
热带海滨植物水芫花(Pemphis acidula)的生理生态学特性
2017-12-29 07:00:44简曙光王佳新吴淑华
生态环境学报 2017年12期
曹 策,简曙光,任 海,王 俊,刘 楠*,王佳新,吴淑华
1. 广东省应用植物学重点实验室//中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049
热带海滨植物水芫花(Pemphis acidula)的生理生态学特性
曹 策1,2,简曙光1,任 海1,王 俊1,刘 楠1*,王佳新1,2,吴淑华1,2
1. 广东省应用植物学重点实验室//中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049
热带珊瑚岛生态系统处于高温、强光、台风频发、季节性干旱等恶劣环境气候条件下,极易退化,有必要进行植被恢复。水芫花(Pemphis acidula)为千屈菜科小灌木,自然分布于热带海滨和石灰质岩岸,在海岸固沙和海岛植被恢复中具有潜在的应用价值。以中国西沙群岛自然生长的水芫花为研究对象,对其叶片和茎秆形态解剖结构、生理学特征、叶片营养元素及适生土壤理化性质进行了分析研究,以期全面了解水芫花的生理生态特性及其对海滨环境的适应机理。研究结果表明,水芫花叶片厚度达到703.798 μm,栅栏组织与海绵组织的厚度比值仅为0.563,叶片高度肉质化,有利于叶片保水。叶片气孔密度低,仅为44.44 mm-2,有利于减小蒸腾。水芫花导管直径偏低(35.639 μm),导管密度为49.528 mm-2,有利于提升水分利用效率并有效避免栓塞。水芫花叶片超氧化物歧化酶活性较高(340.827 U∙g-1),总抗氧化能力高达834.016 U∙g-1,总酚含量较高(16.482 mg∙g-1),丙二醛含量仅为10.490 nmol∙g-1,说明其通过提高抗氧化能力降低膜脂过氧化伤害程度。水芫花生长环境的土壤养分含量低,但钙含量高达38.570 g∙kg-1,含水量仅为9.5%,土壤环境较差;而水芫花叶片的氮磷含量属于正常水平,说明其能够高效利用贫瘠土壤中的养分。综上,水芫花具有较强的抗干旱能力和抗胁迫能力,适宜生长于土壤较为贫瘠的热带滨海生境中,是热带珊瑚岛植被恢复的重要备选物种。
水芫花;生物学特性;海滨植物
水芫花(Pemphis acidula)又称为海芙蓉,是千屈菜科水芫花属多分枝小灌木,高约1 m,有时可长成乔木状,高可达11 m。其叶片对生,较厚,肉质,呈椭圆形、倒卵状矩圆形或线状披针,无叶柄或叶柄很短。其花二型,蒴果革质,倒卵形,种子多数,具厚翅(方文培,1983)。水芫花主要分布于东半球热带海岸,包括亚洲和大洋洲的热带地区。水芫花在中国的分布范围有限,仅分布于海南岛、西沙群岛和台湾地区(王健等,2010)。
水芫花自然生长于热带海滨或海岸地区,主要分布在含盐量及pH较高的珊瑚礁和石灰质岩岸,生长环境光照充足,往往形成单一优势群落(邢福武等,1993)。这些生长条件限制了其分布范围,因而水芫花种群处于濒危边缘(王健等,2010)。作为一种海滨植物,水芫花应归属于真红树植物还是半红树植物仍存在一定的争议(王文卿等,2007;Chapman,1984)。牟美蓉(2007)通过对33种红树植物的叶片结构性状和元素含量进行测定和比较后发现,水芫花具有真红树植物的结构特征和元素含量特征,并建议将其归为真红树植物。水芫花提取物具有一定等抗氧化性和抑菌活性,可用于生物医药,具有抗肿瘤、消炎等作用(王友绍等,2004)。此外,水芫花木材坚硬,不易劈裂,表面光滑,还可用作工具把柄。由此可见,有关水芫花的研究主要集中在形态和分类学特征以及其药用价值的发掘利用方面,尚缺乏对热带海岛原生条件下水芫花生理生态学特征的系统性研究。
中国是海洋大国,有着众多岛屿。海岛作为海洋生态系统的重要组成部分,其资源开发利用关系到国家的可持续发展,受到国际政府和学术界的高度重视(杨红等,2012)。相对于陆地生态系统,海岛大多面积狭小、环境封闭、生态系统极易退化。为此,2017年颁布的《全国海岛保护工作“十三五”规划》将“强化海岛生态空间保护、保护海岛生物多样性、修复海岛生态系统、推动社区共建共享”作为主要工作任务。热带珊瑚岛礁是海洋中一类特殊的岛屿生态系统,高温、强光、台风、季节性干旱、土壤盐碱化、贫瘠等环境特点导致其频繁受到干扰,植被恢复是保护和改善珊瑚岛礁的重要环节,而选取适生植物又是珊瑚岛礁植被恢复工作的重中之重(李婕等,2016;罗琦等,2017)。水芫花作为热带海滨植物,其在热带珊瑚岛植被恢复中具有极大的潜力和应用价值。因此,本文以西沙群岛东岛的野生水芫花为研究对象,测定其叶片的形态解剖学特征、生理学特征、主要元素含量特征及适生土壤理化性质,探究其生态适应性,为水芫花的广泛利用提供参考。
1 材料与方法
1.1 研究地概况
研究地点位于西沙群岛东岛,东经 112°43′13″~112°44′22″,北纬 16°39′34″~16°40′31″。东岛是一个珊瑚岛,面积约1.7 km2,属于热带海洋气候,年平均气温26~27 ℃,年降水量约1500 mm,年蒸发量约 2400 mm,降水集中于 6—11月,占全年的87%,12—5月为干季,年平均湿度为 81%(曹垒等,2003),岛上植物受季节性干旱影响明显。东岛是西沙群岛自然植被保存最好的岛屿,植物种类丰富,优势种主要有白避霜花(Ceodes grandis)、草海桐(Scaevola sericea)、海岸桐(Guettarda speciosa)、银毛树(Messerschmidia argentea),其中水芫花成片生长在岛屿的东北海边(童毅等,2013)。
1.2 研究方法
于2016年6月在水芫花单一优势群落中选取5株健康的野生水芫花成年植株(年龄约 20 a),采集其生长发育良好的叶片和茎秆,分别装袋,并置于船载冰箱中,上岸后用冰盒带回实验室进行各项分析。同时,采集水芫花根部周围土壤(0~20 cm),用于测定分析水芫花适生土壤的主要理化性质。
1.2.1 形态解剖特征
1.2.1.1 叶片解剖结构
选取健康成熟的叶片,以徒手法制作叶片切片,并在Leica DM 4000B显微镜下拍摄,用Image-J软件分析显微图片。在图片中测量叶片厚度、栅栏组织和海绵组织厚度。用纸巾擦净叶背面中部后涂一层透明指甲油,用镊子轻轻将风干的指甲油膜揭下,平铺于载玻片上,盖上盖玻片后于光学显微镜下观察、拍照,每个叶片拍摄10个视野。用Image-J软件处理照片并测量保卫细胞的长度,即气孔长度(SL);计算视野中单位叶面积的气孔数量,即气孔密度(SD)。计算气孔面积指数(SPI),SPI=SL2×SD(Sack et al.,2003)。用LI-3000A叶面积仪测量叶面积,烘干称质量,计算比叶面积(SLA),比叶面积为叶面积与叶干质量之比。
1.2.1.2 茎秆解剖结构
徒手切片法制作茎秆横切面切片。利用 Leica滑走切片机(SM2010R)获取边材横切片(厚度30~50 μm),利用 Leica显微成像系统(DM2500)扫描并拍照,运用Image J软件分析导管直径、导管密度(李荣等,2016)。
1.2.2 叶片生理学特征
1.2.2.1 抗氧化酶活性
过氧化物酶(POD)测定:采用愈创木酚显色法进行测定,POD催化H2O2氧化特定底物,在470 nm处有特征光吸收,以1 min内,1 g组织在1 mL反应体系中A470变化0.5个单位为1个酶活力单位(U∙g-1)(王琰等,2011)。
超氧化物歧化酶(SOD)测定:采用氮蓝四唑还原法进行测定,通过黄嘌呤及黄嘌呤氧化酶反应系统产生超氧阴离子(O2-),O2-可还原氮蓝四唑生成蓝色甲臜,后者在560 nm处有光吸收,以1 g组织抑制氮蓝四唑还原为50%时定义1个酶还原单位(U∙g-1。邓茳明等,2010)。
过氧化氢酶(CAT)测定:采用紫外吸收法进行测定,H2O2在240 nm下有特征吸收峰,CAT能够分解H2O2,使反应溶液在240 nm处的吸光度随反应时间的延长而下降,根据吸光度的变化率可计算出CAT活性,以1 g组织每分钟催化1 nmol H2O2的降解量为一个酶活力单位(U∙g-1。Aebi,1984)。
1.2.2.2 总酚含量和总抗氧化能力
采用福林酚比色法测定,在碱性条件下,酚类物质将钨钼酸还原,产生蓝色化合物,其在760 nm处有特征吸收峰,测定760 nm处的吸光值,即可得样品总酚含量(Singleton et al.,1999)。
采用FRAP法测定叶片的总抗氧化能力,在酸性条件下,Fe3+-TPTZ(三吡啶三吖嗪)可被样品中还原物质还原为蓝色的Fe2+-TPTZ,其在593 nm处具有最大光吸收,在Fe3+-TPTZ过量时,检测蓝色物质的生成量可反映待测样品的总抗氧化能力(Benzie et al.,1996)。
1.2.2.3 脯氨酸(Pro)测定
采用茚三酮比色法测定,用磺基水杨酸提取脯氨酸,加热处理后,脯氨酸与酸性茚三酮溶液反应生成红色沉淀;加入甲苯萃取后,在520 nm处测定吸光度(Bates et al.,1973)。
1.2.2.4 脱落酸(ABA)测定
采用高效液相色谱法测定,取0.1 g叶片,研磨后提取液上高效液相色谱仪,于254 nm处测定样品吸光度,再换算成ABA含量(孙崇臻等,2013)。
1.2.2.5 丙二醛(MDA)测定
采用硫代巴比妥酸法对MDA含量进行测定,分布读取其在532 nm和600 nm处的吸光值,利用其差值计算MDA的含量(孙群等,2006)。
1.3 叶片及土壤元素含量特征
1.3.1 叶片养分元素测定
将挑选完的剩余叶片置于 65 ℃下烘干至恒重后,磨碎过100目筛,称取0.1 g样品酸化后采用靛酚蓝比色法(Cornelissen et al.,2003)测定氮元素含量;称取0.1 g样品采用丘林法(刘伟等,2010)测定总碳含量;称取0.1 g样品采用钼锑抗比色法(刘清等,2007)测定磷元素含量。
1.3.2 土壤理化性质测定
将植物的适生土壤(0~20 cm)充分混合,用镊子剔除植物残体、根系和岩屑等异物,过2 mm孔径(10目)筛。于室温下自然风干,按四分法取30~50 g土样,去除细根后研细过100目筛,加入过量1 mol·L-1HCL于室温下处理24 h,以充分去除碳酸盐,再用蒸馏水反复洗涤至洗涤液呈中性,过滤后于65 ℃下烘至恒重。利用pH 211酸度离子测定仪测定 pH值;采用靛酚蓝比色法(Cornelissen et al.,2003)测定氮元素含量,丘林法(刘伟等,2010)测定总碳含量,钼锑抗比色法(刘清等,2007)测定磷元素含量;利用原子吸收光谱仪测定其他元素含量。
1.4 数据处理
将测定的各项指标数据录入 Microsoft Excel 2013中计算平均值和标准误;运用Adobe Photoshop 5对叶片解剖结构图进行标记。
2 结果与分析
2.1 叶片和茎秆形态解剖特征
如表1和图1所示,水芫花叶片主要由表皮、栅栏组织、海绵组织3部分组成。叶片表皮排列紧密,上表皮外有角质层,且有明显的纤毛分布。叶片高度肉质化,其中栅栏组织厚193.548 μm,海绵组织厚 344.456 μm,叶片总厚度为 703.798 μm,栅栏组织与海绵组织的厚度比值为 0.563,栅栏组织呈条状排列紧密,海绵组织呈散乱排列。气孔密度为44.44 mm-2,气孔指数值为0.085,气孔保卫细胞长为43.324 μm。水芫花茎秆中的导管密度达到49.528 mm2,导管直径偏小,为35.639 μm。
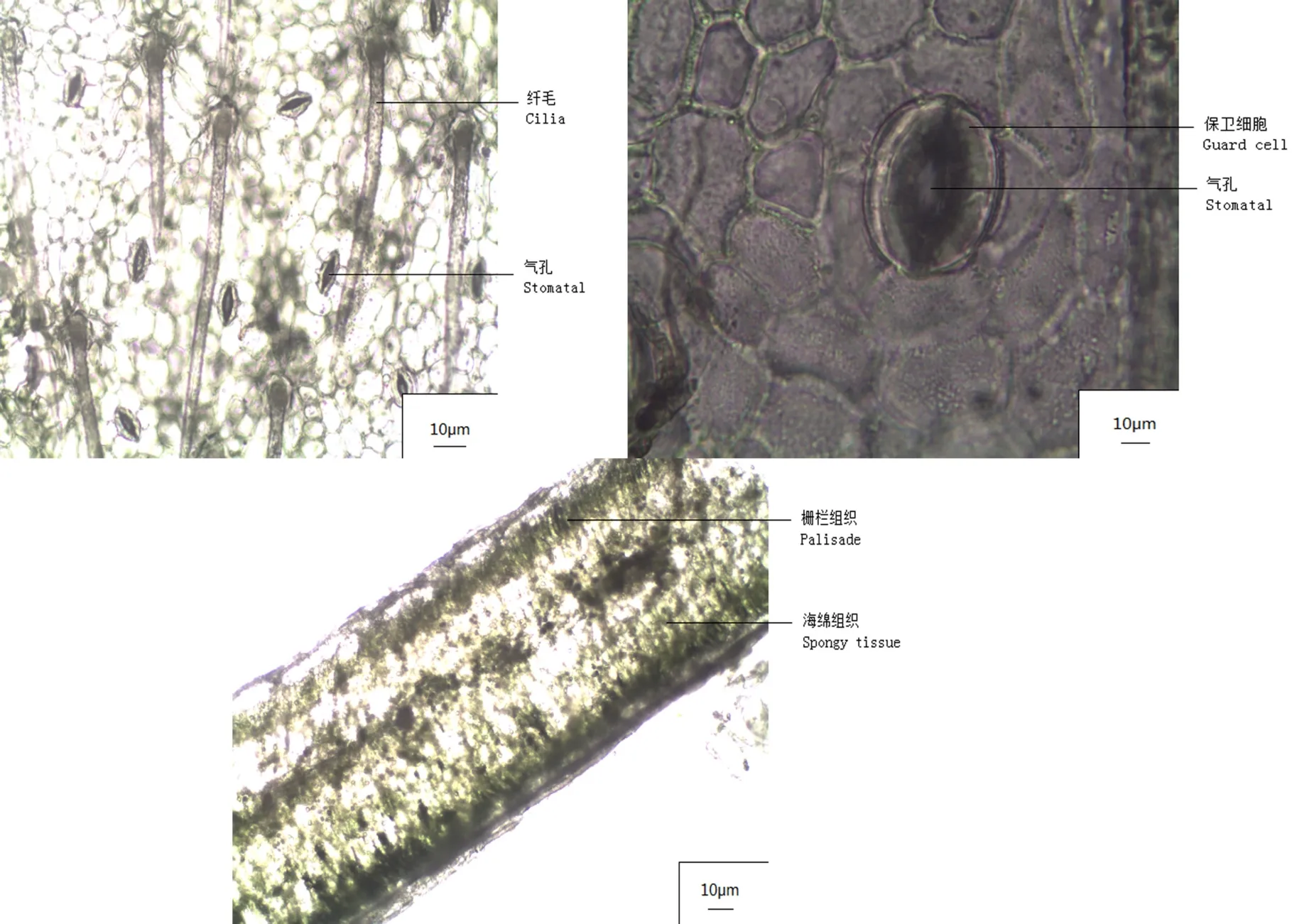
图1 水芫花叶片的解剖结构Fig.1 Anatomical structure of Pemphis acidula
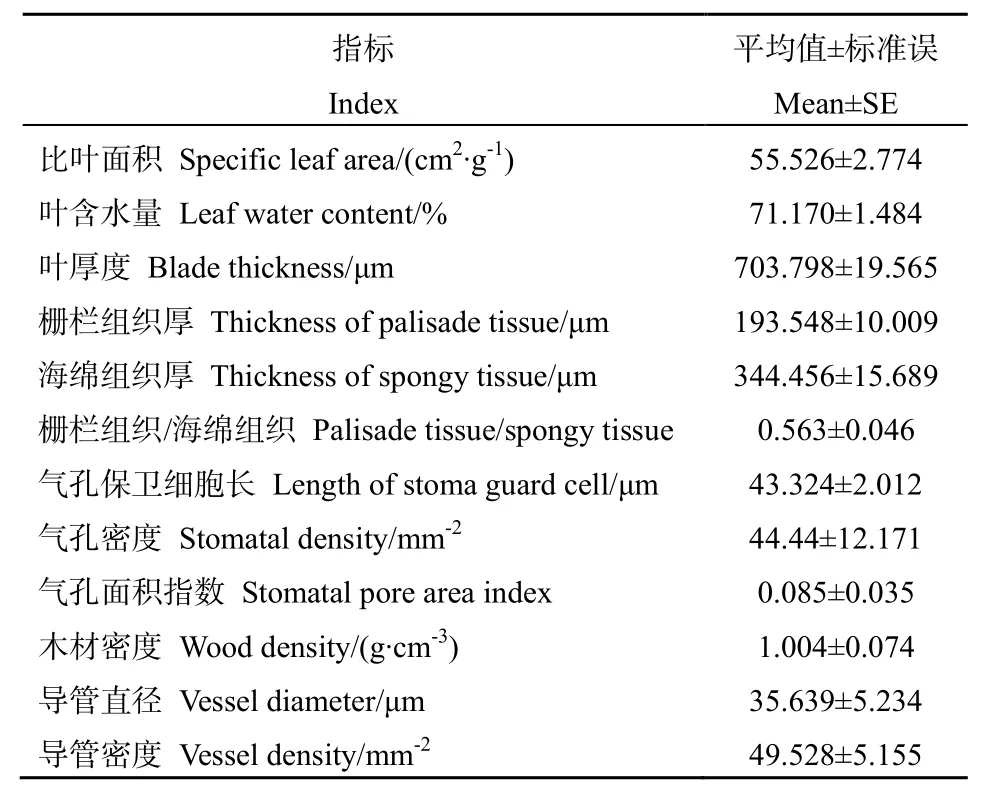
表1 水芫花形态解剖特征Table1 Morphological characteristics of Pemphis acidula
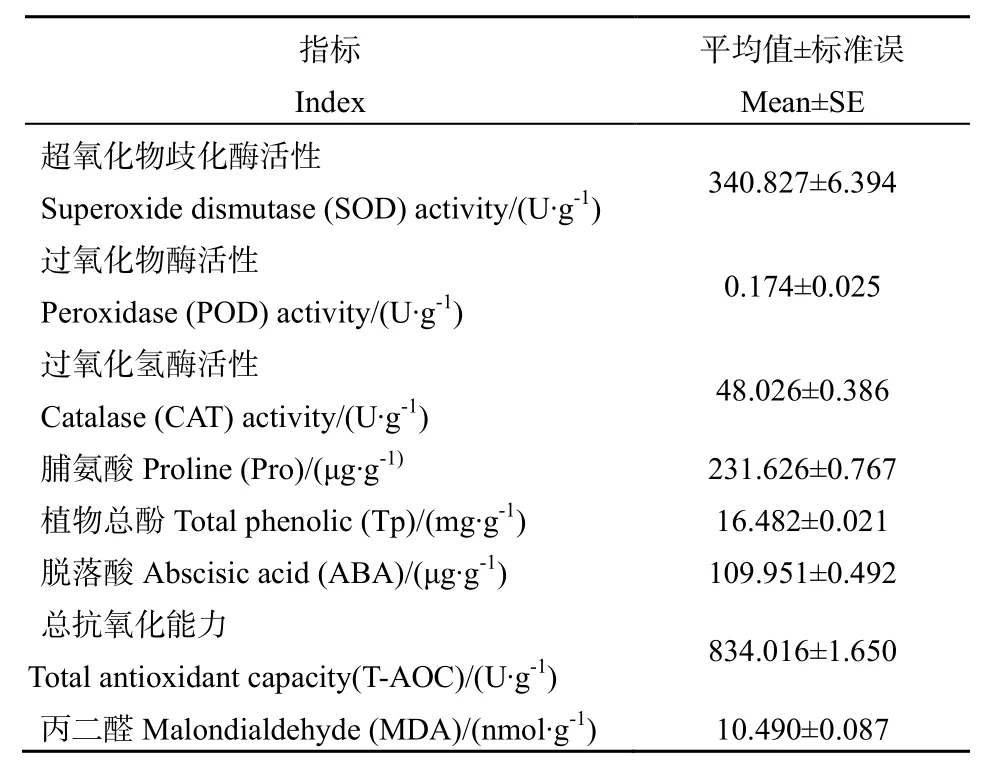
表2 水芫花生理学特征Table2 Physiological characteristics of Pemphis acidula
2.2 叶片生理学特征
水芫花的主要抗逆生理特征属性如表2所示。叶片抗氧化酶中,SOD活性较高,达到 340.827 U∙g-1,POD 和 CAT 活性分别为 0.174、48.026 U∙g-1,总抗氧化能力高达834.016 U∙g-1,表明叶片具有高效的清除和转移活性氧的能力。水芫花叶片中主要抗氧化物质总酚含量为16.482 mg∙g-1,与叶片抗逆生理相关的Pro、ABA含量分别为231.626、109.951 μg∙g-1。叶片中膜脂过氧化产物MDA含量较低,仅为10.490 nmol∙g-1,表明水芫花受到的胁迫不足以导致体内的膜脂系统受到活性氧胁迫,不能引起MDA的积累。
2.3 营养元素及土壤理化性质
水芫花叶片营养元素含量如表3所示。叶片各元素中总碳含量最高,达到358.416 g∙kg-1;其次依次为总氮含量和总磷含量,分别为 5.656 g∙kg-1和2.033 g∙kg-1。水芫花叶片碳氮比为63.369,碳磷比为 176.299。如表 4所示,水芫花适生土壤理化性质中,土壤含水量较低,仅为9.5%,pH值为7.75,呈弱碱性。土壤元素含量中,钙含量最高,达到38.570 g∙kg-1,显示出石灰质土壤基质的主要特征。土壤有机碳含量为 5.519%,镁含量达到 4.34 0g∙kg-1,铁含量为 1.121 g∙kg-1,其余金属元素(钾、钠、锰、锌、铜)含量均甚微,低于0.413 g∙kg-1。

表3 叶片营养元素含量Table3 Leaf nutrient content of Pemphis acidula
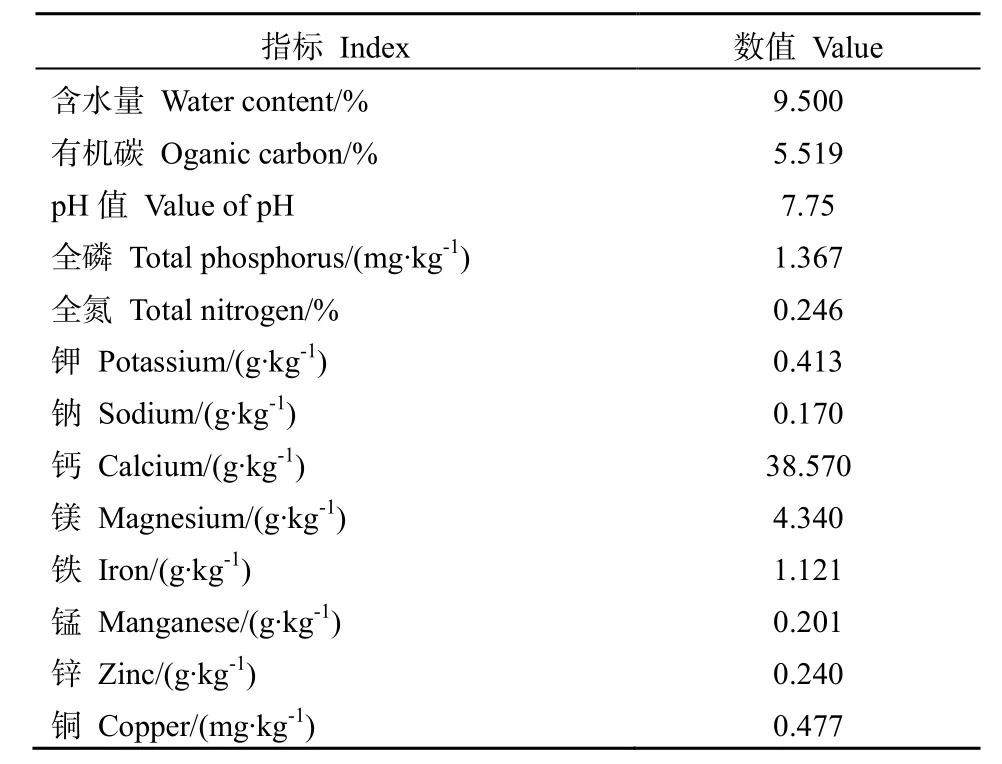
表4 土壤营养元素含量Table4 Soil nutrient content of Pemphis acidula
3 讨论
3.1 水芫花叶片形态解剖特征
比叶面积是指单位干重的鲜叶面积,与植物的耐旱性及光合作用的能力相关(Meziane et al.,1999)。比叶面积是重要的叶片形态解剖特征,较小的比叶面积可以一定程度上防止干旱环境中植物水分的丧失(Costa-Saura et al.,2016)。比叶面积可以反映植物获取资源的能力,干旱环境下的植物比叶面积较低,而具有高比叶面积的植物对体内营养物质的保存能力较强(Lambers et al.,1992)。李玉霖等(2005)研究发现,沙丘生境中5种植物的比叶面积的平均值在89.0~246.0 cm2∙g-1,而生长于西沙群岛东岛的水芫花比叶面积仅为 55.526 cm2∙g-1,远低于沙丘生境植物,说明水芫花可以更好地适应干旱环境。
潘昕等(2015)对青藏高原25种灌木叶片的结构特征进行研究,发现这些植物叶片的平均厚度为186.25 μm,最高可达323.00 μm,并指出叶片厚度是划分植物抗旱性的重要形态学指标之一。与此相比,水芫花叶片厚度为 703.798 μm,远高于干旱半干旱地区的灌木,说明水芫花对干旱条件的适应性较强。同时,水芫花的栅栏组织厚度为193.548 μm,海绵组织厚度为344.456 μm,栅栏组织与海绵组织的厚度比值为0.563,叶片呈高度肉质化的特征,其叶片较厚,海绵组织较厚,栅栏与海绵组织的厚度比值较低,有利于水分在叶片中的存储,提高了其耐旱性。
气孔是控制植物水分运输的重要通道,气孔密度反映了植物的耐旱能力(潘昕等,2015)。张义等(2014)对 11种城市森林植物的研究发现其气孔密度在80.6~408.4 n∙mm-2之间。水芫花气孔密度仅为44.44 n∙mm-2,气孔面积指数也较低,这在一定程度上减弱了其蒸腾作用,降低了植物体内水分的散失,维持了其较高的耐旱能力。
一般导管直径越大,导管密度越高,植物运输水分的效率越高(Bass,1982),但导管直径越大越易发生栓塞导致耐旱性降低(Cai et al.,2010)。李荣等(2016)对6种耐旱植物木质部结构进行研究,发现6种耐旱植物的导管直径在22.991~58.873 μm之间,导管密度在29.632~284.385 mm2之间。水芫花的导管直径为 35.639 μm,导管密度为 49.528 mm2,处于耐旱植物的范围内,同时其导管直径偏低,但其导管密度处于中等水平。这些特征说明水芫花通过较多的导管在提升运输水分效率的同时,减小导管直径以降低栓塞风险,在干旱环境下可以保持体内的水分平衡。
3.2 水芫花叶片生理学特征
MDA是膜脂过氧化的产物,也是表征氧化胁迫导致的膜脂过氧化最常用的的重要指标(龚吉蕊等,2004)。龚吉蕊等(2004)对干旱胁迫下几种荒漠植物抗氧化能力的比较研究发现,干旱胁迫下3种植物的MDA都表现出增加的趋势,其含量为45~70 μmol∙g-1。水芫花内的 MDA 含量仅为 10.490 nmol∙g-1,低于受到干旱胁迫后 3种荒漠植物的MDA水平,说明西沙群岛自然生长的水芫花的抗氧化能力较强,因而膜脂过氧化水平较低。
植物在受到环境胁迫时会大量合成 Pro和ABA,以增强对环境胁迫的抵抗能力(刘红娟等,2008)。Pro是表征植物耐旱性的指标之一,脱落酸具有加速叶片果实脱落的作用,同时可以促进叶片气孔关闭。水芫花Pro和ABA的含量较高,分别为 231.626 μg∙g-1、109.951 μg∙g-1,说明其抵御环境胁迫引起的氧化伤害的能力较高。
3.3 水芫花叶片营养元素和适生土壤理化性质
对水芫花适生土壤养分含量进行分析,结果显示,其土壤含水量为9.5%,pH值为7.75,呈弱碱性,有机碳含量为 5.519%,全氮含量偏低,仅为0.246%,全磷含量为1.367 mg∙kg-1。由于东岛属于珊瑚岛,富含碳酸钙等物质,因此其钙含量较高,可达 38.570 g∙kg-1,但总体来说,其土壤养分含量较低,土壤较为贫瘠。植物营养元素含量可以反映植株的生长状况(刘广全等,2001)。氮和磷是重要的结构组成元素,植物体内氮元素含量约在5~50 g∙kg-1之间,磷元素含量约在 1~5 g∙kg-1之间(卢琼等,2009)。通过对水芫花叶片碳、氮、磷含量的研究发现,水芫花叶片中氮、磷含量分别为 5.656 g∙kg-1,2.033 g∙kg-1,属于正常范围。水芫花生长在较为贫瘠的土壤中,但其生长状况良好,对氮、磷等重要元素的利用满足了其自身生长的需要,其对养分的利用能力较高。
4 结论
本文对水芫花形态解剖学特征、生理特征及营养元素含量及利用能力进行了综合研究,结果表明水芫花是一种阳生性植物,对干旱环境的耐受能力高,对环境逆境引起的氧化胁迫具有较高的抵御能力,对土壤养分的吸收利用能力强。因此,水芫花适宜生长在高温、高光和季节性干旱显著、土壤保水能力差、养分含量低的热带岩石暗礁及珊瑚岛礁上。
水芫花具有适应热带海滨环境的生理生态特性,可作为热带珊瑚岛植被恢复、海滩固沙的重要备选物种。
ABELES Y F, LIANG Y C, LOU Y S, et al. 2003. Influences of silicon on peroxide dismutase activity and lignin content of wheat (Tritium aestivum L.) and its relation to resistance to powdery mildew [J].Scientia Agricultura Sinica, 36(7): 813-817.
AEBI H. 1984. Catalase in vitro [J]. Methods in Enzymology, 105(1):121-126.
BASS P. 1982. New perspectives in wood anatomy [M]. Forestry Sciences,Dordrecht: Springer Science.
BATES L S, WALDREN R P, TEARE I D. 1973. Rapid determination of free proline for water stress studies [J]. Plant and Soil, 39(1): 205-207.
BENZIE I F F, STRAIN J J. 1996. The ferric reducing ability of plasma(FRAP) as a measure of “antioxidant power”: the FRAP assay [J].Analytical biochemistry, 239(1): 70-76.
CAI J, TYREE M T. 2010. The impact of vessel size on vulnerability curves: data and models for within-species variability in saplings of aspen, Populus tremuloides Michx [J]. Plant, Cell & Environment,33(7): 1059-1069.
CHAPMAN V J. 1984. Botanical surveys in mangrove communities[M]//SNEDAKER S C, SNEDAKER J G. The mangrove ecosystem:research methods. Paris: UNESCO: 53-80.
CORNELISSEN J H C, LAVOREL S. GARNIER E, et al. 2003. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide [J]. Australian Journal of Botany, 51(4):335-380.
COSTA-SAURA J M, MARTÍNEZ-VILALTA J, TRABUCCO A, et al.2016. Specific leaf area and hydraulic traits explain niche segregation along an aridity gradient in Mediterranean woody species [J]. Perspectives in Plant Ecology Evolution & Systematics,21(1): 23-30.
LAMBERS H, POORTER H. 1992. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences [J]. Advances in Ecological Research, 23(6): 188-242.
SINGLETON V L, ORTHOFER R, LAMUELA-RAVENTÓS R M. 1999.Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent [J]. Methods in Enzymology. 299 (1): 152-178.
MEZIANE D, SHIPLEY B. 1999. Interacting determinants of specific leaf area in 22 herbaceous species: effects of irradiance and nutrient availability [J]. Plant, Cell & Environment, 22(5): 447-459.
SACK L, COWAN P D, JAIKUMAR N, et al. 2003. The ‘hydrology’ of leaves: co-ordination of structure and function in temperate woody species [J]. Plant Cell & Environment, 26(8): 1343-1356.
WILLEKENS H, INZÉ D, MONTAGU M V, CAMP W V. 1995. Catalases in plants [J]. Molecular Breeding, 1(3): 207-228.
曹垒, 张苏芳, 史洪泉, 等. 2003. 西沙群岛东岛小军舰鸟繁殖种群的初步观察[J]. 动物学研究, 24(6): 457-461.
邓茳明, 熊格生, 袁小玲, 等. 2010. 棉花不同耐高温品系的SOD、POD、CAT活性和MDA含量差异及其对盛花期高温胁迫的响应[J]. 棉花学报, 22(3): 242-247.
方文培. 1983.中国植物志[M]. 第53卷. 北京: 科学出版社: 89-90.
龚吉蕊, 赵爱芬, 张立新, 等. 2004. 干旱胁迫下几种荒漠植物抗氧化能力的比较研究[J]. 西北植物学报, 24(9): 1570-1577.
李婕, 刘楠, 任海, 等. 2016. 7种植物对热带珊瑚岛环境的生态适应性[J]. 生态环境学报, 25(5): 790-794.
李荣, 党维, 蔡靖, 等. 2016. 6个耐旱树种木质部结构与栓塞脆弱性的关系[J]. 植物生态学报, 40(3): 255-263.
李玉霖, 崔建垣, 苏永中. 2005. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较[J]. 生态学报, 25(2): 305-310.
刘广全, 赵士洞, 王耗, 等. 2001. 锐齿栎林个体光合器官生长与营养季节动态[J]. 生态学报, 21(6): 883-889.
刘红娟, 刘洋, 刘琳. 2008. 脱落酸对植物抗逆性影响的研究进展[J]. 生物技术通报, (6): 7-9.
刘清, 李玉, 姚惠源. 2007. Folin-Ciocalteu 比色法测定大麦提取液中总多酚的含量[J]. 食品科技, 4(3): 175-177.
刘伟, 龙瑞军, 王长庭, 等. 2010. 高寒草甸不同群落类型土壤碳分布与物种多样性、生物量关系[J]. 资源科学, 32(10): 2020-2027.
罗琦, 刘慧, 吴桂林, 等. 2017. 基于功能性状评价5种植物对热带珊瑚岛环境的适应性[J]. 生态学报, DOI: 10.5864/stxb201612152597.
卢琼, 王俊, 张倩媚, 等. 2009. 白花油麻藤(Mucuna birdwoodiana)的生态生物学特征[J]. 生态科学, 28(3): 213-216.
牟美蓉. 2007. 基于叶片特征的真红树和半红树植物的比较研究[D]. 厦门: 厦门大学.
潘昕, 邱权, 李吉跃, 等. 2015. 基于叶片解剖结构对青藏高原 25种灌木的抗旱性评价[J]. 华南农业大学学报, 36(2): 61-68.
孙崇臻, 王超, 蔡子哲, 等. 2013. 高效液相色谱测定蜂蜜中的脱落酸、黄酮和酚酸[J] 食品科学, 34(10): 281-285.
孙群, 胡景江. 2006. 植物生理学研究技术[M]. 咸阳: 西北农林科技大学出版社.
童毅, 简曙光, 陈权, 等. 2013. 中国西沙群岛植物多样性[J]. 生物多样性, 21(3): 364-374.
王健, 龚萍, 赵钟鑫, 等. 2010. 水芫花研究进展[J]. 安徽农业科学,38(20): 10729-10730.
王琰, 陈建文, 狄晓艳. 2011. 水分胁迫下不同油松种源SOD、POD、MDA及可溶性蛋白比较研究[J]. 生态环境学报, 20(10):1449-1453.
王文卿,王瑁. 2007. 中国红树林[M]. 北京:科学出版社.
王友绍, 何磊, 王清吉, 等. 2004. 药用红树植物的化学成分及其药理研究进展[J]. 中国海洋药物, 23(2): 26-31.
邢福武, 李泽贤. 1993. 我国西沙群岛植物区系地理的研究[J]. 热带地理, 13(3): 250-257.
杨红, 苏婷, 戴小杰. 2012. 上海市无居民岛土地资源开发适宜性研究[J]. 上海海洋大学学报, 21(1): 92-96.
杨盛昌, 谢潮添, 张平, 等. 2003. 低温胁迫下弓葵幼苗膜脂过氧化及保护酶活性的变化[J]. 园艺学报, 30(1): 104-106.
张玲玲, 韩诗畴, 李志刚, 等. 2006. 艳婀珍蝶取食对薇甘菊叶片生理指标的影响[J]. 生态学报, 26(5): 1330-1336.
张义, 王得祥, 宋彬, 等. 2014. 基于叶片解剖结构的西宁市 11种城市森林植物抗旱性评价[J]. 西北农林科技大学学报(自然科学版),42(8): 87-92.
赵保路. 1999. 氧自由基和天然抗氧化剂[M]. 北京: 科学出版社: 54-59.
The Ecophysiological Characteristics of Pemphis acidula, A Tropical Beach Plant
CAO Ce1,2, JIAN Shuguang1, REN Hai1, WANG Jun1, LIU Nan1*, WANG Jiaxin1,2, WU Shuhua1,2
1. Guangdong Provincial Key Laboratory of Applied Botany//South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 2. University of Chinese Academy of Sciences, Beijing 100049, China
The tropical coral island ecosystem is easy to degenerate under the harsh environment such as high temperature, strong sunlight, frequent typhoon, seasonal drought, so the vegetation restoration is necessary. Pemphis acidula (lythraceae) is a small shrub, mainly distributes in tropical coastal beach and calcareous rocky shores, with potential application value in costal sand fixation and island revegetation. In this study, leaf and branch morphological and anatomical structures, physiological characteristics, leaf nutrient elements and the suitable soil characteristics of P. acidula habitated in Xisha islands were analyzed. The results showed that the leaf thickness of P. acidula was up to 703.798 μm, and the palisade tissue/spongy tissue ratio was only 0.563. The leaf was highly succulent, which was beneficial to water retention. The leaf stomatal density was as low as 44.44 mm-2, which was beneficial to reduce transpiration. The vessel diameter was low (35.639 μm) and the vessel density was 49.528 mm-2, which were beneficial to improve water use efficiency and effectively avoid embolism. The leaf superoxide dismutase activity was high (340.827 U∙g-1), the total antioxidant capacity was up to 834.016 U∙g-1, the total phenolic content was high (16.482 mg∙g-1), and the malondialdehyde content was as low as 10.490 nmol∙g-1, which indicated that P. acidula decreased the degree of oxidative damage by improving its antioxidant capacity. The suitable soil nutrient contents were low, soil calcium content was high, soil water content was as low as 9.5%, showing poor soil environment. But the leaf nitrogen and phrophros contents were high, showing high efficiencies in absorbing and utilizing nutrients. Based on the above results, P. acidula has strong ability in resisting drought and stress, suitable for growing in tropical coastal beach habitats with relatively poor soil, and is a potential species for rehabilitation in tropical coral islands.
Pemphis acidula; biological characteristics; beach plant
10.16258/j.cnki.1674-5906.2017.12.010
Q945.79; X173
A
1674-5906(2017)12-2064-07
曹策, 简曙光, 任海, 王俊, 刘楠, 王佳新, 吴淑华. 2017. 热带海滨植物水芫花(Pemphis acidula)的生理生态学特性[J]. 生态环境学报, 26(12): 2064-2070.
CAO Ce, JIAN Shuguang, REN Hai, WANG Jun, LIU Nan, WANG Jiaxin, WU Shuhua. 2017. The Ecophysiological Characteristics of Pemphis acidula, A Tropical Beach Plant [J]. Ecology and Environmental Sciences, 26(12): 2064-2070.
国家重点研发计划项目(2016YFC1403002);中国科学院A类战略性先导科技专项(XDA13020500);“十二五”农村领域国家科技计划项目(2015BAL04B04);广东省科技计划项目(2016A030303044)
曹策(1992年生),男,硕士研究生,研究方向为森林经理学。E-mail: 2421279691@qq.com
*通信作者。E-mail: liunan@scbg.ac.cn
2017-10-09
猜你喜欢
天天爱科学(2022年12期)2022-11-10 08:33:28
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
装备制造技术(2020年1期)2020-12-25 05:19:10
新疆农业科学(2020年1期)2020-02-14 03:22:46
首都食品与医药(2017年15期)2017-11-03 05:55:30
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中国麻业科学(2015年5期)2015-12-28 06:22:11
航空学报(2015年4期)2015-05-07 06:43:30
中国现代中药(2014年12期)2014-09-26 11:18:09
中国药理学与毒理学杂志(2013年5期)2013-11-12 07:24:14
