
葡萄果肉组织的能量水平和细胞壁代谢对其自溶软化的影响
2018-01-08 02:47周文化
食品科学 2018年1期
张 群,周文化*,谭 欢,刘 伟
葡萄果肉组织的能量水平和细胞壁代谢对其自溶软化的影响
张 群1,2,周文化1,*,谭 欢2,刘 伟2
为研究葡萄采后贮藏过程中自溶软化与能量水平及细胞壁代谢的关系,比较贮藏初期和末期果皮超微结构的变化。采后葡萄经钙联合涂膜和热处理后,低温((4.0±0.5) ℃)贮藏,定期测定果肉自溶指数、硬度、腐烂率、膜透性、能量物质(三磷酸腺苷(adenosine triphosphate,ATP)、二磷酸腺苷(adenosine diphosphate,ADP)、单磷酸腺苷(adenosine monophosphate,AMP))含量及能荷(energy charge,EC)、细胞壁降解酶活力和细胞壁成分的变化,扫描电子显微镜观察贮藏初期和末期不同处理果实内果皮的超微结构,以未处理组为对照。结果显示:随贮藏时间的延长,果实自溶指数、腐烂率和膜透性升高,硬度和能量物质下降,多聚半乳糖醛酸酶(polygalacturonase,PG)、果胶甲酯酶(pectinesterase,PE)和纤维素酶(cellulase,Cx)活力上升,β-半乳糖苷酶(β-galactosidase,β-Gal)活力在贮藏30 d内下降,之后急剧升高。细胞壁组分原果胶、纤维素和半纤维素含量下降,水溶性果胶含量前期上升,后期下降;葡萄内果皮的超微结构破坏,出现了大的孔洞。葡萄果实自溶指数与能量水平、细胞壁降解酶活力和细胞壁组分紧密相关,果实自溶指数与ATP、ADP含量极显著负相关(P<0.01),和AMP显著负相关(P<0.05),与EC无明显相关性(P>0.05);与PG、PE、Cx活力极显著正相关(P<0.01),与β-Gal活力无明显相关性(P>0.05);与原果胶、水溶性果胶、纤维素和半纤维素含量极显著负相关(P<0.01)。钙联合涂膜和热处理能够维持组织的高能量状态和致密的超微结构,抑制PG、PE和Cx活力的升高,延缓细胞壁降解,延缓自溶软化,其中涂膜显著优于热处理(P<0.05)。
葡萄;自溶软化;能量水平;超微结构;细胞壁代谢
葡萄(Vitis vinifera L.)属浆果类落叶藤本植物,营养丰富。但葡萄果实皮薄、汁液多、抵抗力差,且葡萄果实成熟于高温多雨夏季,采后在贮藏过程中易产生色泽、风味、营养物质等变化[1-3],其中最显著的特征变化是果肉质地的软化、流汁,这和龙眼采后“自溶”现象类似[4-5],是限制葡萄贮藏的关键因素之一。目前普遍认为,细胞壁结构改变和组分发生降解是导致果实质地软化的主要原因[4-10]。与葡萄果肉软化相关的细胞壁降解酶主要是多聚半乳糖醛酸酶(polygalacturonase,PG)、果胶甲酯酶(pectinesterase,PE)、纤维素酶(cellulase,Cx)和β-半乳糖苷酶(β-galactosidase,β-Gal)等[6]。研究发现应用钙处理[11-16]和涂膜[2,7,15]措施能抑制细胞壁降解酶(PE、PG、Cx等)活力和减缓细胞壁物质(原果胶、纤维素、半纤维素等)的降解,延缓果实的软化[2,7,11-16]。葡萄自溶软化过程中不仅外部形态发生了明显的变化,其内部的细胞超微结构也会发生变化,葡萄果肉组织超微结构的变化可以直观反映果肉品质劣变的过程[17-18]。
果实采后仍是活的有机体,果实组织能量代谢活动继续有序进行,细胞能量供应不足,加速机体的衰老和死亡[19-27]。近年来的研究发现,园艺作物采后衰老与能量合成下降而造成的细胞能量亏缺有关[19-25]。张群等[28]研究发现贮藏中葡萄果肉组织会出现能量亏损而影响其生理品质的劣变。Wu Ziming等[3]认为热处理能减缓葡萄软化的部分原因可能是热处理增加了碳水化合物代谢酶活性,提高了能量物质的供应,维持高的能量状态。但具体的软化与能量代谢的相关研究鲜见报道。
葡萄采后处理中常用的热处理[2-3,6]和涂膜[2,14-15]可有效延缓果实衰老。钙能维持和加强果实细胞壁和细胞膜的结构和功能,调节果实的呼吸代谢和生理代谢,维持细胞膜结构的完整性和细胞膜系统的区室化,防止胞内底物与酶的接触而导致生理代谢上的紊乱[11-13,16]。葡萄采后采用钙联合短波紫外处理可延长保鲜期[13],钙联合热水处理可缓解梨的细胞壁代谢,延缓软化[10]。研究发现对葡萄采后进行钙联合涂膜和热水处理,可减少葡萄腐烂率[28]。但在葡萄采后处理方法中,采用钙联合涂膜和钙联合热处理对红提葡萄冷藏期间其果肉组织的能量水平和细胞壁代谢影响自溶软化的研究鲜见报道。
本研究以‘红地球’葡萄为研究对象,对其进行钙联合涂膜和热处理并低温贮藏,以未处理组为对照,探悉果肉自溶指数、硬度、腐烂率、膜透性、果肉组织的能量物质、细胞壁降解相关酶(PG、PE、Cx和β-Gal)活力及细胞壁成分(原果胶、水溶性果胶、纤维素和半纤维素)在贮藏过程中的变化规律,并应用扫描电子显微镜(scanning electron microscope,SEM)对不同处理葡萄在贮藏末期的果实内果皮超微结构进行观察,旨在从能量水平和细胞壁代谢的角度探讨葡萄果肉自溶软化的机理,为延缓采后葡萄果实软化衰老、延长果实保鲜期提供理论依据。
1 材料与方法
1.1 材料与试剂
葡萄(品种为欧亚种‘红地球’葡萄),充分成熟,可溶性固形物含量14%以上,2015年8月20日采自湖南省澧县张公庙镇葡萄园,采前10 d停止施水,采收时间为7∶00~9∶00,采收无病害、无霉变、无机械损伤的果实,采收后装入透气的塑料筐内,并于采收当日运回中南林业科技大学食品科学与工程学院进行风冷12 h,去除田间热,于次日进行不同处理。
5’-磷酸腺苷钠盐(纯度大于98.5%)、5’-二磷酸腺苷钠盐(纯度大于95%)、5’-单磷酸腺苷钠盐(纯度大于99.0%)、聚半乳糖醛酸、果胶、羧甲基纤维素钠、p-硝基酚-β-D-吡喃半乳糖苷(纯度大于99.0%) 西格玛奥德里奇(上海)贸易有限公司;乙腈(色谱纯) 美国天地公司;氯化钙、壳聚糖(均为食品级) 焦作冠通化工有限公司;其他化学试剂均为国产分析纯。实验室用水(18 MΩ)由Milli-Q RG超纯水系统制备。
1.2 仪器与设备
PDA 2010AT高效液相色谱(high performance liquid chromatography,HPLC)仪、UV7100紫外分光光度计日本Shimadzu公司;CT3质构分析(textural profile analysis,TPA)仪 美国Brookf i eld公司;Avanti J-26XP高效冷冻离心机 美国贝克曼库尔特有限公司;AL204电子天平、Delta 320 pH计 梅特勒-托利多仪器(上海)有限公司;Evols 10 SEM 德国卡尔蔡司公司。
1.3 方法
1.3.1 果实采后处理方法
对照组(CK组):葡萄未进行处理,直接装盘覆膜贮藏;钙联合涂膜处理组(CT组):将葡萄在质量分数1%壳聚糖和质量分数0.2% CaCl2溶液中浸泡8 min,果实全部浸没,晾干备用[28];钙联合热处理(HT组):葡萄在45 ℃的质量分数为0.2% CaCl2溶液中浸泡8 min,果实全部浸没,晾干备用[28]。每个处理组3 个重复,每个重复3 个托盘,每个托盘装果2.5 kg,表面覆上保鲜膜,于(4.0±0.5) ℃冷藏。处理结束时,取样一次作为第0天的样品,每隔10 d测定葡萄的自溶指数、硬度、腐烂率、膜透性、能量物质、细胞壁成分和细胞壁降解相关酶活力,测定周期为40 d。
测定自溶指数和腐烂率时从每个重复中取1 个托盘,即每个处理组合计3 个托盘取果进行测定。能量物质、细胞壁组分和降解酶活力测定从每个重复中取1 个托盘,即每个处理组合计3 个托盘,葡萄果粒全部液氮粉碎后放于-70 ℃冰箱内保存,取样进行测定。硬度和膜透性是从每个重复中取1 个托盘,合计3 个托盘进行测定,其中内果皮组织SEM扫描在贮藏初期和贮藏末期进行。
1.3.2 果肉自溶评价
参照赵云峰[4]、林河通[5]等的方法。每次随机取50 个果实,按照果肉自溶面积大小把果肉自溶程度分为5 级。0 级:果肉有弹性、果肉无自溶;1 级:果肉变软、果肉自溶面积<1/4;2 级:果肉变软、流汁,1/4≤果肉自溶面积<1/2;3 级:果肉变软、流汁,1/2≤果肉自溶面积<3/4;4 级:果肉糜烂,果肉自溶面积≥3/4。按照公式(1)计算果肉自溶指数。

1.3.3 内果皮组织SEM扫描
参照刘峰娟等[17]的方法,略有改进。用双刃刀片将‘红地球’葡萄果皮切成1 mm×1 mm×2 mm,环氧树脂包埋,用超薄切片机切片,厚度为60 nm。加入到质量分数为2.5%的戊二醛(0.1 mol/L,pH 6.8磷酸盐缓冲液配制)溶液中,在4 ℃冰箱内固定24 h。经相应的磷酸缓冲液(0.1 mol/L,pH 6.8)漂洗后,用体积分数50%、70%、80%、90%、100%丙酮溶液梯度脱水,各级30 min。再用体积分数为100%的叔丁醇浸没10 min后,放入真空干燥器中干燥40~60 min。干燥后用电导胶分别把样品黏贴在样品台上,黏贴时样品观察面朝上,用镀膜仪镀金膜,于l5 kV加速电压下观察扫描结果。
1.3.4 腐烂率的测定
每10 d对贮藏葡萄进行质量测定,分别测定腐烂果的质量,并按式(2)计算腐烂率。

1.3.5 硬度测定
将保留果梗的葡萄横放于TPA夹具正下方,进行硬度测试,选用的夹具直径为50.8 mm,长20 mm的圆柱形探头TA 25/1000。经预实验选取合适的测试参数:目标类型为TPA实验,距离4 mm,触发点负载50 N,测试速率0.5 mm/s,循环2 次。由质地特征曲线得到表征果实质地状况的力学参数,其中硬度以双峰曲线中第1个峰的最大值表示,单位为N。每次取20 粒果实,分别在果实的对角线取2 个点,进行测定,取平均值。测试温度:室温(18~20 ℃)。
1.3.6 细胞膜透性的测定
用电导仪测定葡萄果肉圆片浸提液的电导值,煮沸后再测定浸提液的电导值,以前后2 次电导值之比所得的相对电导率变化来表示细胞膜透性的大小。
1.3.7 能量物质含量测定
参照Liu Hai等[21]的方法,略作调整。取葡萄果肉组织2 g(-70 ℃冰箱内液氮粉碎的样品),加入10 mL 0.6 mol/L高氯酸,冰浴研磨提取1 min,于4 ℃、16 000×g离心15 min。取5 mL上清液迅速用1.0 mol/L KOH中和至pH 6.5~6.8,冰浴中稳定30 min使高氯酸沉淀,之后经4℃、8 000×g离心5 min,取上清液定容至5 mL,并过0.45 μm微孔滤膜过滤。用HPLC法测定三磷酸腺苷(adenosine triphosphate,ATP)、二磷酸腺苷(adenosine diphosphate,ADP)、单磷酸腺苷(adenosine monophosphate,AMP)的含量。色谱条件为C18反相柱(250 mm×4.6 mm,5.0 μm);检测波长254 nm;流动相:pH 7.0的20 mmol/L磷酸氢二钾缓冲溶液(A相),体积分数60%乙腈(B相),A相与B相为4∶6(V/V),柱温30 ℃,流速0.8 mL/min;进样体积20 μL。定量方法采用外标法,根据标准品保留时间和峰面积进行定性定量[21]。按照公式(3)计算能荷(energy charge,EC)。
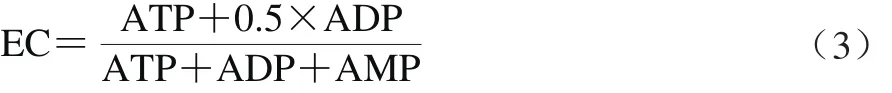
式中:ATP、ADP、AMP分别为果肉组织中的3 种能量物质的含量/(μg/g)。
1.3.8 葡萄果实果肉中细胞壁降解酶的提取和活力的测定
参照赵云峰[4]、王玲利[10]等的方法提取细胞壁降解酶。PE活力以每小时消耗1 μmol NaOH的酶用量为1 个酶活力单位,结果以U/g表示;PG活力以每小时生成1 μmol半乳糖醛酸的酶用量为1 个酶活力单位,结果以U/g表示;Cx活力以每小时生成1 μmol葡萄糖的酶用量为1 个酶活力单位,结果以U/g表示。β-Gal活力以每小时每毫克果肉产生1 μmol硝基酚为一个酶活力单位,结果以U/mg表示。以上各指标均重复测定3 次。
1.3.9 葡萄果实果肉中细胞壁成分的测定
参照赵云峰[4]、王玲利[10]等的方法测定果肉原果胶、水溶性果胶、纤维素、半纤维素等细胞壁组分含量,单位mg/g。
1.4 数据统计分析
数据采用Excel 2010和Origin 8.0软件进行数据统计,采用Tukey方法进行差异显著性分析。
2 结果与分析
2.1 果肉自溶指数、果实硬度、腐烂率、膜透性和内果皮超微结构的变化
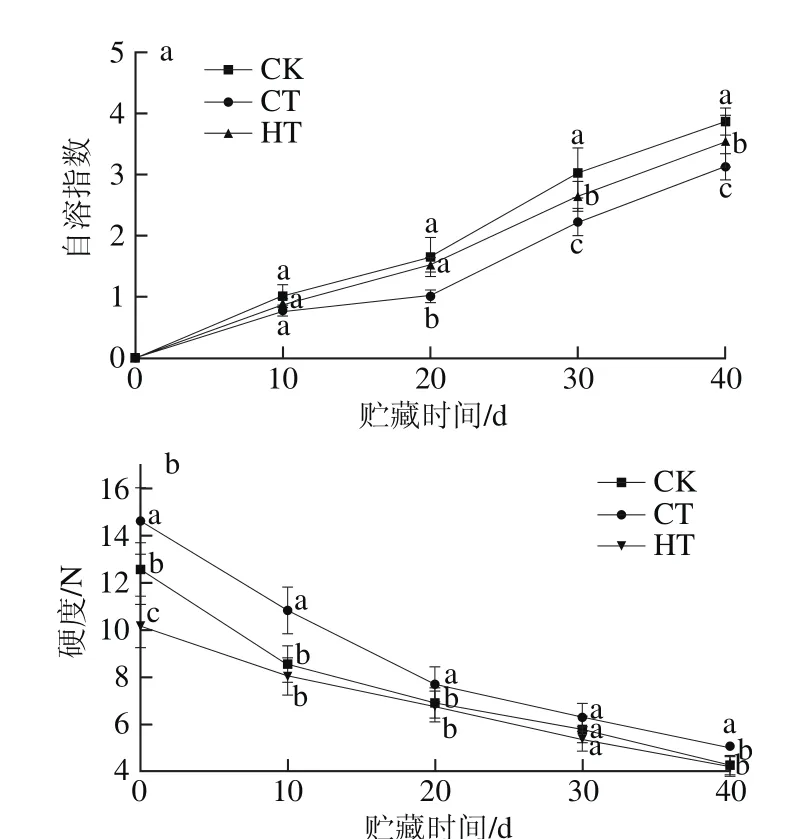

图1 葡萄果实贮藏期间果肉自溶指数(a)、果实硬度(b)、腐烂率(c)和细胞膜透性(d)的变化Fig. 1 Changes in aril breakdown index (a), hardness (b), decay percentage (c) and cell membrane permeability (d) in grape fruits at different storage times
葡萄采后极易出现果肉自溶软化。图1a表明葡萄果肉自溶指数随贮藏时间的延长呈幅度不同的线性增加趋势。3组葡萄果肉自溶指数在0~10 d内无显著差异(P>0.05);贮藏到40 d时,CK组的大部分果肉发生自溶软化和腐烂,CT组葡萄的自溶指数显著比CK组和HT组低(P<0.05),表明钙联合热处理和钙联合涂膜处理均能延缓葡萄果实自溶软化,与热水处理龙眼[4]、热水处理葡萄[14-15]及涂膜处理葡萄的研究结果一致。Wu Ziming[3]、Yuan Xiaozhuan[29]等研究认为热处理减缓软化的部分,原因可能是热处理增加了碳水化合物代谢酶活性,提高了能量物质的供应,维持了高的能量状态,但软化与能量代谢的具体相关性还待进一步的研究。钙联合涂膜延缓自溶软化效果优于钙联合热处理,可能是壳聚糖涂膜处理在葡萄果粒的表面形成一层保护膜,在某种程度上减弱果实的呼吸作用,减少营养物质的消耗,降低蒸腾作用[7,14]。
硬度是果实软化的直观表现。图1b表明,葡萄果实硬度随贮藏时间的延长逐渐下降,与张昆明等[30]对不同保鲜膜包装的葡萄的研究结果一致。图1b显示葡萄果实经不同处理后硬度有所改变,CK组的初始硬度值为(12.57±1.13) N,CT组和HT组的硬度值分别为(14.62±1.41)、(10.17±0.92) N,可见热处理降低了葡萄的初始硬度,这与Wu Ziming[3]、李珍[2]等的研究结果相同。钙联合涂膜处理则增大硬度,可能钙与果胶形成果胶酸钙,涂膜形成保护膜,两者共同作用增大了果实的硬度。在贮藏初期(贮至10 d),CK组、CT组和HT组硬度分别下降了32%、26%和21%,CK组下降最明显。贮藏至40 d,CK组大多数葡萄已经腐烂,不能进行下一步实验。在整个贮藏期间,CT组葡萄果实硬度高于HT组和CK组,这说明钙联合涂膜能够延缓葡萄果实的软化,因为涂膜形成了保护膜,降低营养物质的损耗[2,7,14-15],钙延缓细胞壁物质的降解,从而延缓葡萄果实的软化[11-13,16]。
腐烂率是葡萄感官品质变异程度的一个重要指标,由图1c可看出,随贮藏时间的延长,葡萄腐烂率呈上升趋势。CT组和HT组葡萄贮藏至20 d前未发现腐烂,但CK组则出现了(11.64±1.08)%的腐烂率,这可能是涂膜和热处理增加了果皮和果肉中抗真菌基因的表达诱导并激活耐腐机制,抑制葡萄果实表面微生物的生长繁殖[3,11,14-15]。但贮藏至30 d时,CT组和HT组腐烂率分别为(9.86±0.84)%和(15.84±1.48)%,而CK组为(30.26±2.84)%,是处理组的2 倍多,不同组间腐烂率差异显著(P<0.05)。贮藏至40 d时,各组的腐烂率均快速上升,CK组达(69.72±6.42)%,大多数葡萄已不能食用,CT组为(38.82±3.62)%,HT组为(48.82±4.56)%,处理组间差异显著(P<0.05),HT组比CT组高约10%,可能热处理后葡萄果实的起始硬度下降,红提汁多,水分高,加速果实的软化和腐烂[2-3]。
果实相对电导率大小可表示细胞膜透性的大小[20-27]。由图1d可知,‘红地球’葡萄果实细胞膜的相对电导率随贮藏时间的延长而增加。在同一贮藏时间,3 组的相对电导率从大到小是CK组>HT组>CT组,三者差异显著(P<0.05)。可见CT组和HT组显著抑制了膜渗透性的上升(P<0.05),保持了细胞膜良好的完整性,延缓果实衰老软化,在减轻葡萄果实自溶软化方面起到了有效的作用[3,11,14-15],且钙联合涂膜效果要优于钙联合热处理。
统计分析表明,葡萄自溶指数与组织细胞膜透性呈极显著正相关(P<0.01),可能因为细胞膜透性增加,细胞壁组分与降解酶的区室化分布破坏,大分子可以进入,促进了相关酶如果胶酶等与底物接触而加速软化[4-5,7]。
葡萄自溶软化是一种品质劣变的过程,在这个过程中葡萄组织内部的超微结构发生变化。果实细胞胞间层结构改变,细胞壁总体结构破坏以及胞壁物质降解,宏观上果实变软,微观上表现为细胞壁微丝排列由紧密有序变得松散无序,并部分分解,中胶层电子密度由致密变稀松,细胞壁不均匀,出现许多波形皱褶[4]。
图2为贮藏初期和贮藏末期3 组葡萄内果皮超微SEM扫描结果。贮藏初期(图2d),葡萄内果皮组织结构整齐致密、分布均匀,有清晰的明暗结构,没有孔洞和沟壑,与刘峰娟等[17]对新鲜葡萄SEM结果一致。但在贮藏40 d后,不同处理的葡萄内果皮组织有差别(图2a~c)。CK组葡萄品质严重下降,葡萄果肉组织稀松,有很多较深较大孔洞,已失去食用价值,与赵云峰等[4]对龙眼果实贮藏期间果肉自溶时细胞形态和组织结构发生了显著变化的结果一致;CT组果肉组织较平滑、结构较紧密;HT组的果肉组织叠层较CT组多,且更稀疏,因为钙联合涂膜可在葡萄表面形成一层膜,使果蔬内部形成一个低O2、高CO2的微环境,可以起到更好的保护作用,从而更好地延缓软化;但两个处理组内果皮组织超微结构显示中均没有出现较深孔洞,可能是由于两种处理能降低葡萄果肉细胞壁降解酶活力,从而减缓细胞壁组分的降解,延缓果肉自溶发生。
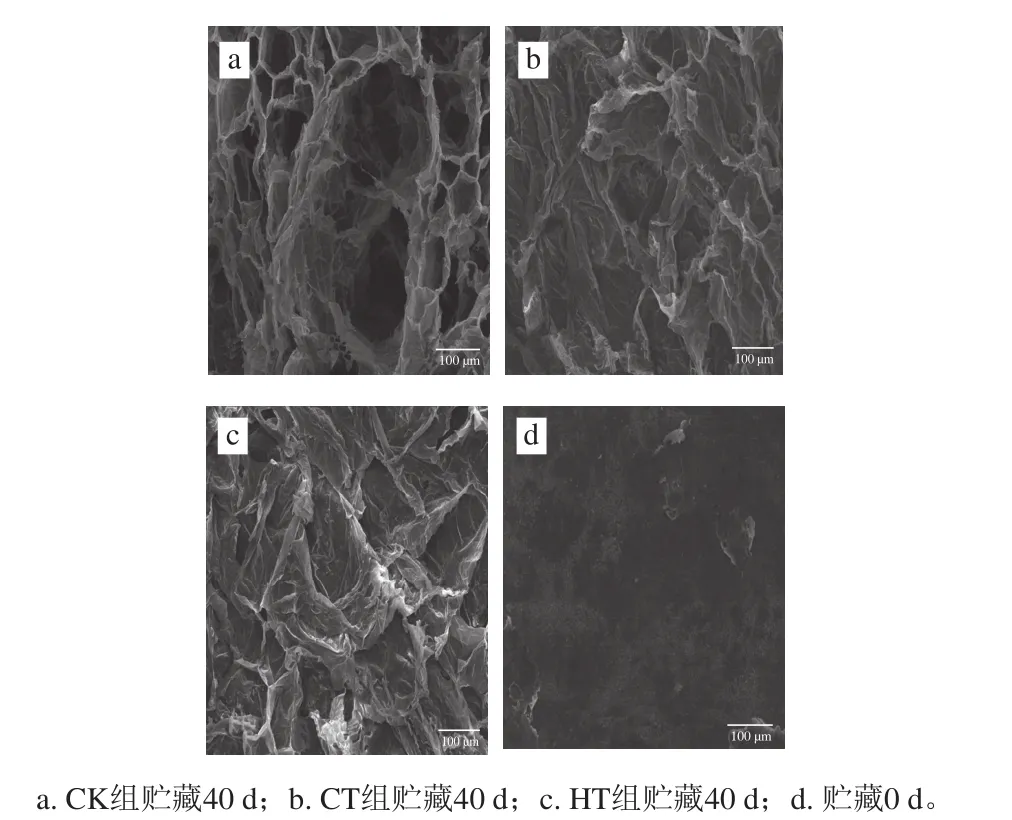
图2 不同处理的葡萄内果皮组织的超微结构(×200)Fig. 2 Ultrastructure of inner pericarp in grape fruits with different treatments before and after 40 days of storage (×200)
2.2 葡萄果肉组织的ATP、ADP、AMP含量及EC与自溶软化的关系
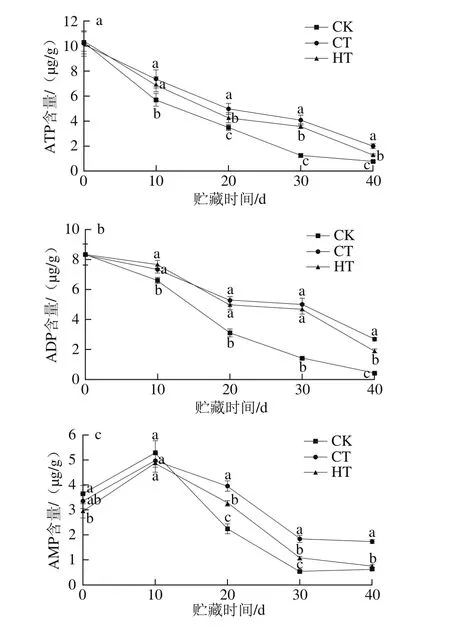

图3 不同贮藏时间葡萄果实中ATP(a)、ADP(b)、AMP(c)含量和EC(d)的变化Fig. 3 Changes in ATP (a), ADP (b) and AMP (c) contents, and EC (d)of grape fruits at different storage times
采收时葡萄果肉组织有高水平的ATP、ADP含量和EC值(图3a、b、d),相对低水平的AMP含量(图3c),这与桃、荔枝、芒果、油木奈、梨贮藏初期能量物质结果一致[20-27,31]。因为ATP、ADP和EC在合成细胞膜脂肪酸中发挥主要作用,细胞膜中不饱和脂肪酸的变化可导致细胞膜性质发生变化,如膜透性增加、细胞衰老[32]。
在整个贮藏期间,ATP含量随贮藏时间的延长不断下降。CK组与处理组间葡萄组织中ATP含量差异显著(P<0.05),CT组ATP含量在贮藏后期(20~40 d)显著高于HT组(P<0.05)。处理组ADP含量的变化与ATP含量变化类似。由图3b可以看出,在整个贮藏期间,除40 d外,处理组间ADP含量无显著性差异(P>0.05),但均比CK组高,可能因为适当地处理在某种程度上可减弱果实的呼吸作用,保持低的呼吸水平[3,14,31]。葡萄果肉组织中AMP含量在10 d内增加,然后下降。贮藏至20~30 d时,AMP含量下降,3 组间差异显著(P<0.05)。在贮藏中后期(10~40 d),处理组的葡萄组织中AMP含量要高于CK组,其中CT组AMP含量显著高于HT组(P<0.05),但CK组与HT组间无显著差异(P>0.05)。
综上所述,钙联合涂膜比钙联合热处理能更好地延缓ATP、ADP、AMP含量下降,果肉组织处于高能量状态,且自溶指数和腐烂率低(2.1节),部分原因可能是由于经过钙联合涂膜处理后葡萄果实中碳水化合物代谢的酶活力增加,加强了能量物质的供应[14-15]。不同处理如何维持高水平的能量及保持葡萄硬度的机理待进一步研究。
EC大小反映细胞中腺苷酸系统的能量状态,是细胞内最重要的能量转换与调节系统[20-27,31]。由图3d可知,3 组葡萄的EC变化幅度为0.51~0.65,总体呈下降趋势。贮藏10~30 d,EC有小幅的上升,因在此期间,AMP含量下降幅度大于ATP和ADP含量。在贮藏末期,EC值缓慢下降,HT组EC最高,CT组与CK组无显著性差异(P>0.05)。
葡萄采后果肉软化与能量水平的关系鲜见报道,但果蔬的衰老劣变与能量代谢有密切关系[19-28,31]。统计分析表明,果肉自溶指数与ATP、ADP含量呈极显著负相关(P<0.01),与AMP含量呈显著负相关(P<0.05)。葡萄硬度与ATP含量呈极显著正相关(P<0.01),与ADP、AMP含量呈显著正相关(P<0.05),与EC无明显相关性(P>0.05)。所以,葡萄果实自溶软化及硬度的下降与能量水平密切相关,ATP、ADP、AMP水平降低,果实自溶软化不断加剧,其机理可能是在果实衰老软化中,细胞膜透性增加,膜完整性受损,膜上的能量代谢酶的功能丧失,影响了能量的供应[19-28,31]。
2.3 果肉细胞壁降解酶活力的变化与自溶软化的关系

图4 葡萄果实贮藏期间果肉组织中PG(a)、PE(b)、Cx(c)和β-Gal(d)活力的变化Fig. 4 Changes in PG (a), PE (b), Cx (c) and β-Gal (d) activities in grape fruits at different storage times
PG的主要功能是水解果实细胞壁中果胶酸的1,4-2-D-半乳糖苷酶,生成低聚半乳糖醛酸,导致细胞壁解体,最终使果实软化[4-10,16]。葡萄果肉组织的PG活力在贮藏过程中逐渐上升(图4a)。采后贮藏至10 d内,3 组间PG活力无显著差异(P>0.05);贮藏10~30 d时,CT组PG活力显著低于CK、HT组(P<0.05),CK和HT组间差异不显著(P>0.05);贮藏至40 d时,葡萄果肉组织的PG活力大小依次为CK组>HT组>CT组,3 组差异显著(P<0.05)。可见,在贮藏中后期钙联合涂膜和钙联合热处理均可抑制PG活力上升。Ca2+能维持细胞内膜系统的区域化,抑制内质网上细胞裂解酶到达作用位点,延缓细胞壁物质降解[10,12],从而延缓果肉自溶软化和腐烂。适宜强度的热处理可使PG活力降低,抑制果实水溶性果胶含量的上升和非水溶性果胶含量的下降,从而达到延缓果实软化的目的[3-4,6,10]。涂膜处理有效抑制PG活力,延缓原果胶向可溶性果胶转变[3-4,6,10,15]。但葡萄果实经热处理后,外果皮受热收缩,气孔关闭,合成的热激蛋白形成了一层保护膜[4,10],Ca2+进入果肉组织受阻,因此,热处理抑制PG活力的能力不如涂膜组(P<0.05)。
PE对果胶物质的降解和果肉自溶起辅助作用,其生理意义在于为PG准备作用底物,果胶的降解是在PG和PE的协同作用下[4-8,10,12]。采后葡萄果肉组织PE活力总体上升(图4b)。贮藏初期PE活力为(87.14±7.72) U/g;贮藏至10 d时,CK组、HT组、CT组果肉组织的PE活力增幅分别为189.99%、159.95%和127.55%,且差异显著(P<0.05),贮藏至40 d时,3 组葡萄果肉组织的PE活力无显著性差异(P>0.05)。在整个贮藏期间,CK组果肉的PE活力均大于处理组,合适的处理可抑制PE活力上升,钙联合涂膜抑制效果优于钙联合热处理。
Cx与细胞壁降解和质地软化启动的关系密切,Cx引起纤维素以及难溶性的半纤维素向易溶的半纤维素转化,直接导致细胞壁中P—C—H“经纬结构松散”,导致果肉出现软化[4-10]。葡萄果肉中Cx活力呈线性升高(图4c),造成纤维素降解、细胞壁解体和果实软化。贮藏初期葡萄果实Cx活力仅为(49.02±3.64) U/g;在贮藏中期,其活力为CT组<HT组<CK组,表明合适的处理在一定程度上抑制了贮藏中期Cx活力的上升,抑制细胞壁的降解,降低了果实软化的速率,与赵云峰等[4]对龙眼的热处理和罗自生等[7]对梨果实涂膜处理结果一致,但钙联合涂膜抑制效果优于钙联合热处理。贮藏结束时,葡萄果实Cx活力分别为CK组(75.28±4.18) U/g、CT组(76.86±7.70) U/g、HT组(75.84±5.84 )U/g,三者无显著差异(P>0.05)。
β-Gal通过降解果胶多聚醛酸侧链的半乳糖残基,降解果胶聚合体,破坏细胞壁结构,从而使果实软化[4-5]。从图4d中可知,采后葡萄果肉β-Gal活力在贮藏0~30 d内呈下降趋势,后30~40 d持续升高,且CT组>HT组>CK组,与赵云峰等[4]对龙眼果实的研究结果一致。但葡萄果肉组织的β-Gal活力与自溶指数无明显相关性(P>0.05),这与薛炳烨等[33]对草莓的研究结果类似,说明β-Gal不是引起葡萄果肉自溶软化的主要降解酶,与β-Gal在龙眼自溶软化后期起重要作用结果不一致[4-5],具体原因还待进一步的研究分析。
本研究中葡萄果肉自溶指数与PG、PE、Cx活力呈极显著正相关(P<0.01),所以相关细胞壁降解酶(PE、PG和Cx)影响采后葡萄果肉自溶软化,与龙眼、葡萄、杨桃和草莓软化的关键因子结果一致[4-6,8,33]。而且钙联合涂膜和热处理可抑制降解酶活力的上升,延缓果实自溶软化,钙联合涂膜处理的抑制效果优于钙联合热处理。
2.4 果肉细胞壁组分变化与自溶软化的关系

图5 葡萄果实贮藏期间果肉原果胶(a)、水溶性果胶(b)、纤维素(c)和半纤维素(d)含量的变化Fig. 5 Changes in protopectin (a), water soluble pectin (b), cellulose (c)and semi-cellulose (d) contents in grape fruits during storage
果肉组织胞间层结构改变造成细胞分离和胞壁物质降解,细胞间黏合力下降,从而引起果实软化[4-10,12]。如图5a所示,随贮藏时间延长,原果胶含量降低,CK、CT和HT组果肉中原果胶含量由贮藏前(1.88±0.12)mg/g分别降至(0.45±0.046)、(0.86±0.082)、(0.68±0.06) mg/g。可见,适当地处理可延缓原果胶下降,自溶指数与原果胶含量呈极显著负相关(P<0.01),与龙眼自溶结果一致[4-5]。PG和PE协同作用促进原果胶降解,原果胶下降与PG、PE活力升高呈极显著负相关(P<0.01)。如图5b所示,水溶性果胶含量先上升,在第20天时达高峰,之后逐渐下降,高峰时CK组、HT组和CT组水溶性果胶含量分别为采收时的1.31、1.19、1.10 倍;40 d时CK组中水溶性果胶含量为(0.34±0.02) mg/g、CT组为(1.38±0.14) mg/g、HT组为(0.90±0.08) mg/g,三者差异显著(P<0.05)。统计分析表明,葡萄果肉自溶指数与水溶性果胶含量呈极显著负相关(P<0.01)。
果肉自溶软化不仅与果胶降解有关,而且与纤维素的结构变化有关[4-10,12]。在整个贮藏期间,果肉纤维素和半纤维素含量快速下降(图5c、d),纤维素和半纤维素的分解为果胶质的降解提供重要的条件,与龙眼自溶结果一致[4-5]。贮藏至40 d时,CK组、CT组和HT组的葡萄果肉组织中纤维素含量由贮前的(2.84±0.22) mg/g分别下降到(0.04±0.00)、(0.57±0.07)、(0.45±0.04) mg/g,下降幅度达99%、80%和84%,采后处理可缓解纤维素含量下降,但CT组纤维素含量高于HT组和CK组。贮藏初期半纤维素含量为(1.24±0.23) mg/g;贮藏至30 d,三者半纤维素含量差异显著(P<0.05),葡萄果肉组织中半纤维素含量大小依次是CT组>HT组>CK组;贮藏至40 d时,CK组、CT组和HT组的葡萄果肉组织中半纤维素含量分别下降到(0.00±0.00)、(0.24±0.16) mg/g和(0.01±0.01) mg/g,下降幅度达100%、64%和99%,CT组与CK、HT组间差异显著(P<0.05),但CK组和HT组间差异不显著(P>0.05)。统计分析表明,葡萄果肉自溶指数与纤维素、半纤维素含量呈极显著负相关(P<0.01),这与龙眼自溶结果一致[4-5]。适当的采后处理可使纤维素酶活力降低,果胶质与纤维素不易分离,果胶质不能顺利分解,纤维素含量高,自溶指数低,这与龙眼自溶结果一致[4-5]。
3 结 论
本研究比较了钙联合涂膜处理、钙联合热处理和未处理3 组葡萄果实低温贮藏下果肉自溶软化与果肉组织能量水平和细胞壁代谢的变化及相互关系,且进行了贮藏初期和末期果实内果皮组织的超微结构SEM观察。研究认为随贮藏时间的延长,葡萄果肉出现自溶软化、硬度下降、腐烂率升高、能量水平下降、细胞膜透性增大、细胞膜完整性被破坏、内果皮组织结构出现大的空洞、细胞超微结构被破坏、细胞壁降解酶与底物的区室化作用被破坏,引起果肉分解软化。自溶指数与ATP、ADP呈极显著负相关(P<0.01),与AMP呈显著负相关(P<0.05),与EC水平无明显相关性(P>0.05),与细胞壁降解酶(PG、PE和Cx)活力呈极显著正相关(P<0.01),与β-Gal活力无明显相关性(P>0.05)。自溶指数与细胞壁组分(原果胶、水溶性果胶、纤维素和半纤维素)含量呈极显著负相关(P<0.01)。表明葡萄果肉衰老中的自溶软化与果肉组织结构、能量水平和细胞壁代谢特性有密切关系。
钙联合涂膜和钙联合热处理可减轻葡萄内果皮超微结构的损伤,较好地保持细胞超微结构的完整性。钙联合涂膜和钙联合热处理缓解膜透性增加,维持葡萄组织高能量水平,抑制细胞壁降解酶降解细胞壁组分,延缓葡萄果肉组织的自溶软化;但钙联合涂膜处理在延缓果肉自溶软化和维持高的能量水平和低的细胞壁代谢能力方面要显著优于钙联合热处理(P<0.05)。
[1] 李志文, 张平, 刘翔, 等. 1-MCP 结合冰温贮藏对葡萄采后品质及相关生理代谢的调控[J]. 食品科学, 2011, 32(20): 300-306.
[2] 李珍, 哈益明, 李咏富, 等. 不同处理对红提葡萄冷藏品质的影响[J].中国食品学报, 2015, 15(1): 123-128.
[3] WU Ziming, YUAN Xiaozhuan, LI Huan, et al. Heat acclimation reduces postharvest loss of table grapes during cold storage-analysis of possible mechanisms involved through a proteomic approach[J].Postharvest Biology and Technology, 2015, 105(7): 26-33.DOI:10.1016/j.postharvbio.2015.03.012.
[4] 赵云峰, 林河通, 王静, 等. 热处理抑制采后龙眼果肉自溶及细胞壁物质降解[J]. 农业工程学报, 2014, 30(11): 268-275. DOI:10.3969.issn.1002-6819.2014.11.033.
[5] 林河通, 赵云峰, 席玙芳. 龙眼果实采后果肉自溶过程中细胞壁组分及其降解酶活性的变化[J]. 植物生理与分子生物学学报, 2007,33(2): 137-145.
[6] 寇莉萍, 刘兴华, 任亚梅, 等. 热处理对轻度加工葡萄细胞壁组分及相关酶活性的影响[J]. 保鲜与加工, 2008, 44(1): 21-24.
[7] 罗自生, 寿浩林. NSCC涂膜对黄花梨软化和细胞壁代谢的影响[J].果树学报, 2011, 28(1): 143-146.
[8] 陈艺晖, 张华, 林河通, 等. 1-甲基环丙烯处理对采后杨桃果实软化和细胞壁代谢的影响[J]. 热带亚热带植物学报, 2013, 21(6): 566-571. DOI:10.3969/j.jssn.1005-3395.2013.06.011.
[9] 赵博, 饶景萍. 柿果实采后胞壁多糖代谢及其降解酶活性的变化[J].西北植物学报, 2005, 25(6): 1199-1202.
[10] 王玲利, 刘超, 黄艳花, 等. ‘黄冠梨’采后热处理和钙处理对其钙形态及细胞壁物质代谢的影响[J]. 园艺学报, 2014, 41(2): 249-258.
[11] CICCARESE A, STELLACCI A M, GENTILESCO G, et al.Effectiveness of pre- and post-veraison calcium applications to control decay and maintain table grape fruit quality during storage[J].Postharvest Biology and Technology, 2013, 75(1): 135-141.DOI:10.1016/j.postharvbio.2012.08.010.
[12] ORTIZ A, GRAELL J, LARA I. Cell wall-modifying enzymes and firmness loss in ripening ‘golden reinders’ apples: a comparison between calcium dips and ULO storage[J]. Food Chemistry, 2011,128(4): 1072-1079. DOI:10.1016/j.foodchem.2011.04.016.
[13] WANG L J, LING M A, XI H F, et al. Individual and combined effects of CaCl2and UV-C on the biosythesis of reservatrols in grape leaves and berry skins[J]. Journal of Agricultural and Food Chemistry, 2013,61(29): 7135-7141. DOI:10.1021/jf401220m.
[14] MENG X H, LI B, LIU J, et al. Physiological responses and quality attributes of table grape fruit to chitosan preharvest spray and postharvest coating during storage[J]. Food Chemistry, 2008, 106(2):501-508. DOI:10.1016/j.foodchem.2007.06.012.
[15] DE OLIVEIRA C E V, MAGNANI M, DE SALES C V, et al. Effects of post-harvest treatment using chitosan from Mucor circinelloides on fungal pathogenicity and quality of table grapes during storage[J]. Food Microbiology, 2014, 44(6): 211-219. DOI:10.1016/j.fm.2014.06.007.
[16] 王贵禧, 韩雅珊, 于梁. 浸钙对猕猴桃果实硬度变化影响的生化机制[J]. 园艺学报, 1995, 22(1): 21-24.
[17] 刘峰娟, 冯作山, 孟阳, 等. 脱水速度对‘无核白’葡萄果皮褐变和细胞超微结构的影响[J]. 食品科学, 2016, 37(6): 220-225.DOI:10.7506/spkx1002-6630-201606040.
[18] 周会玲, 李嘉瑞. 葡萄果实组织结构与耐贮性的关系[J]. 园艺学报,2006, 33(1): 28-32.
[19] 赵颖颖, 陈京京, 金鹏, 等. 低温预贮对冷藏桃果实冷害及能量水平的影响[J]. 食品科学, 2012, 33(4): 276-281.
[20] WANG H, QIAN Z, MA S, et al. Energy status of ripening and postharvest senescent fruit of Litchi (Litchi chinensis Sonn.)[J]. BMC Plant Biology, 2013, 13(1): 1-16. DOI:10.1186/1471-2229-13-55.
[21] LIU Hai, SONG Lili, YOU Yanli, et al. Cold storage duration affects litchi fruit quality, membrane permeability, enzyme activities and energy charge during shelf time at ambient temperature[J]. Postharvest Biology and Technology, 2011, 60(1): 24-30. DOI:10.1016/j.postharvbio.2010.11.008.
[22] 陈京京, 金鹏, 李会会, 等. 低温贮藏对桃果实冷害和能量水平的影响[J]. 农业工程学报, 2012, 28(4): 275-281. DOI:10.3969/j.issn.1002-6819.2012.04.046.
[23] LI P Y, ZHENG X L, LIU Y, et al. Pre-storage application of oxalic acid alleviates chilling injury in mango fruit by modulating proline metabolism and energy status under chilling stress[J]. Food Chemistry,2014, 142(1): 72-78. DOI:10.1016/j.foodchem.2013.06.132.
[24] SAQUET A A, STREIF J, BANGERTH F. Energy metabolism and membrane lipid alterations in relation to brown heart development in‘Conference’ pears during delayed controlled atmosphere storage[J].Postharvest Biology and Technology, 2003, 30(2): 123-132.DOI:10.1016/S0925-5214(03)00099-1.
[25] 程顺昌, 魏宝东, 朱益鹏, 等. 1-MCP和CO2对‘南果梨’冷藏后货架期能量代谢特性的影响[J]. 西北植物学报, 2013, 33(6): 1177-1182.
[26] 李辉, 林毅雄, 林河通, 等. 1-MCP延缓采后‘油木奈’果实衰老及其与能量代谢的关系[J]. 现代食品科技, 2015, 31(4): 121-127.DOI:10.13982/j.mfst.1673-9078.2015.4.020.
[27] 祝美云, 白欢, 梁丽松, 等. 冷锻炼处理减轻低温贮藏桃果实冷害的能量代谢机理[J]. 农业工程学报, 2012, 28(23): 257-264.DOI:10.3969/j.jssn.1002-6819.2012.23.034.
[28] 张群, 周文化, 谭欢, 等. 涂膜和热处理对葡萄能量水平和贮藏生理指标及品质的影响[J]. 农业工程学报, 2016, 32(9): 255-262.DOI:10.11975/j.issn.1002-6819.2016.09.036.
[29] YUAN Xiaozhuan, WU Ziming, LI Huan, et al. Biochemical and proteomic analysis of ‘kyoho’ grape (Vitis labruscana) berries during cold storage[J]. Postharvest Biology and Technology, 2014, 88(2): 79-87. DOI:10.1016/j.postharvbio.2013.10.001.
[30] 张昆明, 张平, 李志文, 等. 葡萄贮藏期间果肉质地参数变化规律的TPA表征[J]. 食品与生物技术学报, 2011, 30(3): 353-358.
[31] KAN J, WANG H M, JIN C H. Changes of reactive oxygen species and related enzymes in mitochondrial respiration during storage of harvested peach fruits[J]. Agricultural Sciences in China, 2011, 10(1):149-158. DOI:10.1016/S1671-2927(11)60317-9.
[32] DUAN X W, JIANG Y M, SU X G, et al. Role of pure oxygen treatment in browning of litchi fruit after harvest[J]. Plant Science,2004, 167(3): 665-668. DOI:10.1016/j.plantsci.2004.05.009.
[33] 薛炳烨, 毛志泉, 束怀瑞. 草莓果实发育成熟过程中糖苷酶和纤维素酶活性及细胞壁组成变化[J]. 植物生理与分子生物学学报, 2006,32(3): 363-368.
Effects of Energy Level and Cell Wall Metabolism on Aril Breakdown in Grape Fruits
ZHANG Qun1,2, ZHOU Wenhua1,*, TAN Huan2, LIU Wei2
(1. College of Food Science and Engineering, Central South University of Forestry and Technology, Changsha 410004, China;2. Hunan Agricultural Product Processing Institute, Hunan Academy of Agricultural Sciences, Changsha 410125, China)
In order to understand the relationship of aril breakdown with energy levels and cell wall metabolism in harvested grapes, the ultrastructural changes in the inner pericarp were examined during the early and late stages of storage.‘Red globe’ grapes were subjected to CaCl2immersion combined with chitosan coating or heat treatment and then stored at (4.0 ± 0.5) ℃, and the untreated samples were used as a control. Changes in aril breakdown index, hardness, decay percentage, cell membrane permeability, energy contents (adenosine triphosphate (ATP), adenosine diphosphate (ADP) and adenosine monophosphate (AMP)), energy charge (EC), cell wall-degrading enzyme activities and cell wall components were monitored at regular intervals during storage. The results indicated that aril breakdown index, decay incidence and cell membrane permeability increased, hardness and energy contents declined, and the activities of the cell walldegrading enzymes polygalacturonase (PG), pectinesterase (PE) and cellulase (Cx) ascended with storage time; however,β-galactosidase (β-Gal) activity dropped during the fi rst 30 days, followed by a sharp increase. The contents of cell wall components such as protopectin, cellulose and semi-cellulose continuously decreased during storage, while water-soluble pectin dropped after an initial increase. Furthermore, the ultrastructure of the inner pericarp was damaged as indicated by the appearance of large holes. Aril breakdown index showed a highly signif i cant inverse correlation with ATP and ADP contentsand the contents of cell wall components (P < 0.01), a highly signif i cant positive correlation with PG, PE and Cx activities(P < 0.01), and a signif i cant inverse correlation with AMP level (P < 0.05), but it had no signif i cant correlation with EC or β-Gal activity (P > 0.05). In conclusion, combined treatment with calcium chloride and chitosan or heat could delay aril breakdown in grapes by maintaining the integrity of the cell membrane and high energy status, inhibiting the transformation of protopectin to water-soluble pectin, inhibiting the activities of cell wall-degrading enzymes (PG, PE and Cx), and suppressing the degradation of cell wall components (proto-pectin, cellulose and semi-cellulose), and chitosan coating was signif i cantly more effective than heat treatment (P < 0.05).
grape fruit; aril breakdown; energy levels; ultrastructure; cell wall metabolism
10.7506/spkx1002-6630-201801040
TS255.3
A
1002-6630(2018)01-0264-09
张群, 周文化, 谭欢, 等. 葡萄果肉组织的能量水平和细胞壁代谢对其自溶软化的影响[J]. 食品科学, 2018, 39(1):264-272.
10.7506/spkx1002-6630-201801040. http://www.spkx.net.cn
ZHANG Qun, ZHOU Wenhua, TAN Huan, et al. Effects of energy level and cell wall metabolism on aril breakdown in grape fruits[J]. Food Science, 2018, 39(1): 264-272. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201801040. http://www.spkx.net.cn
(1.中南林业科技大学食品科学与工程学院,湖南 长沙 410004;2.湖南省农业科学院湖南省农产品加工研究所,湖南 长沙 410125)
2016-10-20
湖南省重点研发计划(农业支撑领域)项目(2015NK3027)
张群(1972—),女,研究员,博士研究生,研究方向为果蔬贮藏与加工。E-mail:zqun208@163.com
*通信作者简介:周文化(1969—),男,教授,博士,研究方向为农产品加工与贮藏。E-mail:13786129879@126.com
猜你喜欢
塑料包装(2021年3期)2021-01-25
三农资讯半月报(2020年18期)2020-10-14
数码世界(2018年1期)2018-12-23
食品研究与开发(2018年8期)2018-05-07
浙江工业大学学报(2017年5期)2018-01-22
食品科学(2018年2期)2018-01-04
中国调味品(2017年10期)2017-10-18
广东农业科学(2017年5期)2017-08-29
中国酿造(2014年5期)2014-04-24
食品工业科技(2014年21期)2014-03-11
