
与鱼道水力设计相关的草鱼幼鱼游泳行为特性研究
2018-01-21 07:17穆祥鹏白音包力皋王秀英陈云飞
水利学报 2017年12期
曹 平,穆祥鹏,白音包力皋,王秀英,陈云飞
(中国水利水电科学研究院 流域水循环模拟与调控国家重点实验室,北京 100038)
1 研究背景
鱼类洄游是因其生长发育和外界环境变化的需要而在长期适应过程中所获得的特性,是一种在越冬场、产卵场和索饵场之间的周期性、定向性和集群性的迁移活动。“四大家鱼”青、草、鲢、鳙是我国主要的经济鱼种,属于典型的江(河)湖半洄游性鱼类,平时在通江湖泊中摄食育肥,繁殖季节结群逆水洄游到江(河)干流各产卵场生殖,栖息在江河下游的亲鱼也会洄游到中游或上游产卵场产卵。产后的亲鱼、幼鱼洄游到食料丰盛的河湾、支流或通江湖泊中索饵育肥。冬季湖水降落,鱼群又回到江(河)干流深水区或较深的岩坑处越冬[1]。
近几十年来,我国大坝、闸堰等水利工程大量兴建,鱼类“三场”之间的洄游通道被人为阻断,干流内的家鱼不能进入通江湖泊或支流摄食、育肥,湖泊中的家鱼不能进入干流越冬、繁殖,导致野生“四大家鱼”种群数量持续下降。2013年长江三峡工程生态与环境监测公报显示[2],2012年5—7月,三峡坝下监利江段四大家鱼鱼苗径流量平均值仅为蓄水前(1997—2002年)的3.8%。保护野生家鱼种群、恢复鱼类洄游通道已刻不容缓。鱼道是鱼类通过溯河障碍物的重要通路,其水力学设计必须与鱼类游泳行为特性相适应。国内外普遍认同的表征鱼类游泳行为特性的指标主要有临界游泳速度、突进游泳速度和感应流速。针对四大家鱼开展相关研究,对指导鱼道工程水力设计是极其重要的。
以往四大家鱼游泳能力的研究多集中于临界游泳速度,其反映了鱼类长时间的游泳能力,是设计鱼道池室主流、确定休息池间距的重要指标。鲜雪梅等[3]对体长7.93±0.08 cm青鱼的临界游泳速度进行了试验,得到青鱼在25℃时的相对临界游泳速度为5.25±0.18BL/s(BL代表鱼体长度)。房敏等[4]对体长11.35±0.45 cm鲢在不同温度下的临界游泳速度进行了试验,得到25℃条件下鲢的相对临界游泳速度为6.85±0.16BL/s。蔡露等[5]对体长9.8~12.3 cm鳙的临界游泳速度进行了试验,得到的鳙在20℃时的相对临界游泳速度为4.57±0.56BL/s。龚丽等[6]对体长5.0~15.0 cm的草鱼临界游泳速度进行了研究,发现其临界游泳速度随鱼体长增加而线性增长,临界游速在68.0~100.0 cm/s之间。
鱼类能否通过鱼道高流速区主要与其突进游泳速度相关。熊锋等[7]采用递增流速法研究了松花江流域体长20cm左右的四大家鱼突进游泳速度,发现其相对突进游泳速度大小排序为鲢>草>青>鳙。此外黎采微[8]、牛宋芳[9]、路波等[10]采用惊吓方法分别测量了不同体长青鱼 (9.21±1.89 cm、17.83±2.67 cm、61.45±0.8 cm)、鲢鱼(体长1.55±0.25 cm、20.05±2.05 cm、43.19±4.26 cm)、草鱼(体长8.47±0.73 cm、17.93±1.27 cm、51.42±3.24 cm)在快速启动过程中的加速-滑行游泳行为,发现绝对最大疾冲速度随体长的增加而增加,相对最大疾冲速度随体长的增加而减小,试验鱼达最大疾冲速度后,均以身体保持直线的方式减速滑行。
感应流速是鱼能够辨别水流方向的最小流速。对于“四大家鱼”感应流速的研究较少,赵希坤等[11]曾测定过体长15~20 cm的草鱼和10~25 cm的鲢鱼的感应流速,两种鱼的感应流速均约为0.2 m/s。青鱼、鳙鱼的感应流速研究尚未见报道。
从以往的研究成果来看,四大家鱼游泳特性和游泳能力的研究主要集中在临界游泳速度方面,突进游泳速度和感应流速的研究成果较少,仅有的一些研究成果也是针对体长>15 cm的鱼类开展的,体长5~15 cm幼鱼突进游速和感应流速的相关研究尚未见报道。胡茂林[12]调查发现四大家鱼幼鱼索饵洄游进入鄱阳湖口的时间主要集中在7月中旬至8月底,湖口平均水温约28~30℃左右,幼鱼体长集中在5.00~14.99 cm,所以开展该体长范围内幼鱼突进游速和感应流速研究对于长江家鱼资源保护更具有实际意义。而且鱼类游泳能力与其体长成正比,幼鱼游泳能力相对较弱,开展幼鱼相关游泳特性指标的研究,对于改善或消除鱼道水流障碍、指导鱼道水力设计是尤为重要的。
本文在充分调研国内外鱼类游泳行为特性和游泳能力研究的基础上,利用鱼类游泳行为试验装置,设计相应的游泳行为特性指标的测试方法,针对5~16 cm草鱼幼鱼,重点测定其感应流速和突进游速,并分析鱼类游泳行为对于快速变化的水流流速的响应,研究成果补充了四大家鱼游泳特性指标,为鱼道水力学研究和设计提供基本参数。
2 试验设计
2.1 鱼类游泳行为试验水槽鱼类游泳能力和行为特性指标一般通过密闭空间中流速可调的均匀流场来测定。试验过程中认为鱼类游泳速度与水流速度相等,通过一定时间内递增流速,观测试验鱼的游泳行为。本研究采用的鱼类游泳行为试验装置见图1。该装置是一个近似椭圆形的密闭循环水槽,通过变频电机带动螺旋桨在水槽内制造出循环水流。通过变频控制器来调节电机转速,以控制水槽流速。密闭水槽两侧弯道处设圆弧形导流板,试验段上游侧设置管簇整流栅,从而在试验段获得均匀、稳定的流场。试验水槽截面25 cm×25 cm,试验段长80 cm,试验段上下游均设置拦网,以保证试验鱼始终在试验段内游泳。装置整体采用有机玻璃制作,方便观测鱼类游泳行为。试验段正上方和正立面各安置一个摄像机,记录鱼类游泳试验的全过程。
采用毕托管对试验段水流流速进行标定,在标定断面上横向选取左、中、右3个测流垂线,每个测流垂线上每隔5 cm水深设置一个流速测点,断面流速取各流速测点的平均值,并分析试验段断面流速的均匀性。
水流分布不均匀系数采用以下公式进行计算:
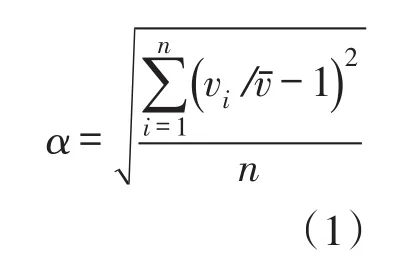
式中:vi为各测点的水流流速,m/s;vˉ为断面平均水流流速,m/s;n为断面的测点个数。
根据式(1)计算出各频率下的水流分布不均匀性系数的平均值为0.017,系数较小,说明试验段的水流均匀性良好。试验水槽的流速调节范围为0.00~1.50 m/s,实验段流速与变频器输出频率Hz的关系如图2所示。
有研究表明,当试验鱼体的横截面小于试验水槽的横截面积的10%时,试验鱼对水流的阻挡效应可忽略不计[13]。本试验水槽横截面积、试验段长度尺寸均较大,横断面流速分布均匀,开展草鱼幼鱼游泳试验能够获得理想的试验结果。
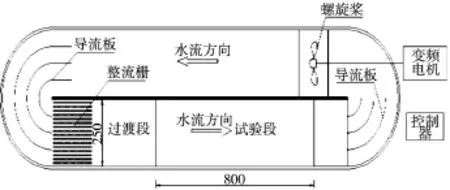
图1 鱼类游泳能力试验装置(单位:mm)

图2 试验段水流速度与调控频率的关系

图3 试验草鱼体长与体重关系
2.2 试验用鱼试验用草鱼取自北京某渔场,采集时间为8月中旬,试验鱼体长范围:5.0~16.0 cm;试验鱼体重范围:3.80~39.08 g。鱼体长与鱼体重的关系如图3所示。根据鱼体长-体重关系式:W=bLa[14]推出试验草鱼体长(L)与体重(W)的关系式为:W=0.04L2.49(R2=0.96)。本试验用鱼a=2.49<3,呈强异速性生长[15]。
2.3 试验条件胡茂林对四大家鱼幼鱼的调查[12]表明,家鱼幼鱼进入鄱阳湖的索饵洄游期为7月中旬至8月底,本次试验时间为8月下旬。按照鱼类生理试验的驯化和暂养要求,将试验鱼放置于长宽2.0 m×1.5 m的矩形水池中暂养2周,水深1.0 m,暂养水为曝气5 d的自来水,水温为25±1℃,溶解氧浓度维持在6.0 mg/L以上,满足草鱼幼鱼正常生长需求[16],光照为室内自然光。试验前2天停止喂食,正式试验时,水温为25±0.5℃,溶解氧浓度维持在6.0 mg/L以上,为了避免由于日照产生的温差,试验选择在室内进行。暂养与试验水温、溶解氧条件与7—8月家鱼鱼苗索饵洄游的自然水环境条件基本一致。
试验开始前的水流适应条件对鱼类游泳能力的准确测定非常重要。为了避免鱼类在试验装置中不适应,导致生理应激而致试验失败,需要在试验开始前,让试验鱼在试验装置中适应一段时间。徐革峰等[17]通过调整适应流速、适应时间、流速递增时间步长和递增幅度来研究适应条件对细鳞鲑幼鱼最大持久游泳时间和临界游泳速度的影响,发现适应流速为1.0~1.5BL/s条件下的最长适应时间不应超过1 h,试验开始前试验鱼适应环境的时间设定为1 h最佳,最多不应超过2 h。针对该研究成果,并参考国内外鱼类游泳能力试验方法,在进行突进游速试验前,使试验鱼在1BL/s的流速下适应1 h;在感应流速试验前,使试验鱼在0流速下适应1 h。
2.4 游泳行为特性测定方法(1)感应流速测定方法:鱼类在游泳时通过水流来感知方向,感应流速指能够使鱼类产生趋流反应的流速值,在试验时通常以不断增加流速使鱼类游动方向朝向来流方向为指示标准。在鱼道的设计过程中,感应流速是一个重要参数,它反映鱼道以及鱼类洄游路线中鱼类可以感应到的最小流速,在这个流速以上,鱼类会较易感知来流而确认洄游方向。
试验时取30尾暂养24 h后的试验鱼,分为3组,每组10条,置于游泳能力测试水槽中,在静水中适应1 h后逐步调大试验流速,同时观察鱼的游泳行为,直至超过半数的试验鱼掉转方向逆流游动,以此时的流速作为试验鱼的感应流速[18]。
(2)突进流速测定方法:突进游泳速度指鱼类在较短时间内(<20 s)达到的最大游泳速度,一般以其作为评价鱼类能否通过水流障碍的重要指标。美国TRB2009年会报告[19]指出:鱼类通过鱼道孔口或窄缝时的游泳速度为突进游泳速度;Blake[20]通过研究发现鱼类通过竖缝式鱼道的竖缝时运用突进游泳速度,直到疲劳才停下来休息,并认为突进游速是鱼道内一些特殊结构及高流速区的水力设计依据。
本研究中突进流速采用“递增流速法”测定,该方法相对耗时较短,流速可控性强,且得到统计学上有意义的结果值所需的试验鱼数目较少,因而应用最为广泛。试验时首先对暂养24 h后的某一体长范围内试验鱼进行2次突进游速预估试验,得到其突进游速预估值,再根据预估值确定试验的速度增量。具体方法如下:
突进游速预估试验时,每次选取1尾鱼放入试验段,试验前使试验鱼在1BL/s的流速下适应1 h,以消除转移过程对鱼体的胁迫。然后每隔20 s使试验流速增加0.4BL/s,直至鱼疲劳,记录此时的流速值,利用式(2)得到突进游速的预估值。将2次预估值的平均值作为最终预估值,供正式试验参考。试验鱼疲劳的判断标准为:试验鱼被水流冲至试验段下游拦网上无法游动的时间超出20 s。
正式试验时,将同体长范围的单尾试验鱼放入试验装置,使其在1BL/s的流速下适应1 h后开始试验。首先将流速在10 s内增至60%的突进游泳速度预估值,然后每隔20 s将流速增加15%的突进游速预估值,同时观察记录鱼的游泳行为,直至试验鱼疲劳无法继续游动,此时结束试验,记录此时水流速度和游泳时间,通过式(2)计算突进游速。
突进游速按式(2)计算:

式中:Uburst为突进游泳速度,cm/s;U为试验鱼疲劳之前的水流速度,cm/s;ΔU为流速增量,即15%的突进游速预估值 ,cm/s;t为该时段鱼疲劳时所用的时间,s;Δt为流速增加的时间间隔,s。
相对突进游速计算公式如下:

式中:U′burst为相对临界游泳速度,BL/s;BL为鱼类体长,cm。
3 试验结果与分析
3.1 感应流速用于感应流速测定的试验用鱼共30尾,每组试验10尾,分成3组。试验鱼体长范围10.31~1 6.01 cm,体重范围12.16~39.08 g。其中第一组试验用鱼体长范围:12.01~15.00 cm,体重:17.10~36.05 g,测得感应流速为12.25 cm/s。第二组试验用鱼体长范围:11.30~14.60 cm,体重:17.49~34.23 g,测得感应流速为10.72 cm/s。第三组试验鱼的体长范围:10.30~16.01 cm,体重:12.16~39.08 g,测得感应流速为15.55 cm/s。试验结果见表1。

表1 草鱼幼鱼的感应流速
通过上述试验可知,体长10.31~16.01 cm,体重12.16~39.08 g的草鱼幼鱼的感应流速范围为10.72~15.55 cm/s,平均感应流速为12.84 cm/s。在进行鱼道水流设计时,要确保池室内的主流流速大于感应流速,这样鱼类上溯时容易感知游泳方向。
3.2 突进游速突进游速正式实验共用草鱼样本30尾,鱼体长范围在6.00~15.00 cm之间,鱼重3.80~30.01 g。试验得到鱼体长与绝对突进游泳速度的关系如图4所示。由图4可以看出草鱼幼鱼绝对突进游泳速度随着草鱼体长的增加而增加,两者基本呈线性相关,由此得到的线性拟合关系式如下。
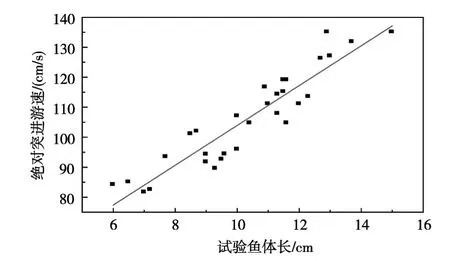
图4 草鱼幼鱼绝对突进游泳速度与鱼体长的关系

图5 草鱼幼鱼相对突进游泳速度与鱼体长的关系

鱼体长和相对突进游泳速度的关系如图5,由图5可以看出相对突进游泳速度随鱼体长的增加而减小,其拟合公式如下:

3.3 草鱼幼鱼临界游速与突进游速的关系龚丽等[6]采用同一试验装置,利用递增流速法对体长在5.00~15.00 cm范围内的草鱼幼鱼临界游泳速度进行了测定,试验时每隔5 min增加0.5BL/s的流速至60%的预估临界流速值,然后每隔20 min增加15%的预估临界流速值,直至试验草鱼疲劳(试验水温28±1℃,溶解氧浓度大于6.5 mg/L),得到的临界游速与鱼体长的关系如下:

式中Ucrit为临界游泳速度,cm/s。
水温是影响鱼类游泳能力的主要因素[21-22]。不同水温下,鱼类代谢能力不同[23-24],继而造成鱼类游泳能力不同。有研究表明鱼类临界游泳速度与水温呈“钟形”或“线形”关系,而突进游泳速度反映的是鱼躲避敌害、穿越障碍的应激能力,通常只与鱼种及体长相关[25-26]。目前关于草鱼幼鱼临界游泳速度与水温的定量关系尚未明确,但草鱼属于温水鱼,而温水鱼的临界游泳速度的最大值出现在25 ~30℃[27-28]。从以上这些研究成果看,可以认为龚丽所做的草鱼幼鱼临界游泳速度试验条件与本次突进游速试验条件是相似的,可以将二者的试验结果进行比较。
结合本次试验得出的草鱼幼鱼突进游泳速度公式,可以得出体长在5.0~15.0 cm范围内的草鱼幼鱼绝对临界游速与绝对突进游速的关系如下(单位同上):
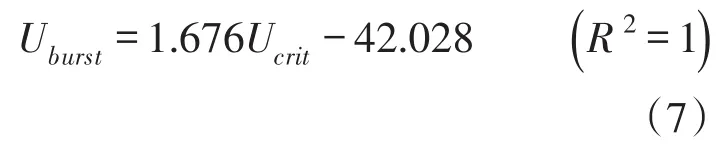
对于体长5.0~15.0 cm范围的草鱼幼鱼,其突进游速比临界游速大6.14~30.2 cm/s,突进游速约为临界游速的1.05~1.28倍,而且鱼的体长越长,突进游速超出临界游速就越多。由式(4)减去式(6)可得到突进游速与临界游速的差值,如下式所示:

3.4 草鱼幼鱼的突进游泳行为与流速的关系在测定鱼类的突进游速时,将流速在10 s内增至60%的突进游泳速度预估值,然后每隔20 s将流速增加15%的突进游速预估值,通过快速而频繁的增加流速,迫使鱼体迅速调整自己的身体机能以适应过快的流速变化,使鱼体始终处于胁迫和紧张状态,以模拟鱼类通过水流障碍时的爆发式游泳行为。
鱼类游泳行为是一种状态不稳定的运动,其游泳行为随着流速的变化而变化,阶段性持续式游泳、暂停及偶发性的冲刺游泳运动相互穿插。
本研究通过视频回放观察预实验及正式试验的草鱼游泳录像,分析了草鱼幼鱼在不同水流速度条件下游泳行为,观测发现试验鱼在突进游速测量过程中的游泳状态主要有:顺流而下、逆流静止、逆流冲刺、逆流后退,这4种游泳状态相互穿插,并且在不同的流速阶段各类游泳状态所占的时间比例各不相同。根据其游泳行为的变化,可以将整个试验过程分成4个阶段:
第1阶段:突进游速测定的开始阶段,鱼体迅速调整自己的身体机能以适应快速和大幅度的流速变化。这一阶段鱼类逆流静止所占时间比例约为30%~57%左右;逆流向前时间约在21%~42%左右;逆流后退时间占20%~30%,3种状态分布相对均匀。顺流而下所占比例很小,仅为1%~3%。这一阶段试验鱼逆流前进和逆流后退的距离均较小,不超过1BL,未出现快速冲刺现象,总体而言水流的胁迫作用不明显,该阶段试验鱼的最大游速约为突进游速的52%~60%。
第2阶段:当流速增加到超过突进游速的60%时,试验鱼的摆尾频率增加明显,鱼体肌肉处于相对紧张状态。该阶段试验鱼游泳行为主要以逆流静止状态为主,所占比例约为41%~85%,逆流向前时间约占8%~32%,逆流后退时间约占7.5%~28%,且逆流前进和逆流后退的距离较小,不超过1BL。该阶段水流对鱼类的游泳的阻碍作用开始显现,但未出现试验鱼快速冲刺现象,这一阶段试验鱼的最大游速约为突进游速的76%~79%。
第3阶段:当水体流速继续增大到一定阶段,试验鱼开始出现反复快速冲刺和后退现象,冲刺距离较前两个阶段明显增大, 冲刺距离约3~5BL,最大可从试验段的后部连续冲刺到试验段的前部(最大冲刺距离接近70 cm),反复2到3次。该阶段鱼类的最大游速约为试验鱼突进游速的90%~96%。这一阶段试验鱼主要以逆流冲刺和逆流后退为主,所占时间比例分别为:24%~52%和20% ~47%。逆流静止所占比列减小到9%~46%。水流对试验鱼的胁迫作用已经非常明显。
第4阶段:当试验流速继续增加,试验鱼不断后退,其尾部开始触到下游拦网,鱼类反复逆流向前和后退,但距离较小,一般不超过1BL,试验鱼已经接近力竭状态。在该阶段,逆流静止、逆流向前和逆流后退所占比例相当分别为:23%~43%,24%~40%,20%~53%,试验鱼已经无力冲刺。
将试验用鱼按体长分为以下4类,“1”代表体长6.00~8.50 cm鱼、“2”代表长8.71~11.01 cm的鱼、“3”代表长11.30~12.30 cm的鱼、“4”代表12.70~15.00 cm的鱼。4种体长的草鱼幼鱼在上述4个阶段的最大游速如表2所示,各种游泳状态所占时间比例如图6所示。在这4个游泳阶段,顺流而下只出现在了第1阶段,且时间很短,其余3个阶段均只出现了逆流游泳的状态。随着流速的增加,逆流静止状态呈现先增加后减小的趋势,逆流前进和逆流后退则随着流速的增加有增加的趋势。仅在第三个阶段试验鱼开始出现反复逆流长距离冲刺现象,进入第四阶段试验鱼已经无法冲刺,各逆流游泳状态比例相当,接近疲劳状态。每个阶段的最大游速依次约为鱼类突进流速的52%~60%、76%~79%、90%~96%和100%。
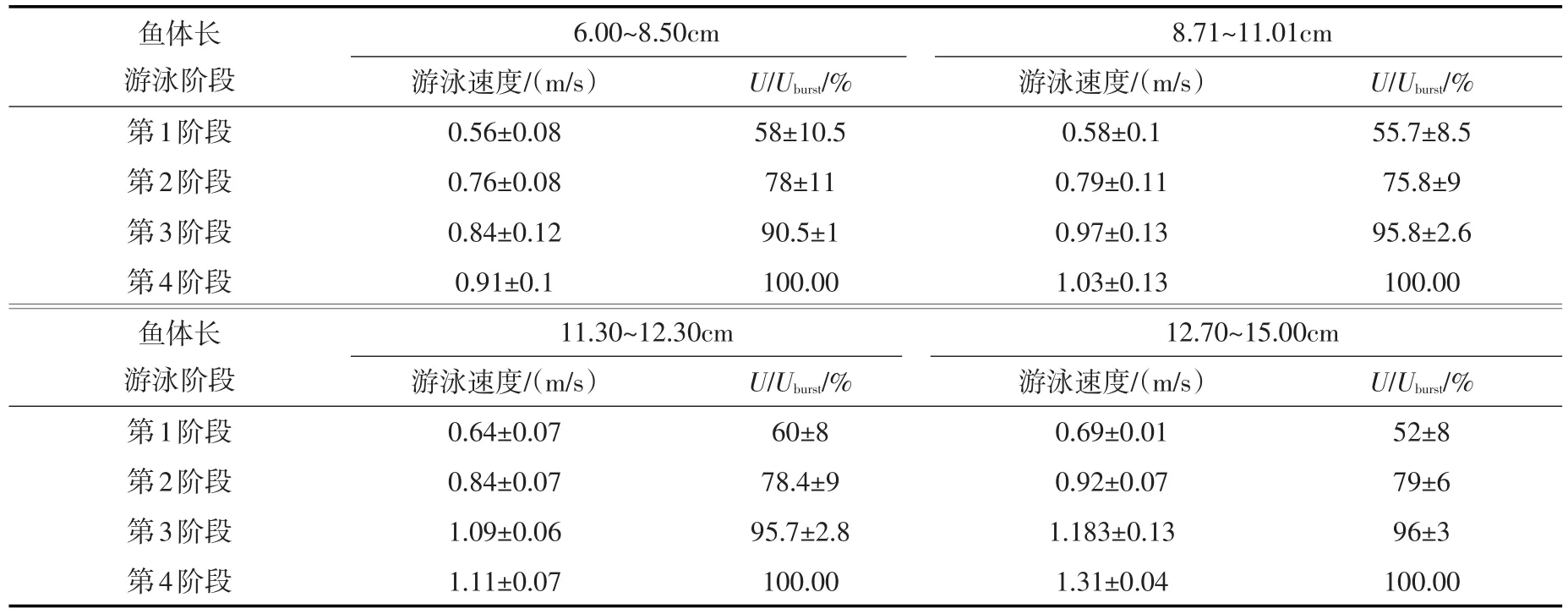
表2 草鱼幼鱼各游泳阶段的最大流速
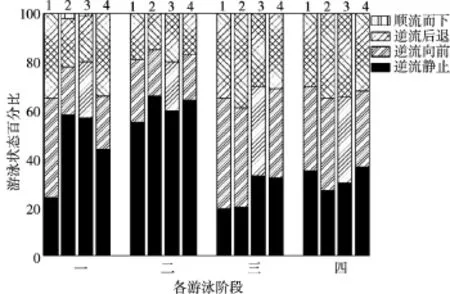
图6 突进游速测定过程中草鱼幼鱼游泳行为
由以上分析可知,随着流速的增加,水流对鱼类的胁迫作用开始显现并逐渐明显,最终达到鱼类游泳能力极限。从上述4个游泳阶段的分析来看,从第2阶段开始,水流对试验鱼的胁迫作用开始出现,进入第3个阶段试验鱼开始出现反复冲刺现象,水流胁迫效应变得十分明显,因此以草鱼为过鱼对象的鱼道池室主流最大流速不宜超过第1阶段的最大流速(突进游速的52%~60%),该条件下鱼类能够较为容易的沿主流上溯。房敏等[29]研究了体长 8~9.7 cm的草鱼幼鱼的临界游泳速度,通过分析试验鱼的新陈代谢,认为以草鱼为过鱼对象的鱼道内流速不应大于0.8Ucrit,即45~55.6 cm/s,而本次试验获得相应体长范围内的草鱼幼鱼第1阶段的最大流速范围为50.8~57.1 cm/s,二者所得池室临界流速范围较为接近,可以相互印证。
而对于竖缝、孔口等鱼道的高流速区,其最佳流速应该不高于第2阶段的最大流速(突进游速的76%~79%),此条件下草鱼幼鱼能够较为容易的穿越高流速区的水流障碍;如果竖缝、孔口处的流速难以降低,可以适度增加,但绝不能超过第3阶段的最大流速(突进游速的90%~96%),否则竖缝、孔口将成为草鱼幼鱼上溯的水流障碍。
4 结论
本文在25±0.5℃水温条件下,针对体长在5.0~16.0 cm范围内的草鱼幼鱼,利用鱼类游泳行为测试水槽开展了鱼类感应流速和突进游泳速度的试验研究,并对比分析了草鱼幼鱼临界游速与突进游速之间的关系、草鱼幼鱼突进游泳行为对于流速变化的响应,具体结论如下:(1)通过感应流速试验,得到了体长10.31~16.01 cm,体重范围12.16~39.08 g的草鱼幼鱼的感应流速范围为10.72~15.55 cm/s,其平均感应流速为12.84 cm/s。在进行鱼道设计时,要确保鱼道主流流速大于感应流速,使鱼类上溯时能够感知前进方向。(2)绝对突进游泳速度随着鱼体长的增加而增加,呈线性正相关,草鱼幼鱼突进游速与体长的关系可表示为:Uburst=6.637BL+37.572;相对突进游泳速度随鱼体长的增加而线性减小,其关系式为:U′burst=-0.430BL+14.908。(3)通过将试验得到的草鱼幼鱼突进游速公式与之前龚丽等[6]得出的5~15 cm的草鱼幼鱼临界游速公式进行对比分析,得出草鱼幼鱼临界游速与突进游速的关系式Uburst=1.676Ucrit-42.024。鱼的体长越长,突进游速超出临界游速就越多。(4)在突进游速试验过程中,试验鱼游泳状态主要有:顺流而下、逆流静止、逆流冲刺、逆流后退4种,这4种游泳状态相互穿插。根据草鱼幼鱼游泳行为的变化,可将整个流速递增的试验过程分成4个阶段:仅第1阶段试验鱼出现了顺流而下的游泳状态,草鱼幼鱼可以在实验区自由游泳,受水流的胁迫作用不明显,该阶段试验鱼的最大游速约为突进游速的52%~60%,以草鱼为过鱼对象的鱼道池室主流最大流速不宜超过第1阶段的最大流速。第2—4阶段试验鱼始终保持逆流游泳状态。第2阶段试验鱼摆尾频率增加,以原地逆流游泳为主,水流胁迫作用开始显现;第3阶段试验鱼出现反复快速冲刺随后逆流后退的现象,水流胁迫作用十分明显;第4阶段试验鱼已经无法长距离冲刺,接近力竭状态。因此对于竖缝、孔口等鱼道的高流速区,其最佳流速应该不高于第2阶段的最大流速(突进游速的76%~79%),绝不可超过第3阶段的最大流速(突进游速的90%~96%)。
[1]农业大词典委员会.农业大词典[M].北京:中国农业出版社,1998.
[2]长江三峡工程生态与环境监测公报[R].中国环境监测总站,中华人民共和国环境保护部,2013.
[3]鲜雪梅,曹振东,付世建,等.4种幼鱼临界游泳速度和运动耐受时间的比较[J].重庆师范大学学报:自然科学版,2010,27(4):16-20.
[4]房敏,蔡露,等.温度对鲢幼鱼游泳能力及耗氧率的影响[J].水生态学杂志,2013,34(3):49-53.
[5]蔡露,涂志英,袁喜,等.鳙幼鱼游泳能力和游泳行为的研究与评价[J].长江流域资源与环境,2012,21(Z2):89-95.
[6]龚丽,吴一红,白音包力皋,等.草鱼幼鱼游泳能力及游泳行为试验研究[J].中国水利水电科学研究院学报,2015,13(3):211-216.
[7]熊锋,王从锋,刘德富,等.松花江流域青鱼、草鱼、鲢及鳙突进游速比较研究[J].生态科学,2014,33(2):339-343.
[8]黎采微,路波,陈廷,等.青鱼逃逸过程中的疾冲-滑行游泳行为[J].动物学杂志,2014,49(6):860-866.
[9]牛宋芳,路波,罗佳,等.鲢快速逃逸游泳行为研究[J].水生生物学报,2015(2):394-398.
[10]路波,刘伟,梁圆圆,等.草鱼快速启动过程的加速—滑行游泳行为[J].水产学报,2014,38(6):829-834.
[11]赵希坤,韩桢锷.鱼类克服流速能力的试验[J].水产学报,1980,4(1):31-37.
[12]胡茂林.鄱阳湖湖口水位、水环境特征分析及其对鱼类群落与洄游的影响[D].南昌:南昌大学,2009.
[13]JAIN K E,FARRELL A P.Influence of seasonal temperature on the repeat swimming performance of rainbow trout Oncorhynchus mykiss[J].Journal of Experimental Biology,2003,206(20):3569-3579.
[14]殷名称.鱼类生态学[M].北京:中国农业出版社,1995.
[15]黄真理,常剑波.鱼类体长与体重关系中的分形特征[J].水生生物学报,1999,23(4):330-336.
[16]王武.鱼类增养殖学[M].北京:中国农业出版社,2000.
[17]徐革锋,尹家胜,韩英,等.不同适应条件对细鳞鲑幼鱼游泳能力的影响[J].生态学报,2015,35(6):1938-1946.
[18]白艳勤,路波,罗佳,等.草鱼、鲢和瓦氏黄颡鱼幼鱼感应流速的比较[J].生态学杂志,2013,32(8):2085-2089.
[19]BLANK M D,CAHOON J,MCMAHON T.Using three dimensional hydrodynamic modeling and fish swimming energetics to assess culverts as potential physical barriers to upstream fish movement[C]//Transportation Research Board 88th Annual Meeting.2009.
[20]BLAKE R W.Functional design and burst-and-coast swimming in fishes[J].Canadian Journal of Zoology,1983,61(11):2491-2494.
[21]袁喜,涂志英,韩京成,等.流速对鲫游泳行为和能量消耗影响的研究[J].水生态学杂志,2011,32(4):103-109.
[22]田凯,曹振东,付世建.速度增量及持续时间对瓦氏黄颡鱼幼鱼临界游泳速度的影响[J].生态学杂志,2010,29(3):534-538.
[23]LYON J P,RYAN T J,SCROGGIE M P.Effects of temperature on the fast-start swimming performance of an Australian freshwater fish[J].Ecology of Freshwater Fish,2008,17(1):184-188.
[24]KIEFFER J D.Limits to exhaustive exercise in fish[J].Comparative Biochemistry&Physiology Part A Molecular&Integrative Physiology,2000,126(2):161-179.
[25]蔡露,房敏,涂志英,等.与鱼类洄游相关的鱼类游泳特性研究进展[J].武汉大学学报:理学版,2013,59(4):363-368.
[26]RANDALL D,BRAUNER C.Effects of environmental factors on exercise in fish[J].Journal of Experimental Biology,1991,160(2):113-126.
[27]房敏,蔡露,高勇,等.温度对鲢幼鱼游泳能力及耗氧率的影响[J].水生态学杂志,2013,34(3):49-53.
[28]HAMMER C.Fatigue and exercise tests with fish[J].Comparative Biochemistry&Physiology Part A Physiology,1995,112(1):1-20.
[29]房敏,蔡露,高勇,等.运动消耗对草鱼幼鱼游泳能力的影响[J].长江流域资源与环境,2014,23(6):816-820.
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
基层中医药(2021年11期)2021-03-26
小学生作文(低年级适用)(2019年4期)2019-04-29
散文诗(2017年18期)2018-01-31
猪业科学(2016年1期)2016-03-30
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
应用海洋学学报(2014年1期)2014-11-22
天然产物研究与开发(2014年8期)2014-04-27
