
长白山系榛子灌木林根系对优先流的影响
2018-01-02 09:31王赵男赵雨森
林业科学研究 2017年6期
王赵男,辛 颖*,赵雨森
(东北林业大学林学院,黑龙江 哈尔滨 150040)
长白山系榛子灌木林根系对优先流的影响
王赵男1,辛 颖1*,赵雨森1
(东北林业大学林学院,黑龙江 哈尔滨 150040)

根长密度;根生物量;优先流;基质流;
土壤优先流是一种常见的土壤水分和溶质快速运移的现象,其研究理论被越来越多的土壤学家、水分地质学家和森林水文学家关注[1],开展优先流的研究,有利于了解降雨对地下水位的影响、地表径流和壤中流的形成机制。土壤地下水分和溶质的运移一直是土壤学和水文学研究的重点和难点。
林木根系是土壤层中的重要组成结构,对土壤理化性质有明显的改善作用[2],根系通过穿插、网络和固结将土壤单粒级有机物质粘结起来改善土壤团聚体结构和孔隙性[3],形成了微型的拦土过滤筛,阻止了土粒的搬运和滚动,加强了水分和溶质的运移[4]。根长密度、根质量密度和根生物量是林木根系结构的重要指标,影响优先流路径发生和发展的过程。目前国内外对森林生态系统根系对优先流的路径分布的研究较多,但没有明确其优先流类型,而不同径级根系对优先流的贡献率需要进一步明确。
1 研究地区与研究方法
1.1 研究区概况
1.2 研究方法
1.2.1 样地的选择和实验设备的设置 在帽儿山实验林场老山试验站,选取地势较为平坦,遭受重大破坏后恢复较好的榛子灌木林。设置面积为400 m2(20 m×20 m)的试验观测样地。机械布设3个样点放置实验专用不锈钢铁框和模拟降雨器,确保每个样点与周围灌木距离相距1 m且数量大体相同。不锈钢铁框长×宽×高(1.0 m×1.0 m×0.6 m),放入土中0.5 m,模拟降雨器由4根内直径2.5 cm长1 m不锈钢管,每隔0.2 m处安装一个1.2 mm的雾状喷头,组成的相互连通等压的1 m×1 m的降雨器,距离铁框内部土壤平面0.5 m。
1.2.2 染色示踪实验及图片的获得 于2015年9月27日—10月5日进行试验。选用浓度为4.0 kg·m-3[7-9]体积25 L(50 mm为当地大雨降雨量)。亮蓝染色剂(Brilliant Blue FCF),利用模拟降雨器喷洒在选好的样点内,喷洒完毕后用2 m×2 m透明塑料布将铁框密封,防止再降雨和枯枝落叶的落入[10]。24 h后撤去塑料,开挖剖面,用佳能相机拍摄染色剖面,图像以JPG格式储存。以10 cm为一层,用标准环刀(100 cm3)在剖面10 cm处取原状土柱,分染色与未染色,3个重复,(每层6个),最终剖面修成80 cm×80 cm×60 cm的矩形剖面。纵向每隔2 cm为准,用相机拍摄剖面,每个剖面拍摄3次,直至染色消失。纵剖面规格:80 cm×60 cm×80 cm。
1.2.4 根系的获得及处理 在纵向切剖面的同时,将横向每10 cm的土层内的所有根系全部取出带回,在实验室内将根系样品分成染色与未染色两类,用80目筛子在流水中冲洗,洗好后用滤纸吸干[13],用电子游标卡尺测量并分成d<1 mm,1
1.2.5 根贡献率的测定 先分别算出剖面内5种不同根系径级的优先流区和基质流区的总根长密度,之后用每个根系径级的优先流区的根长密度与该根系径级的总根长密度之比得到该根系径级的优先流区所占的比率即为该根系对优先流的贡献率。
2 结果与分析
2.1 优先流的空间分布
对研究点内的拍摄后的照片经过Image-ProPlus6.0等图像处理软件的处理,提取出研究点内的优先流路径数量和优先流的空间分布情况,见图1。
实际的绝热加速量热仪实验中,受限于样品热电偶动态特性及炉体加热系统响应时间,在样品绝热反应阶段,样品和炉体之间存在一定温度差,使式(2)不能完全满足,导致动力学求解误差。为了量化这一温度差对动力学求解的影响,本文通过绝热反应过程的建模仿真对其进行分析。

图1 优先流区的二维与三维纵向剖面提取信息Fig1 2D and 3D vertical profiles of the preferential flow area
图1纵向二维,三维剖面图,白色为未染色区,黑色为染色区[14],三维图中柱状长短代表该处染色强度(长为弱,短为强)。
土壤表层染色较为均匀,染色剂呈网状扩散式蔓延并绕过基质区快速下渗,形成极其明显的“优先路径”,整体呈现扭曲环绕特征。染色区域内的“空点”多而密,形成连贯的曲线与直线相互连通的网状通路。染色剖面的“优先路径”大多伴随着土壤内的植物根系、石砾、裂隙出现,图中可以看到明显的沿根系染色和绕过石砾的痕迹。土壤优先流类型主要为大孔隙流,漏斗流和环绕流,并出现侧向入渗现象。
2.2 根长密度对优先流的影响
根系结构特征是一项很重要的指标[18],而单位面积上的根长(根长密度)是衡量根系结构特征的指标之一,决定根系吸收养分和水分的能力。不同土层深度的不同根系径级密度见图2。
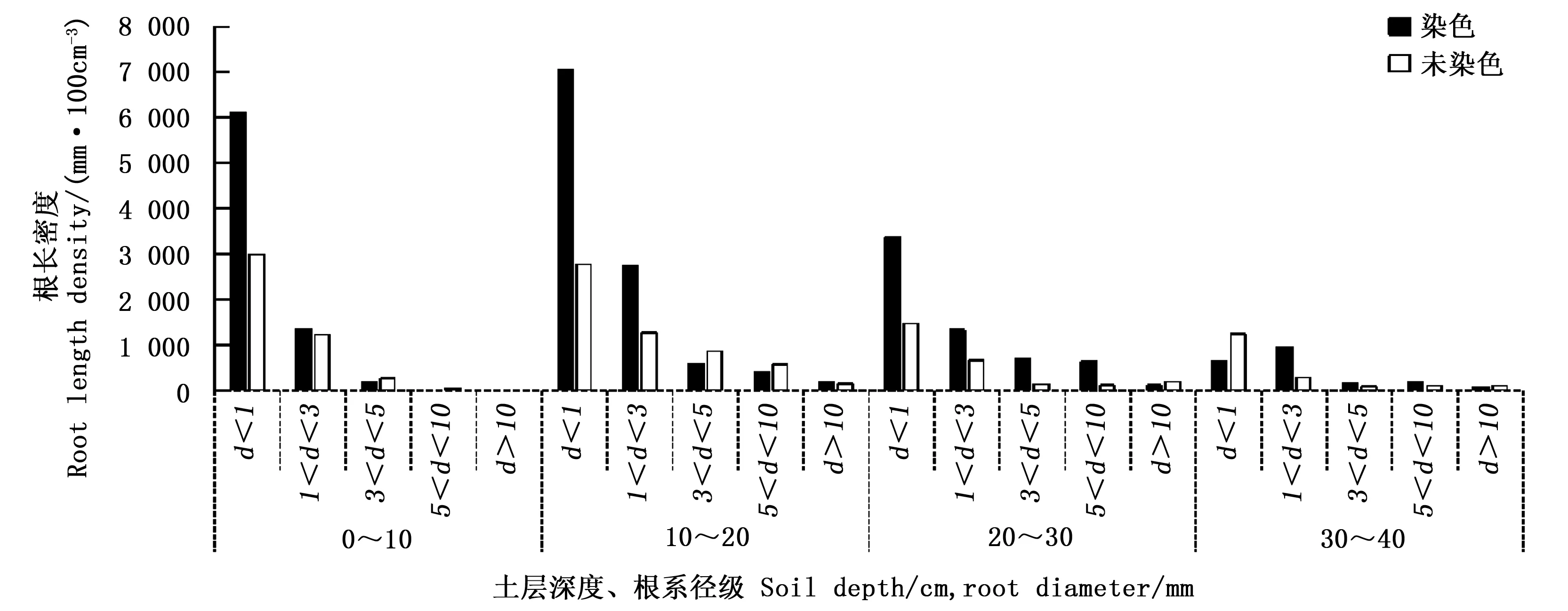
图2 不同土壤深度下根长密度在优先流区和基质流区的分布Fig.2 Distribution of root length density in preferential pathways and soil matrix for different soil depth
根系生长是一个动态的随机性的过程[20],研究点中不同土层内的水分和养分的分布,以及土壤内异质性使得根系在水平与垂直的空间分布存在明显的差异,同时活根系与死根系的相互作用,在图1中均匀染色区有显示。都导致了整个剖面内根长密度出现了较大的差异。
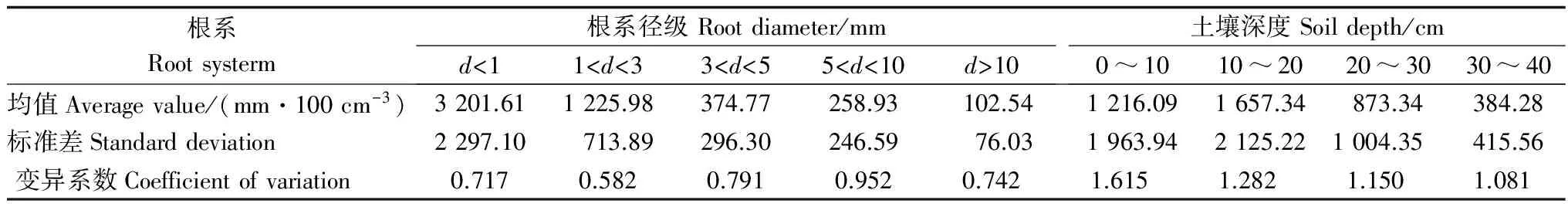
表1 优先流根长密度的均值、标准差随根系径级和土壤深度的变化
榛子灌木林优先流区根长密度基本都大于基质流区,细根系(d<1 mm)对优先流贡献率最大(67.9%),且5个不同根系径级贡献率均值都在58.5%以上,对优先流的形成为促进作用。这是因为灌木内土层土壤有一层致密的枯枝落叶层。这层枯枝落叶层内部与根系营造了许多缺氧状态下的小室,给许多厌氧动物与微生物提供了生存环境,同时也加速了死根系的腐败,使得原来紧实的土壤变的更加松动,土壤结构发生变化,转变为有机物质,加速了土壤团聚体的形成。进而改善了土壤自身理化性质[21],与根系结构(活根与死根)构成了相互连通的土壤空间网络,促进了土壤水分与溶质的快速运移。
无论根系径级的不同还是土壤层次的不同,其根长密度有显著的差异,优先流区的根系含量也相对较高,进而得出染色面积百分比与根长密度的关系,图3。
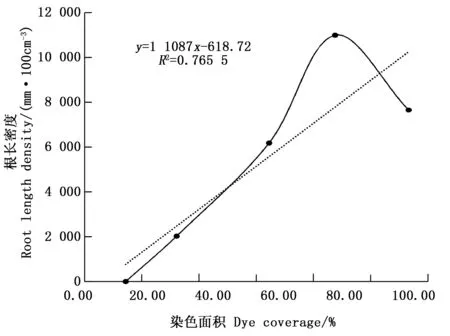
图3 染色面积百分比与根长密度的关系Fig.3 Relationship between dye coverage and root length density
图3为实验剖面内不同根系径级根长密度之和与平均染色面积百分比。随着根长密度的增加,到达根长密度最大值时(10 962.11 mm·100 cm-3),染色面积百分比也随之增加,而后根长密度降低,染色面积百分比依然增加。这是因为榛子灌木林的根系生长主要集中在土壤表层,致密的枯枝落叶层内土壤团聚体的含量较高,根系与土壤接触,容易在平面内扩散伸长和垂直穿插生长,增加了土壤孔隙的密度,增强了根隙孔道的连通性[22]。且主根系的伸长穿透力强,易于向下生长,形成较大孔道,改善土壤性质,使土壤松弛度提高,造成了许多大孔隙,利于细根在大孔隙内部的扩散和伸长,在相互作用和共同影响下,为水分和溶质的运移创造了良好的条件[23]。在土壤下层中土壤紧实,密度高,微生物与动物少,并且夹杂许多大的未风化的石砾,阻碍了根系生长,致使水分无法继续下渗,对优先流的形成起反作用。
2.3 根生物量对优先流的影响
根生物量是评价根系吸收养分和水分的能力[24-26],即为土壤单位体积内的根系的质量[28],对研究点不同根系径级的根系生物量进行分析,见图4。
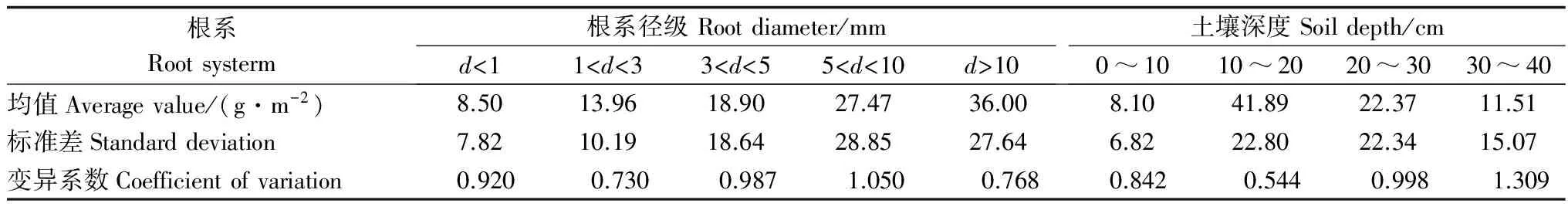
表2 优先流和基质流根生物量的变异特征
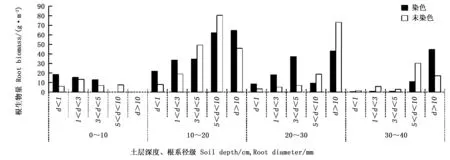
图4 不同土层根系生物量在优先流区和基质流区的变化Fig.4 Change of root biomass in preferential flow and soil matrix for different soil depth
仅仅得出优先流区的根生物量比基质流区的根生物量整体高出1%左右,并不能清楚的表明根生物量对优先流的作用是正相关还是负相关。因此用优先流区与基质流区根生物量的差值与优先流区根生物量之比(根生物量差值百分比)[27]来进行分析,见图5。
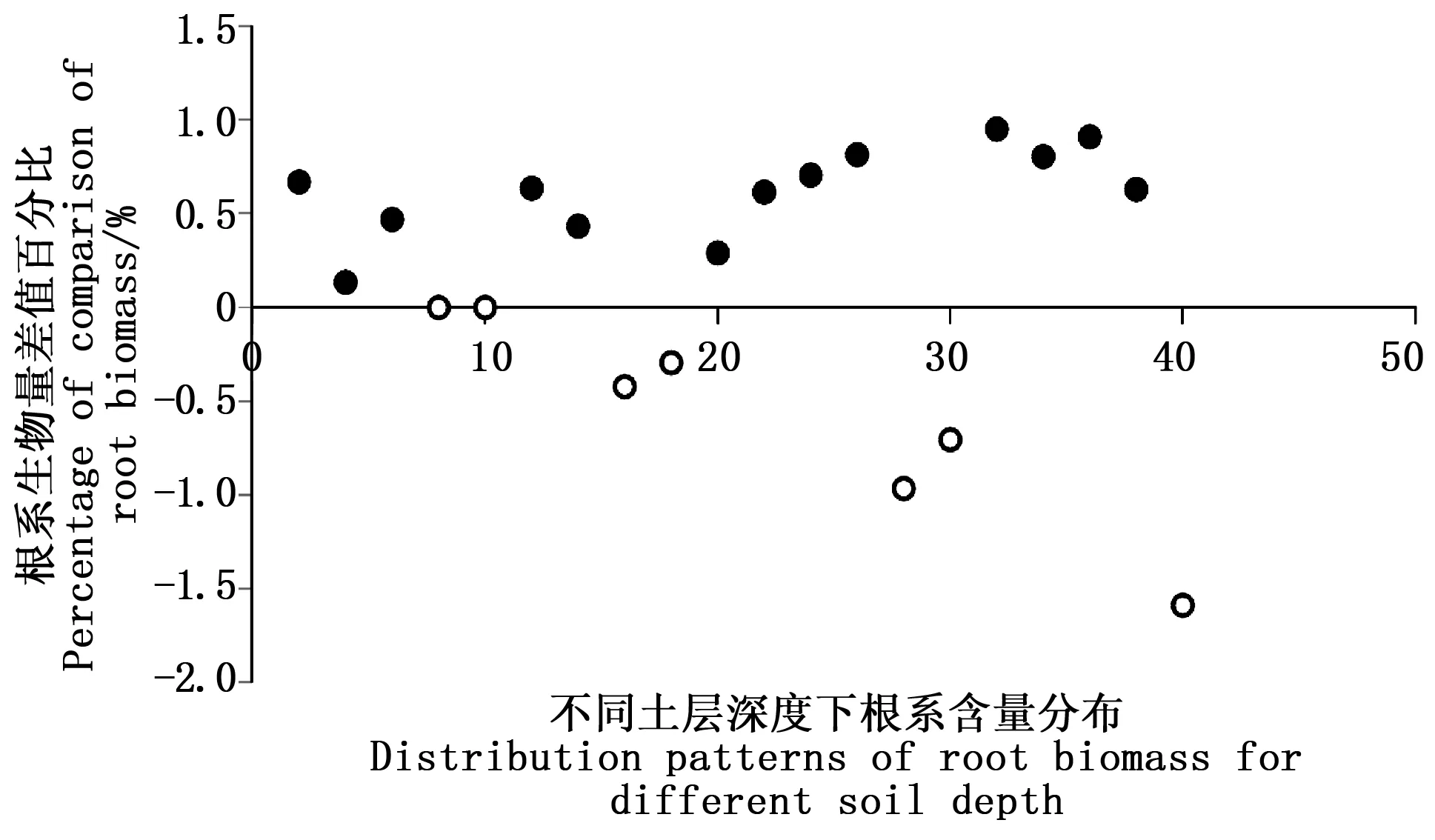
注:黑点表示优先流区和基质流区的比值为正值,白点表示两区的比值为负值。Note: the black spot indicates that the ratio of the preferential flow zone and the matrix flow area is positive, and the white spot indicates that the ratio of the two zones is negative.图5 优先流区和基质流区的根生物量差值百分比Fig.5 Different between percentages of root biomass in preferential flow and soil matrix
图5表明:当两区域差值百分比为正时,染色区的根生物量应该较大,处于主导地位,促进优先流路径的产生,当两区域的差值比为负时,未染色区的根生物量较大,处于主导地位,阻碍了优先流现象的发生,但图示中的数据信息却与现实的优先流图像提取信息不相符(图2),无论优先流区根生物量大,还是基质流区根生物量大,都有优先流路径的产生,所以根生物量对于优先流现象的产生不起决定性的作用。
3 讨论
根系是形成土壤团聚体和结构孔隙度的一个关键因素[28]。研究发现,林木根系主要集中在土壤的表层,能够改善土壤理化性质,降低土壤密度[29]。根在自然生长情况下,与根系接触的土壤会受到膨胀和挤压,使土壤孔隙变大,密度减小,而当根腐烂时,产生孔隙的几率较高,甚至会有明显的土壤管留在土壤中[30]。利于新的根在孔道中继续生长,给优先流路径提供了发生条件。生长的根与腐烂的根形成一个具有不同孔隙率的非常复杂的网络,森林土壤中的根系与石砾相互连接处有土粒相连,能形成连续的流径,根长密度随土层深度增加而降低,根长密度能反映出根系生长情况,是优先流发生的贡献者[31]。本研究表明优先流区和基质流区根长密度和根生物量都是随土壤深度的增加而减小,且优先流区要高于基质流区,这与上述的研究结果一致。
在众多研究优先流的实验方法中,野外染色剂示踪法最为直观[32],可以直接观察到优先流的现象和优先流路径的走向和数量。本研究区域是黑龙江省东部重要的水源涵养区,生态系统复杂多样化,有明显的优先流现象发生,优先流路径表现得也极为明显。Hagedoen和Bundt[33]认为,理论上优先流路径应保持稳定至下层土壤,事实上在土壤表层的土壤水分流动系统中,沿根隙孔道的优先流占主导地位,流至土壤中,根密度变小,土壤水分均匀渗透到土壤层中,产生了多条优先流路径。M Ghestem[34]研究表明在土壤水分的运移活动中,94.7%的水流是通过根系孔道运移的,仅有5.3%是通过其他方式流失的。本研究结果表明,在25 cm以下土壤中出现明显的优先流路径,这与上述结论一致。且优先流区的根长密度总是大于基质流区,同时细根系(d<1 mm)的优先流贡献率最大,且不同根系径级的优先流贡献率均值都在58.5%以上,类似的结果也在其它研究中有报道[35]。
染色面积比是作为一个衡量优先流的一个特性指标,可以推测出土壤孔隙度额度数量和孔径的大小[36]。Bogner[37]认为染色面积比的可变化性是现实优先流现象的一个重要指标。本文研究中,随着土层深度的增加,染色面积百分比逐渐降低,直至变为0,根长密度也随染色面积的增加而增加。与粗根系(d>5 mm)相比,细根系(d<1 mm)与优先流的相关性更好。因为细根系密度多,与土壤接触的表面积大,吸水能力强,渗透性大[38],促进优先流现象的产生。
4 结论
[1] Mallants D, Mohanty B P, Vervoort A,etal.Spatial analysis of saturated hydraulic conductivity in a soil with macropores[J].Soil Technology,2013,10(2):115-131.
[2] 林代杰,郑子成,张锡洲,等.不同土地利用方式下土壤入渗特征及其影响因素[J].水土保持学报,2010,24(1):33-36.
[3] Czarnes S, Hallett P D, Bengough A G,etal.Root and microbialderived mucilages affect soil structure and water transport[J]. European Journal of Soil Science,2000,51(3):435-443.
[4] 王芝芳,杨亚川,赵作善,等.土壤-草本植被根系复合体抗水蚀能力的土壤力学模型[J].中国农业大学学报,1996,1(2):39-45
[5] 孔祥飞,赵雨森,辛 颖,等.黑龙江省东部山地天然次生林土壤水分的研究[J].森林工程,2009,25(04):6-9.
[6] 孔 亮,蒙宽宏,陈 宇,等.黑龙江省东部山地灌木林土壤水分动态变化[J].东北林业大学学报,2005,33(05):44-46.
[7] Flury M, Flühler H. Brilliant Blue FCF as a Dye Tracer for Solute Transport Studies—A Toxicological Overview[J].Journal of Environmental Quality,1994,23(5):1108-1112.
[8] Morris C, Mooney S J, Young S D.Sorption and desorption characteristics of the dye tracer, Brilliant Blue FCF, in sandy and clay soils[J]. Geoderma,2008,146(146):434-438.
[9] Flury M, Flühler H.Tracer Characteristics of Brilliant Blue FCF[J].Soil Sci Soc Am J,1995,59(1):22-27.
[10] Mooney S J, Morris C. A morphological approach to understanding preferential flow using image analysis with dye tracers and X-ray Computed Tomography[J].Catena, 2008, 73(2):204-211.
[11] Fransen B, Kroon H D, Berendse F. Root Morphological Plasticity and Nutrient Acquisition of Perennial Grass Species from Habitats of Different Nutrient Availability[J]. Oecologia, 1998, 115(3):351-358.
[12] 田香姣,程金花,杜士才,等.2种土地利用方式下的优先流特征[J].水土保持学报,2014,28(03):37-41.
[13] 黄 林,王 峰,周立江,等.不同森林类型根系分布与土壤性质的关系[J].生态学报,2012,32(19):6110-6119.
[14] Hagedorn F, Bundt M.The age of preferential flow paths[J]. Geoderma,2002,108(1):119-132.
[15] 蒋小金,王恩姮,陈祥伟,等.典型黑土耕地土壤优先流环绕特征[J].应用生态学报,2010,21(12):3127-3132.
[16] 王国梁,刘国彬.黄土丘陵区长芒草群落对土壤水分入渗的影响[J].水土保持学报,2009,23(3):227-231.
[17] 邵明安,杨文治,李玉山.植物根系吸收土壤水分的数学模型[J].土壤学报,1987,24(4):295-305.
[18] Volkmar K M.A comparison of minirhizotron techniques for estimating root length density in soils of different bulk densities[J].Plant & Soil,1993,157(2):239-245.
[19] Steele S J, Gower S T, Vogel J G,etal.Root mass, net primary production and turnover in aspen,jack pine and black spruce forests in Saskatchewan and Manitoba, Canada.[J].Tree Physiology,1997,17(8-9):577-587.
[20] Wells C E, Glenn D M, Eissenstat D M.Changes in the risk of fine-root mortality with age: a case study in peach,Prunuspersica(Rosaceae).[J].American Journal of Botany,2002,89(1):79-87.
[21] Zhang X Q.Fine-root Biomass, Production and Turnover of Trees in Relations to Environmental Conditions[J].Forest Research,Jan 2001,14(5):566-573.
[22] Cannavo P, Michel J C.Peat particle size effects on spatial root distribution,and changes on hydraulic and aeration properties[J].Scientia Horticulturae,2013,151(2):11-21.
[24] Xiamen, Xing X. Fine root biomass and production of Castanopsis eyrei forests in Wuyi Mountains[J]. Chinese Journal of Applied Ecology, 1998,9(4):337-340.
[25] Liao L, Chen C, Sinica A,etal. Turnover of fine roots in pure and mixedCunninghamialanceolataandMicheliamacclureiforests[J]. Chinese Journal of Applied Ecology, 1995,6(1):7-10.
[26] 毛伶俐.生态护坡中植被根系的力学分析[D].武汉理工大学,2007.
[27] 张英虎,牛健植,朱蔚利,等.森林生态系统林木根系对优先流的影响[J].生态学报,2015,35(6):1788-1797.
[28] Angers D A, Caron J.Plant-induced Changes in Soil Structure: Processes and Feedbacks[J].Biogeochemistry,1998,42(1):55-72.
[29] Bogner C, Wolf B, Schlather M,etal. Analysing flow patterns from dye tracer experiments in a forest soil using extreme value statistics[J]. Eur J Soil Sci, 2008,59(1): 103-113.
[30] Aubertin G M.Nature and extent of macropores in forest soils and their influence on subsurface water movement[A]. USDA. FOREST SERVICE RESEARCH PAPER NE.1971.
[31] Bengough A G. Water Dynamics of the Root Zone: Rhizosphere Biophysics and Its Control on Soil Hydrology[J]. Vadose Zone Journal, 2012, 11(2):460-460.
[32] Everts C J, Kanwar R S, Alexander E C,etal.Comparison of Tracer Mobilities under Laboratory and Field Conditions[J]. Journal of Environmental Quality,1989,18(4):491-498.
[33] Hagedorn F, Bundt M.The age of preferential flow paths[J].Geoderma,2002,108(1):119-132.
[34] Ghestem M,Sidle R C,Stokes A.The Influence of Plant Root Systems on Subsurface Flow: Implications for Slope Stability[J].Bioscience,2001,61(11):869-879.
[35] 董宾芳.黄土丘陵区林地植物根系与土壤优势流关系研究[D]. 昆明:西南大学,2007.
[36] 陈晓冰,张洪江,程金花,等.基于染色图像变异性分析的优先流程度定量评价[J].农业机械学报,2015,46(5):93-100.
[37] Bogner C, Borken W, Huwe B.Impact of preferential flow on soil chemistry of a podzol[J].Geoderma,2012,175(3):37-46.
[38] 刘道平,陈三雄,张金池,等.浙江安吉主要林地类型土壤渗透性[J].应用生态学报,2007,18(3):493-498.
EffectofPlantRootSystemonPreferentialFlowofHazelnutShrubinChangbaishanMountains
WANGZhao-nan1,XINYing1,ZHAOYU-sen1
(College of Forestry, Northeast Forestry University,Harbin 150040, Heilongjiang, China)
ObjectiveTo reveal the effect of root diameter classes on the occurrence of soil preferential flow in different depth of soil layer.MethodThe study area located in the hillslope with hazelnut shrub forest in Zhangguangcailing of Changbaishan Mountains .The blue dye tracing method was applied to confirm and evaluate the changes in root biomass and root length density along preferential pathways in the soil matrix.ResultThe macropore flow, funnel flow and surround flow were the main soil water flow movement patterns which occurred rapidly with surround features. The root length density decreased with the increase of soil depth. In addition, the contributions of root length densities with root diameter (d) <1 mm, 1 root length density; root biomass; preferential flow; soil matrix. 10.13275/j.cnki.lykxyj.2017.06.001 2016-12-28 国家自然科学基金(No.31300593)。 王赵男(1989—),男,在读硕士,主要从事水土保持研究。Email:zhaonan_wang@163.com * 通讯作者:辛 颖(1979—),女,博士,副教授主要从事水土保持与林业生态研究。Email:xinying2004@126.com。 S794 A 1001-1498(2017)06-0887-08 彭南轩)
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
小小说月刊·下半月(2021年8期)2021-09-05
商周刊(2018年25期)2019-01-08
传媒评论(2018年5期)2018-07-09
动漫界·幼教365(大班)(2018年3期)2018-05-14
湖南大学学报·自然科学版(2016年12期)2017-05-12
中国卫生(2016年12期)2016-11-23
江苏农业科学(2016年4期)2016-06-14
西北植物学报(2015年9期)2015-07-05
