
我国观赏植物组织培养育种研究进展
2018-01-10 09:27:28傅聿峰
河北林业科技 2017年4期
傅聿峰
(河北省塞罕坝机械林场,河北 围场 068450)
我国观赏植物组织培养育种研究进展
傅聿峰
(河北省塞罕坝机械林场,河北 围场 068450)
现代组织培养育种技术主要包括单倍体育种、多倍体育种、原生质体与体细胞杂交育种和胚挽救技术育种等4种。在对过去10a中观赏植物组织培养育种的研究情况进行总结的基础上,对其发展进度、应用情况进行了简介,列出了试验成功的主要观赏植物种类,并对观赏植物组织培养育种的材料进行了综述和分析,指出当前阶段存在的问题和不足,以期为今后观赏植物组织培养育种提供参考。
观赏植物;组织培养;育种;研究进展
观赏植物组织培养,就是通过无菌操作分离观赏植物体的一部分,如花、茎尖、茎段、叶、幼胚等,并配合一定的营养、生长调节物质,通过严格控制温度、光照等因素,使其迅速生长,能够缩短其生长周期,获得大量遗传性状一致的幼苗,又称植物克隆育苗技术(Plant Gene Cloning)[1]。观赏植物组织培养对新品种的选育,或者利用现代种质资源及野生种质资源进行常规的杂交、倍性、辐射及基因工程育种具有重要意义,得到了广泛应用。兰花(Cymbidium ssp.)、菊花(Chrysanthemum)、唐菖蒲(Gladiolus gandavensis Vaniot Houtt)、非洲紫罗兰(Saintpaulia ionantha Wendl)、 水 仙(Narcissus tazettaL.var.chinensisRoem.)、 百 合(Lilium brownii var.viridulum Baker)、矮牵牛(Petunia hybrid Vilm)、鹤望兰(Strelitzia reginae Aiton)等的主要繁殖方式就是组织培养。
现在除了常规方法外,在现代花卉新品种的选育中,植物组织培养技术参与辅助培育或直接培育的新品种所占比例超过80%,主要是通过倍性育种、细胞育种、原生质体育种、胚挽救技术等来实现的。本文对10a来这4个方面的研究成果进行讨论。
1 单倍体育种
单倍体育种是利用植物组织培养技术诱导产生单倍体植株,再通过某种手段使染色体组加倍(如用秋水仙素处理),从而使植物恢复正常染色体数,具有高速、高效、基因型一次纯合等优点,现已作为一种崭新的育种手段开始应用。单倍体获得途径包括花粉培养、花药培养、胚培养、子房培养等。花药、花粉培养是在离体情况下培养成单倍体植株最常用的技术,是得到大量单倍体最有效的方法,可缩短育种年限,提高选择效率,在很多观赏植物上已经获得成功植株(表1)[2-22]。诱导花粉植株频率低或不能诱导出花粉植株的种类,可通过培养未授精子房诱导单倍体植株。子房的人工培养,可有效解决受精后的发育障碍,使大量远缘杂交胚继续发育成正常种子以加快育种进程,缩短育种周期,同时可获得大量的远缘杂交后代,创造新种质。
花药是植物雄性生殖器官,由2种不同倍性的细胞组成,药壁和药隔组织为二倍体细胞,雄性生殖细胞(花粉粒)为单倍体细胞。花药和花粉培养是改变花粉原来的发育途径,诱导花粉分裂增殖,形成愈伤组织,经过再分化,发育成单倍体植株[23]。
1964年Guha和Maheshwari首次从曼佗罗花药培养中诱导出单倍体。我国的花药培养育种研究始于1972年,到现在为止,至少20个以上的花粉植株是我国学者首先完成的,居于世界领先地位。近10年来,我国观赏植物的花粉培养主要是对花粉最佳萌发条件进行研究,如茉莉(Jasminum sambac Aiton))、梅花(Prunus mume)、桔梗[Platycodon randiflorum(Jacq.)A.DC]、金花茶(Camellia nitidissima)、大叶相思花(Acacia auriculiformis)、宫粉羊蹄甲(Bauhinia variegate L.)、唐菖蒲、桃(Prunus persica(L.)Batsch)、无距虾脊兰(Calanthe tsoongiana T.Tang et F.T.Wang)、有斑百合(Lilium concolor var.pulchellum)[24-33]等。
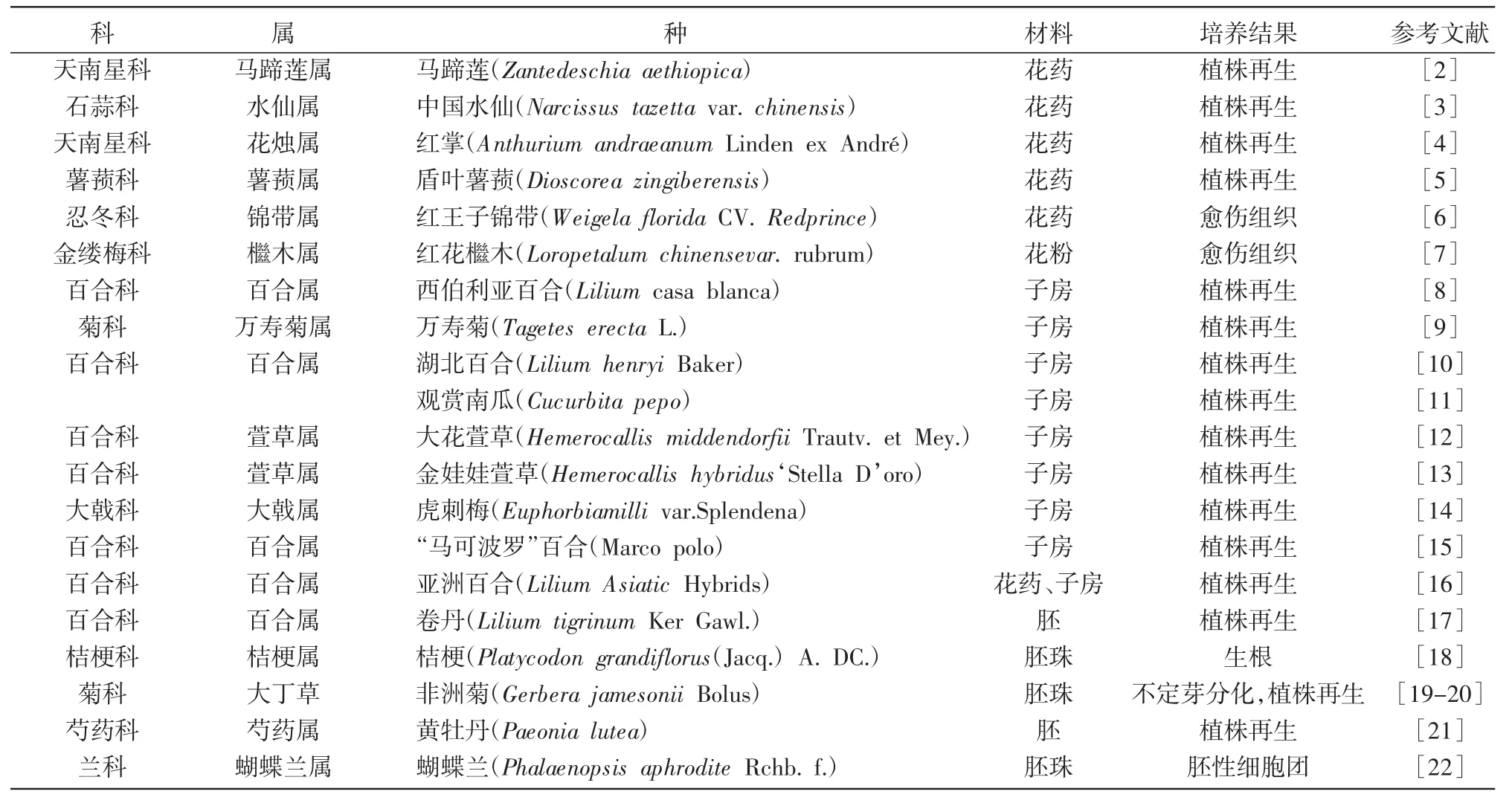
表1 单倍体离体培养的主要观赏植物科、属、种类(2006~2016年)
2 多倍体育种
多倍体在自然界广泛存在,具有植株巨型化、抗逆性增强、营养成分含量增加等特点,被广泛应用于育种研究中。被子植物的胚乳是双受精的产物,属于三倍体,通过被子植物的胚乳离体培养可以获得三倍体植株,三倍体品种具有产量高、观赏性强及不结实等优势而备受育种工作者的关注。而将三倍体植株通过染色体加倍,可以获得六倍体植株,以解决三倍体不结实的问题,并用于培育多倍体新品种。多倍体生物一旦形成,它和原来的物种就发生生殖隔离,因而成了新种。
与组织培养相结合的多倍体诱导技术已经成为当今多倍体诱导的主要手段,能够使人工诱导多倍体的成功率大大提高,获得大量材料,提高诱导成功率,及时分离嵌合体。在组培上常用现代生物技术诱导多倍体,材料一般是种子、幼苗、生长点、茎尖、愈伤组织、胚状体、悬浮细胞系、小孢子、原生质体或单细胞等,诱变材料选取植株或组织分裂最旺盛的时期和部位。诱变剂种类一般选用秋水仙素或除草剂,或添加辅助剂,包括二甲基亚砜(DMSO)、甘油等。诱导方法主要有3种,即组织培养结合化学诱变、胚乳培养、原生质体融合。近几年来多倍体诱变结果见表2。利用秋水仙素处理组织培养过程中的丛生芽、原球茎、体细胞胚、种子萌发幼苗等是诱导观赏植物多倍体的有效方法,目前已在百合、大岩桐等多种花卉上取得成功。胚乳是天然的三倍体组织,因此,胚乳培养可获得三倍体植株,在盾叶薯蓣、乌桕中成功获得植株[34-36],花卉中应用较少,再生植株多为非整倍体、混倍体,不稳定。
虽然经过处理后能够得到大量多倍体,但是同时也得到大量的非整倍体和嵌合体,需要做后期鉴定。

表2 多倍体离体诱导的主要观赏植物科、属、种类(2006~2016年)
3 原生质体育种与体细胞杂交育种
植物原生质体(protoplast)是指除去细胞壁的,由质膜包裹的裸露“细胞”,因为没有细胞壁的束缚,无论亲缘关系如何,都具有彼此融合的能力,实现远缘的植物细胞质融合,体细胞杂交,是在离体条件下将同一物种或不同物种的原生质体融合,培养并获得再生植株。
融合方式常用的有对称融合和不对称融合两种。植物的原生质体培养及其体细胞杂交是创造新物种、克服不亲和性的有效手段,可以利用其进行遗传转化,可以实现抗病性、抗逆性、细胞质雄性不育和优良品质特性的有效转移[51]。叶片、下胚轴、胚性愈伤组织、花粉等都是原生质体的良好来源,再生途径通常有3条:①经细胞团发育形成愈伤组织,再分化成芽长成植株。②直接形成胚状体,再发育成植株。③先形成愈伤组织,再由愈伤组织分化形成胚状体,最后发育成植株。
Carlson等在1972年首次成功运用体细胞融合技术获得烟草种间的体细胞杂种再生植株,自此越来越多的物种取得了进展,现在很多观赏植物悬浮细胞培养的体系都已经建立,获得多个种间、属间和科间的体细胞杂种植株。研究成果见表3。
但是,利用原生质体育种也有一定的不足。首先,双亲原生质体融合时,有利基因连同不利基因可能一并融合到了杂种细胞中,若要去除携带的不利基因,则需通过传统育种中的连续多次回交才有望实现,该操作耗费精力颇多,且年限较长;其次,当双亲的2个核不融合时,引发形成嵌合体,使得优良性状不能如期整合到杂种细胞中并遗传到下一代[79]。
4 胚挽救技术
植物远缘杂交受精后,由于生理原因造成植物难以播种成苗或胚在发育早期阶段败育、退化,可在胚未完全败育前进行杂种胚的离体培养,用培养基来提供胚成长过程中需要的各类营养,从而使胚能够正常生长并且发展成幼苗,这种方法称之为胚挽救技术。此技术可以克服生殖隔离障碍,成功获得远缘杂种植株,大大提高了成功率。胚挽救方式主要有子房切片培养、胚培养、胚珠培养等3种方式,杂种胚培养成功的关键,一是筛选适宜的培养基,二是选择培养的适宜时期。现在主要研究不同胚救方式、不同胚拯救时间、不同杂交组合对胚挽救结果的影响,在百合上发现通过胚培养方式进行胚拯救获得的萌发率、愈伤形成率和成苗率均要高于胚珠培养[80],母本在石蒜属植物育种中的选择很关键,杂交自然结实率越高的组合胚拯救成功率越高[81]。
目前,已通过胚挽救的方法获得了很多种间、甚至属间的杂种植株,扩大了杂种优势的利用范围。在观赏植物中,研究最多的是百合[82-84],以及鸢尾属[85]、牡丹[86]、石蒜[87]等。研究情况见表 4。

表3 细胞、原生质体培养的主要观赏植物科、属、种类(2006~2016年)

表4 细胞、原生质体培养的主要观赏植物科、属、种类(2006~2016年)
5 小结
目前,组织培养育种技术是今后发展的方向,培养体系越来越成熟,培育出了很多新品种,但是还存在诸多问题:(1)此技术必须建立在对某物种组培技术比较成熟的基础上且培养体系探索过程很长。(2)在进行多倍体诱导时获得不定结果较多,还需做倍性鉴定。(3)没有普遍性指导意义的结果。所以,组织培养育种技术有待于进一步深入研究。
[1]Niu G,Kozai T,Kitaya Y.Simulation of the timeco of
concentmtmfon in the Culture vesseland net photosynthetic rate ofCymbidium Tissue Culture technology,flower,modification of characters,plantlets[J].Transactions of the ASAE,1996,39(4):1567-1573.
[2]张喜艳.马蹄莲花药培养技术研究[D].昆明:云南农业大学,2009:35.
[3]张清国,黄敏玲,叶秀仙.中国水仙花药培养及植株再生体系建立[J].分子植物育种,2010,8(2):399-404.
[4]杜宝贵,黄丽娟,张志胜.红掌花药培养[J].生物技术通报,2009(增刊):189-195.
[5]闫立霞,胡春根,姚家玲.盾叶薯蓣花药培养及单倍体植株的获得[J].云南植物研究,2007,29(1):33-37.
[6]王丹.红王子锦带花药离体培养研究[D].哈尔滨:东北林业大学,2012:31.
[7]李炎林,于晓英,熊兴耀,等.红花檵木花粉单倍体愈伤组织的诱导与培养[J].湖南农业大学学报(自然科学版),2011,37(6):632-636.
[8]申玉华,段永平,唐立红,等.西伯利亚百合子房的组织培养及离体快繁研究[J].北方园艺,2008(11):151-152.
[9]张华丽,赵剑颖,张睿鹂.万寿菊杂交亲本的离体培养[J].西北植物学报,2011,31(12):2545-2550.
[10]潘娟.湖北百合组织培养与快繁技术研究[D].重庆:西南大学,2009:30.
[11]孙守如.观赏南瓜未受精子房、胚珠离体培养及其机理研究[D].郑州:河南农业大学,2011:66.
[12]杨丽莉,张晓,杨睿,等.大花萱草‘莎蔓’的组织培养技术研究[J].北方园艺,2012(9):134-137.
[13]杨丽莉,王德平,张晓,等.以子房为外植体的‘金娃娃’萱草组织培养技术的研究[J].中国农学通报,2012,28(25):184-190.
[14]孔德平,王增池.虎刺梅的组织培养及快速繁殖初报[J].安徽农业科学,2008,36(15):6208.
[15]杨美纯,卢赛清,陈燕霞,等.东方百合“马可波罗”花器官的组织培养[J].广西农业生物科学,2008,27(2):134-137.
[16]姚绍嫦,杨美纯,凌征柱.亚洲百合花器官组织培养再生植株研究[J].安徽农业科学,2010,38(7):3459-3460.
[17]雷家军,庞兰,林翼飞,等.卷丹百合种间杂种胚培养研究[J].沈阳农业大学学报,2009,40(1):84-87.
[18]徐芳芳.桔梗胚囊的发育特点与胚珠组织培养研究[D].延边:延边大学,2015:35.
[19]单芹丽,李绅崇,王继华,等.基因型和预处理对非洲菊胚珠离体培养的影响[J].江西农业学报,2015,27(7):44-47.
[20]王丽花,瞿素萍,吴学尉,等.非洲菊未受精胚珠离体培养影响因素研究[J].西南农业学报,2013,26(4):1639-1644.
[21]律春燕.黄牡丹野生种与牡丹、芍药栽培品种远缘杂交研究[D].北京:北京林业大学,2010:46.
[22]伍成厚,潘一山,罗开梅,等.蝴蝶兰未受精胚珠离体培养的研究[D].园艺学报,2006,33(4):891-894.
[23]沈海龙.植物组织培养[M].北京:中国林业出版社,2005:125-128,200-203.
[24]邓衍明,叶晓青,梁丽建,等.茉莉花粉离体培养萌发及花粉管生长特性研究[J]. 华北农学报,2014,29(5):107-113.
[25]赵宏波,房伟民,陈发棣.梅花花粉离体萌发和花粉管生长研究[J].广西植物,2007,27(3):393-396.
[26]刘自刚,呼天明,杨亚丽,等.桔梗花粉萌发与花粉管生长研究[J].植物研究,2011,31(3):271-276.
[27]张佩霞,于波,陈金峰,等.金花茶花粉离体萌发及低温处理探究[J].西部林业科学,2016,45(2):94-97.
[28]詹妮,黄烈健.大叶相思花粉离体萌发适宜条件及活力检测方法[J].林业科学,2016,52(2):62-73.
[29]贺庆梅,陈岩岩,李启虔,等.宫粉羊蹄甲花粉离体萌发培养基的筛选[J]. 南方农业学报,2015,6(2):2173-2179.
[30]黄蓓丽,耿兴敏,胡凤荣,等.唐菖蒲花粉离体萌发的液体培养基研究[J]. 安徽农业大学学报,2009,36(4):593-597.
[31]杜纪红,叶正文,苏明申,等.桃花粉离体萌发和花粉管生长特性研究[J].西北植物学报,2011,31(1):0064-0071.
[32]钱鑫,刘芬,牛晓玲,等.无距虾脊兰花粉离体萌发及储藏条件的研究[J].西北植物学报,2014,34(2):0314-0348.
[33]郭思佳,朱玉菲,刘冬云,等.有斑百合花粉培养条件的研究[J].河北农业大学学报,2014,37(3):24-28.
[34]李明军,郭婧,李翔,等.盾叶薯蓣胚乳再生体系的建立及其染色体倍性鉴定[J].植物学报,2012,47(6):654-660.
[35]郭永兵,江道菊,李特林.盾叶薯蓣胚乳的组织培养[J].北方园艺,2007(10):175-177.
[36]田良涛.乌桕胚乳三倍体植株再生与转基因研究[D].武汉:湖北大学,2011:23
[37]赵波.黄璧组织培养及转基因体系的建立[D].哈尔滨:东北林业大学,2007:19.
[38]熊海浪,于振艳,张宗申,等.影响濒危药用植物滇重楼愈伤组织发生的因素[J]. 时珍国医国药,2012,23(6):1372-1274.
[39]林美珍,巫庆珍,郑松,等.巴戟天的组织培养和多倍体诱导[J].中国医药杂志,2011,36(17):2325-2328.
[40]余道平,李策宏.迎阳报春四倍体诱导及鉴定[J].核农学报,2014,28(6):0961-0966.
[41]于娜丽,王良桂.百合多倍体诱导及鉴定[A/C].中国风景园林学会2011年会论文集,2011:1015-1021.
[42]曹方莉.灰毡毛忍冬金银花愈伤组织及多倍体的诱导[D].重庆:中南林业科技大学,2008:42.
[43]葛蓓学.细叶百合组织培养及多倍体诱导研究[D].哈尔滨:东北林业大学,2010:34.
[44]梁微.大叶秦艽的组织培养及离体诱导多倍体的研究[D].西安:西北大学,2007:60.
[45]谈永霞.百里香组织培养及秋水仙素诱导多倍体的初步研究[D].兰州:兰州理工大学,2011:39.
[46]苏小玲,张双双,许雯婷,等.阔叶风铃草多倍体诱导与鉴定[J].安徽农业科学,2010,38(21):11627-11629.
[47]李政,黄静洁,李凌.秋水仙碱诱变绿玉树多倍体研究[J].西南大学学报(自然科学版),2007,29(2):106-110.
[48]李志虹,高英,倪苏,等.秋水仙素离体诱导对非洲紫罗兰形态的影响[J].北方园艺,2010(5):153-155.
[49]陈艾,杨利平,谭艳.秋水仙素诱变离体卷丹多倍体的研究[J].植物遗传资源学报,2014,15(6):1385-1389.
[50]王瑞,张亚楠,王雅英,等.中国水仙六倍体的诱导和染色体数目的变异(简报)[J].分子细胞生物学报,2007(40):263-270.
[51]聂琼,杨川龙,徐如宏,等.聚乙二醇法诱导烟草原生质体融合的条件优化[J].山地农业生物学报,2012,31(4):288-212.
[52]芦婕.盾叶薯蓣三倍体细胞悬浮培养及植株再生[D].郑州:河南师范大学,2014:37.
[53]吴丹丹.黄秋葵组织培养及胚性细胞悬浮体系的建立[D].福州:福建农林大学,2008:22.
[54]单国燕.柱花草组织培养和细胞悬浮培养的研究[D].长沙:湖南农业大学,2010:44.
[55]潘兵兵.5个现代月季品种愈伤组织再生体系的建立与体细胞培养的研究[D].哈尔滨:东北农业大学,2011:53-55.
[56]王娟,李玉珠,师尚礼.苜蓿愈伤组织原生质体游离与培养[J].草地学报,2010,18(2):25-262.
[57]赵小强,马晖玲,林栋,等.草地早熟禾新格莱德胚性愈伤组织原生质体培养及植株再生的研究[J].草业学报,2010,19(2):55-60.
[58]冯博.神农香菊(Dendranthema indicum var.Aromaticu)胚性细胞悬浮培养及原生质体融合的初步研究[D].哈尔滨:东北林业大学,2012:32.
[59]肖望,涂红艳,邓崇会.红姜花胚性细胞悬浮体系的建立和原生质体培养[J].北方园艺,2015(5):104-108.
[60]付广俊.山茱萸细胞悬浮培养及熊果酸含量分析[D].合肥:安徽农业大学,2012:26.
[61]朱新洲.白鲜组织培养技术及其内生真菌分离和作用的研究[D].长沙:湖南农业大学,2008:46.
[62]徐世千,李晓东,张建国.百里香愈伤组织的诱导及细胞悬浮培养体系的建立[J].西北农业学报,2011,20(10):112-119.
[63]毛艳萍,苏智先,胡进耀,等.濒危植物珙桐愈伤组织的诱导及悬浮细胞培养初探[J].武汉植物学研究,2010,28(4):510-515.
[64]李玉平.大花金挖耳细胞培养活性物质的研究[D].杨凌:西北农林科技大学,2008:191.
[65]赵荣荣.大叶秦艽组织与细胞培养及NO对次生代谢物龙胆苦苷形成的作用[D].西安:西北大学,2012:24.
[66]陈慧丽.厚朴细胞培养与超低温保存的研究[D].福州:福建农林大学,2012:41.
[67]赵俊,吴虹丽,马林.鸡屎藤愈伤组织诱导及细胞悬浮培养研究[J].南方农业,2013(5):53-56.
[68]董春艳.神农香菊(Dendranthema indicum var.Aromaticu)组织培养及细胞悬浮培养体系的建立[D].哈尔滨:东北林业大学,2011:36.
[69]张彦妮,陈素波.长叶点地梅的细胞悬浮培养条件[J].草业科学,2014,31(8):1481-1486.
[70]刘剑锋,程云清,陈智文.高山红景天叶肉原生质体分离培养与植株再生[J].中草药,2009,40(7):1127-1131.
[71]张改娜,贾敬芬.草木樨状黄芪A4转化系原生质体培养研究[J].河南农业科学,2009(8):46-50.
[72]杨仙,马丹丹,蒋福升,等.滴水珠原生质体的分离纯化与植株再生[J].2014,39(21):4211-4515.
[73]陈颖,梁建丽,陈晓丽,等.多花蔷薇胚性细胞悬浮系原生质体分离及再生植株[J]. 园艺学报,2008,35(10):1491-1496.
[74]李名扬,陈薇.菊花花瓣愈伤组织原生质体培养再生植株[J].农业生物技术学报,1996,4(3):243-248.
[75]范小峰,李东波,刘灵霞,等.南蛇藤原生质体培养及植株再生[J].植物研究,2011,31(3):300-305.
[76]郝建国,贾敬芬.沙葱叶基愈伤组织原生质体再生体系的建立[J].基因组学与应用生物学,2009,28(5):998-1001.
[77]王鹏凯,吕伟光,胡雪怡,等.北美鹅掌楸原生质体的分离与培养[J].西北植物学报,2013,33(22):0254-0260.
[78]秦晓杰,段华金,朱永平,等.东方百合‘Sorbonne’原生质体培养初步研究[J].分子植物育种,2013,11(5):600-604.
[79]戴雪梅,黄天带,孙爱花,等.植物原生质体融合研究进展及其在育种中的应用[J].热带作物学报,2012,33(8):1516-1521.
[80]郑思乡,刘飞虎,龚建华,等.切花百合组间杂交的初步研究[A/C].第四届全国花卉资源、育种、栽培及应用技术交流会论文汇编,2016:145-152.
[81]金梅.石蒜属植物种间杂交及后代培养的研究[D].南昌:江西农业大学,2013.
[82]夏晶,吴学尉,崔光芬,等.西伯利亚与黄天霸正反交实验及杂种胚离体培养[J].西南大学学报(自然科学版),2010,32(6):64-67.
[83]崔光芬,王继华,贾文杰,等.OT百合新品种‘火凤凰’,和‘金龙’[J].园艺学报,2013,40(10):2097-2098.
[84]张朝君,郑思乡,许超,等.OT型百合与O型百合远缘杂交试验[J].贵州农业科学,2010,38(4):17-18.
[85]韩耀祖.唐菖蒲与射干属间杂交初步研究[D].哈尔滨:东北林业大学,2013.
[86]郭子霞.牡丹花粉贮藏及杂交幼胚的挽救[D].郑州:河南农业大学,2012.
[87]郑思乡,吴学蔚,许超,等.淡黄花百合与东方百合‘如意’远缘杂交的研究[J].西部林业科学,2009,38(4):5-8.
Research Progress on Tissue Culture and Breeding of Ornamental Plants in China
FU Yu-feng
(The Forest Farm of Saihanba in Hebei,Weichang,Hebei 068450,China)
The modern organization culture breeding includes haploid breeding and polyploid breeding and protoplast and somatic hybridization and embryo rescue technology breeding.In the past 10 years,the development progress and its application of modern organization culture breeding were introduced,and the main ornamental plants types of successful test were listed,at the same time,the breeding material of ornamental plant tissue culture were summarized and analyzed.Finally this paper pointed out the existing problems and shortcomings in order to provide reference for breeding in future.
Ornamental plants;Tissue culture;Breeding;Research progress
S722.8
A
1002-3356(2017)04-0042-06
2017-11-22
傅聿峰(1968-),男,林业高级工程师,从事林业产业管理工作。
猜你喜欢
今日农业(2020年14期)2020-12-14 19:47:34
诊断学(理论与实践)(2020年6期)2020-12-09 04:07:52
山西农业科学(2020年8期)2020-08-13 07:44:06
河南农业科学(2018年5期)2018-01-19 08:25:55
食用菌(2017年5期)2017-10-19 03:02:28
临床与实验病理学杂志(2017年3期)2017-03-07 09:33:00
大连工业大学学报(2015年4期)2015-12-11 04:06:50
中国学术期刊文摘(2015年8期)2015-10-29 09:51:18
特产研究(2015年1期)2015-04-12 06:36:11
西北农林科技大学学报(自然科学版)(2015年5期)2015-02-21 16:17:12
