change in the invasive plantCentaureastoebe[J].JournalofEcology, 2010, 98(4): 800-813. [31] 刘建, 李钧敏, 余华, 等. 植物功能性状与外来植物入侵[J]. 生物多样性, 2010, 18(6): 569-576. LIU J, LI J M, YU H, et al. The relationship between functional traits and invasiveness of alien plants[J].BiodiversityScience, 2010, 18(6): 569-576.(in Chinese with English abstract) [32] BRADLEY B A, BLUMENTHAL D M, WILCOVE D S, et al. Predicting plant invasions in an era of global change[J].TrendsinEcology&Evolution, 2010, 25(5): 310-318. [33] 黄伟, 王毅, 丁建清. 入侵植物乌桕防御策略的适应性进化研究[J]. 植物生态学报, 2013, 37(9): 889-900. HUANG W, WANG Y, DING J Q. A review of adaptive evolution of defense strategies in an invasive plant species, Chinese tallow (Triadicasebifera)[J].ChineseJournalofPlantEcology, 2013, 37(9): 889-900.(in Chinese with English abstract)3.3 入侵乌桕较之本地乌桕具有较低的UV- B抗性和较高的株高生长优势、光能转化效率
猜你喜欢
林业与生态(2022年5期)2022-05-23 01:16:51花卉·上半月(2022年9期)2022-04-29 00:44:03阅读(科学探秘)(2020年8期)2020-11-06 06:22:48农村百事通(2019年17期)2019-10-08 02:24:55中国果业信息(2019年1期)2019-01-05 17:41:42现代园艺(2017年21期)2018-01-03 06:42:18生物学教学(2017年9期)2017-08-20 13:22:32浙江农业科学(2017年5期)2017-06-21 15:12:46现代农业科技(2017年1期)2017-03-06 23:08:18江苏农业科学(2016年8期)2017-02-15 19:19:29
