
寡照胁迫对设施葡萄叶片光合特性的影响及评价*
2017-12-18 08:30李凯伟杨再强杨世琼
中国农业气象 2017年12期
李凯伟,杨再强,2**,肖 芳,王 琳,杨世琼
寡照胁迫对设施葡萄叶片光合特性的影响及评价*
李凯伟1,杨再强1,2**,肖 芳1,王 琳1,杨世琼1
(1.南京信息工程大学气象灾害预报预警与评估协同创新中心,南京 210044;2.南京信息工程大学江苏省农业气象重点实验室,南京 210044)
以“红提”葡萄(L.)为试材,于人工气候箱设置L1(80μmol·m-2·s-1)、L2(200μmol·m-2·s-1)两种辐射水平,分T1、T3、T5、T7、T9(持续胁迫1、3、5、7、9d)5种胁迫天数对试材进行寡照处理,以L0(600μmol·m-2·s-1)为对照(CK),胁迫结束后统一以L0辐射水平进行16d的恢复试验。测定葡萄叶片相对叶绿素含量、光合参数和叶绿素荧光动力参数等指标,利用主成分分析方法筛选所测指标定义胁迫程度指数(Light stress index, LSI),结合恢复生长法划分寡照胁迫等级。结果表明:(1)寡照胁迫使葡萄叶片光合作用受到抑制,随着寡照胁迫程度加深,葡萄叶片的相对叶绿素含量(SPAD)、光饱和点(LSP)、最大光合速率(Pnmax)、表观量子效率(AQE)、气孔导度(Gs)、蒸腾速率(Tr)、光化学淬灭系数(qP)、光合电子传递速率(ETR)均表现为降低的趋势,非光化学淬灭系数(qN)呈上升趋势。L1处理的各指标变化幅度大于L2处理,胁迫9d时,Pnmax降至CK的26.5%,qN增至CK的1.89倍。Fv/Fm随胁迫时间表现为先增大后减小的趋势,L1T5处理达最大为0.79;(2)寡照胁迫后,0~8d为葡萄叶片光合参数快速恢复阶段,恢复至相对稳定状态需要12d左右。短时轻度寡照胁迫对葡萄叶片光合作用无明显抑制,甚至在恢复光照后表现出一定的刺激作用。长时重度寡照胁迫会对葡萄叶片光合系统造成不可逆损伤。(3)综合不同水平胁迫后葡萄叶片光合参数的恢复情况,结合恢复12d时葡萄叶片的LSI值,将设施葡萄寡照灾害分为无灾0级(L1寡照1d,L2寡照1~3d),轻度灾害Ⅰ级(L1寡照2~3d,L2寡照4~7d),中度灾害Ⅱ级(L1寡照4~7d,L2寡照8~9d),重度灾害Ⅲ级(L1寡照7d以上,L2寡照9d以上)4个等级。
寡照胁迫及恢复;胁迫程度指数;恢复生长法
葡萄(L.)属葡萄科,是落叶藤本植物,设施葡萄栽培是经济效益高、发展前景广阔的高效林果产业[1]。近年来,中国设施葡萄发展迅速,截至2013年,设施葡萄栽培面积超过13.3万hm2,居世界第一[2]。设施栽培的发展,扩大了栽培区域,延长了果品上市供应期,显著提高了葡萄产业的经济效益。寡照是设施环境中比较突出的一个环境问题,葡萄植株的生长发育对寡照胁迫较为敏感[3],而光合和荧光动力参数是植株叶片生理活动对寡照胁迫反应的主要探针,因此,研究寡照对葡萄叶片光合特性的影响对揭示设施葡萄寡照致灾规律具有重要意义。
国内外关于寡照对设施果树生长发育的影响有一定报道,对设施葡萄逆境生理的研究主要集中在干旱[4]、低温[5]、高湿[6]、盐分[7]胁迫等,前人的研究表明,寡照胁迫使苹果[8]、桃[9]、葡萄[10]的植株叶片变大变薄,叶色变淡,角度平展;弱光环境中的葡萄叶片对环境产生了一系列的生理适应,比叶面积、比茎增加,根冠比增加,遮阳90%时葡萄植株出现黄化现象[11-12];一般来说,短期寡照胁迫下,光饱和点、补偿点降低,气孔导度减小,蒸腾减弱,水分利用率增大,有利于植物在弱光下维持碳平衡。光照强度减弱使用于光合作用的能量减少,使得净光合速率降低[13-15]。关于叶绿素荧光参数,一些研究表明,遮光条件下光系统Ⅱ活性下降,遮光时间越长、强度越大,qP下降的幅度越大[16-17]。弱光处理可使脐橙[18]的最大光能转换效率Fv/Fm和Fm增加,并且始终高于对照,表明弱光环境下植物可以通过提高光化学效率来捕获更多的光能。
迄今为止,关于寡照对葡萄光合特性的影响缺乏系统性的研究,特别是对葡萄叶片在不同程度寡照胁迫处理后恢复水平的研究少有报道。本研究通过人工气候箱试验,系统研究葡萄叶片光合特性、荧光参数对寡照胁迫的响应并对胁迫等级进行划分,以期为设施葡萄寡照灾害防御及小气候环境优化调控提供依据。
1 材料与方法
1.1 试验设计
试验于2015年12月在南京信息工程大学人工气候箱(TPG1260,Australian)中进行。以1a生葡萄品种“红提”设施盆栽植株为试材。盆的规格为28cm(高)×34cm(上口径)×18cm(底径)。供试土壤为中壤土,pH7.4,有机质含量18.4g·kg-1,全氮0.79g·kg-1,全磷0.75g·kg-1,土壤体积含水量为32.5%。待植株叶片超过20叶后,于12月2日7:00起,每隔2d各放入3盆长势相同的植株在辐射水平为L1(100μmol·m-2·s-1)、L2(200μmol·m-2·s-1)人工气候箱内进行寡照处理,最后一次于12月10日放入,各处理按放入时间分别记为T9、T7、T5、T3、T1。同时设置辐射水平为L0(600μmol·m-2·s-1)的气候箱为对照(CK),在12月11日对所有处理进行测定。随后统一设置气候箱辐射水平为L0进行16d的恢复处理,每隔4d观测一次。胁迫开始前所有植株均在对照气候箱适应性处理3d,试验期间保证水分和养分在适宜水平,白天平均温度为25℃±1℃,晚上15℃±1℃,相对湿度设定75%±5%。
1.2 光合色素含量测定
使用日本产SPAD-502型相对叶绿素含量测定仪,每株选取从顶端向下数第5-8片功能叶片测量光合色素含量,每处理3株,重复测量3次。测定时间分别在12月11日、15日、19日、23日、27日的8:00-14:00,下同。
1.3 叶片光合参数测定
利用便携式光合作用测定系统LI-6400(LI-COR Inc,USA)测定叶片光合参数,测定时LI-6400的叶室内设定温度25℃,CO2浓度为390μmol·mol-1,光合有效辐射设置为1800、1500、1200、1000、800、600、400、200、100、50、20、0μmol·m-2·s-1共10个水平,分别测量不同PAR水平下的净光合速率。测量指标包括净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、大气CO2浓度(Ca,μmol·mol-1)及胞间CO2浓度(Ci,μmol·mol-1),由Ca和Ci计算得到气孔限制值Ls,即

水分利用率(Water Use Efficiency,WUE)的计算式为[19]
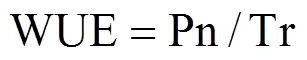
1.4 叶绿素荧光参数测定
利用LI-6400光合作用测定系统的6400-40荧光叶室测定荧光参数。在植株暗适应30min后,测定Fm、Fo等暗适应荧光参数,光下活化2h后测定光适应荧光参数Fm’、Fo’及 Fs等,根据以上参数计算得到PSII最大量子效率Fv/Fm、光化学猝灭系数qP、非光化学猝灭系数qN及电子传递效率ETR。光适应测定时光合有效辐射设定为600μmol·m-2·s-1。
1.5 葡萄寡照胁迫指数的确定

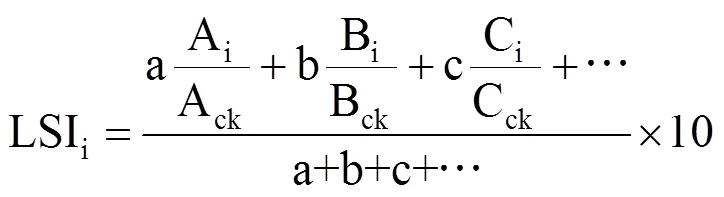
式中,A、B、C分别为筛选出的指标,i和ck分别表示不同处理和同期对照组的实测值。a、b、c为不同指标的权重,LSI代表葡萄叶片受寡照胁迫程度,其值越低表示受胁迫程度越深,对照组LSI值为10。
1.6 数据处理
利用Photosynthesis Work Bench程序进行光合作用光响应曲线拟合,得到光补偿点、光饱和点、最大净光合速率、暗呼吸速率、表观量子效率等参数。采用DPS软件及Microsoft Excel 2013对试验数据进行Duncan多重比较分析、相关性分析、主成分分析与图表绘制。
2 结果与分析
2.1 寡照胁迫对葡萄叶片相对叶绿素含量的影响
由图1a可见,寡照胁迫结束后,即恢复0d时CK处理的叶片相对叶绿素含量(SPAD)平均为36.2,短时寡照处理T1、T3的叶片SPAD平均值分别为 36.5和36.3,略高于CK(P<0.05),而寡照处理超过5d后叶片SPAD则显著降低,T5、T7、T9叶片SPAD平均值分别为35.2、34.5和34.1,明显可见在L1(80μmol·m-2·s-1)辐射水平条件下,随寡照时间延长叶片SPAD明显递减。在随后的16d恢复期中,短时寡照处理(T1、T3)叶片SPAD与CK处理中一致,均随恢复时间延长逐渐增加,且T1、T3一直高于CK;而T5、T7、T9处理中叶片SPAD一直保持较低水平,且显著低于CK(P<0.05),仅在恢复后期12d或16d时SPAD才略有提高。图1b显示,在L2(200μmol·m-2·s-1)辐射水平寡照处理结束时,所测叶片SPAD的排列顺序与L1水平时相同,但其变幅更小,T1、T3、T5、T7、T9处理叶片SPAD分别为36.6、36.7、35.9、35.8、38.4;而且,在整个恢复期T1、T3与CK差异不显著,其它处理则有随恢复时间延长而先降后升。可见,无论L1还是L2寡照水平,历经较短时间(1d、3d)寡照的叶片SPAD略有提高,且在恢复阶段保持较高的水平。但随着寡照时间的延长,叶片SPAD明显递减,辐射水平越低SPAD减小幅度越大,且恢复的越慢,达不到对照水平。
2.2 寡照胁迫对葡萄叶片光响应曲线参数的影响
利用Photosynthesis Work Bench程序进行光合作用光响应曲线拟合,得到不同辐射水平和持续寡照胁迫天数结束时葡萄叶片的光饱和点(Light Saturation Point,LSP)、光补偿点(Light Compensation Point,LCP)和表观量子效率(Apparent Quantum Efficiency,AQE)的变化情况(表1)。由表1可见,葡萄叶片LSP随着胁迫天数的增加呈现下降的趋势,L1辐射水平下,T1、T3处理的LSP下降幅度较小,分别为CK的85.1%和75.8%,T5处理显著(P<0.05)降低,为CK的45.9%。T7、T9则在同一水平,分别降至CK的37.4%和36.3%。LCP表现为先减小后增加的趋势,CK平均值为39.6μmol·m-2·s-1,T3处理即显著减小,T5处理达最小值,为21.6μmol·m-2·s-1,T9处理恢复至32.4μmol·m-2·s-1。AQE随着寡照天数增加而显著减小,T1、T3、T5、T7、T9处理的AQE分别为CK的79.5%、59.8%、51.2%、40.2%、33.9%。L2辐射水平处理与L1处理变化趋势相同,其T1和T3处理的LSP和AQE与CK在同一水平,无显著变化。相同胁迫天数下,L2处理的LSP和AQE值高于L1处理,且光合参数对寡照胁迫的响应落后于L1处理。可知,寡照胁迫使葡萄叶片LSP降低,AQE减小,LCP随胁迫天数增加呈先增加后减小的趋势。短期(1d、3d)寡照胁迫对葡萄叶片LSP和AQE影响较弱,胁迫5d时显著减小。LCP对寡照胁迫响应较为迅速,胁迫3d即显著降低,之后逐渐恢复。
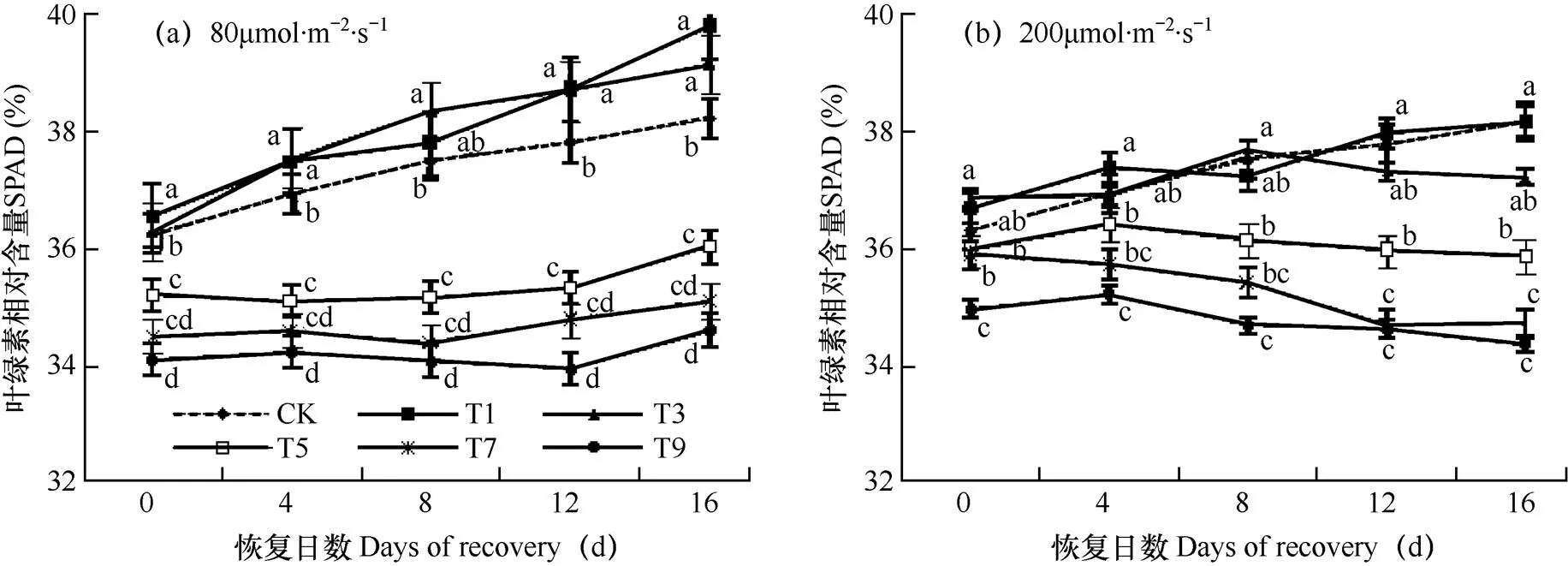
图1 两种寡照处理持续不同天数后葡萄叶片相对叶绿素含量随恢复时间变化的比较
注:L为辐射水平,T为持续时间;小写字母表示处理间Duncan检验在0.05水平上的差异显著性;短线表示均方差。下同
Note:L is the radiation level,T is the duration of stress. Lowercase indicates the difference significance among treatments at 0.05 level by Duncan’s significant test. The short bars mean square deviation. The same as below
由图2可知,经L1、L2辐射水平寡照胁迫后,葡萄叶片光响应曲线拟合所得最大光合速率(maximum photosynthetic rate,Pnmax)随胁迫时长的增加而显著减小(P<0.05)。在L1辐射水平处理下,T1、T3、T5、T7、T9处理的Pnmax分别降至CK的77.2%、53.8%、36.0%、32.0%、26.5%,胁迫超3d后Pnmax显著减小,之后保持稳定在相对较低水平。在L0辐射水平下恢复4d后,T1和T3处理的Pnmax分别为CK的95.0%和91.6%,恢复情况较好,T5、T7、T9处理较胁迫处理结束时有所好转,但Pnmax依然相对较低,为CK的59.1%、54.6%和49.0%。随着恢复时间的增加,各处理Pnmax比胁迫结束时均有不同程度的提高。恢复16d后,T1的Pnmax组比对照提高了6.6%,T3处理接近对照水平,其余各组处理分别恢复至对照的87.5%、71.3%、41.6%。在L2辐射水平处理下,胁迫相同天数Pnmax减小幅度较小于L1处理。恢复16d后,T1和T3恢复至对照水平,T5、T7、T9处理恢复至对照的96.4%、85.7%、54.6%。在恢复光照期间,各处理的Pnmax在0~4d迅速恢复,在8d之后保持相对稳定,变化幅度较小,经胁迫程度较弱(L1T1,L2T1,L2T3)的寡照处理后,在恢复期Pnmax会略高于CK。
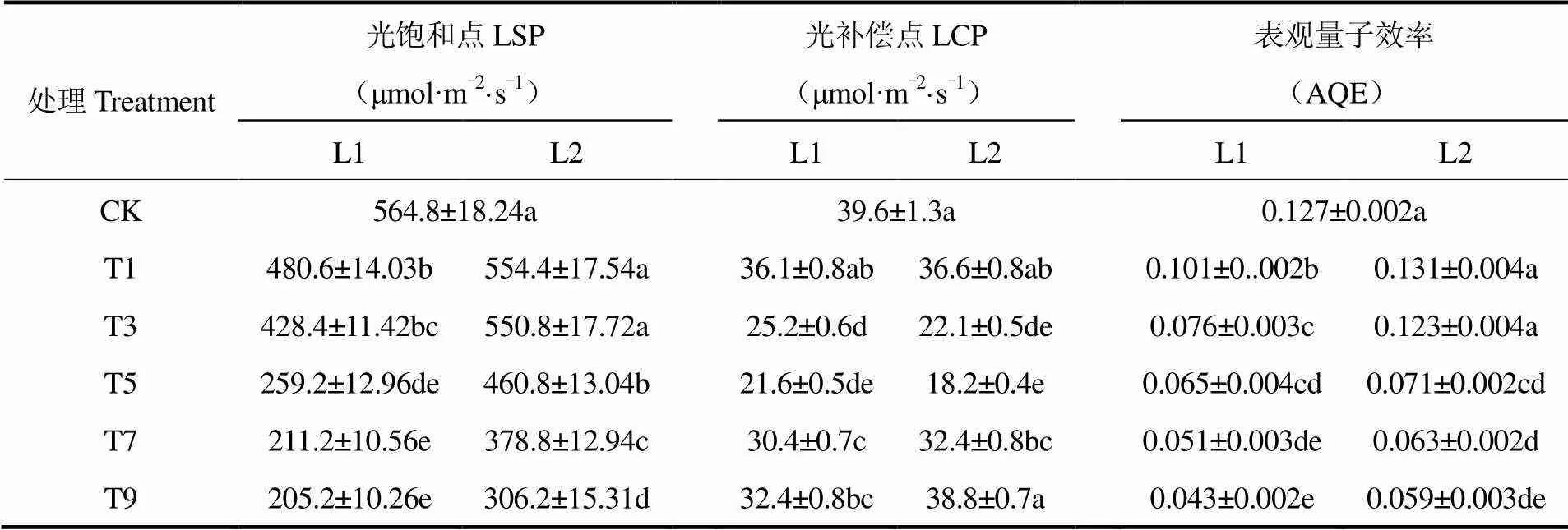
表1 两种寡照处理持续不同天数后葡萄叶片光合参数的比较

图2 两种寡照处理持续不同天数后葡萄叶片最大光合速率随恢复时间变化的比较
2.3 寡照胁迫对葡萄叶片气体交换参数的影响
不同寡照胁迫处理后植株在各辐射水平下的气体交换参数如图3所示,由图可见,寡照处理阶段,随着寡照时间的增加葡萄叶片的气孔导度(Gs)和蒸腾速率(Transpiration rate,Tr)呈减小的趋势,相同胁迫天数下Gs和Tr随光照强度的变化表现为光强越弱降幅越大。寡照处理5d以内,Gs和Tr迅速减小,之后降幅减小趋于平稳。L1辐射水平下,T1、T3、T5、T7、T9处理的Gs降至CK的72.9%、46.7%、24.5%、19.7%、15.5%,Tr降至CK的Ls随处理时间呈现出先减小后缓慢回升的趋势,L1辐射水平寡照处理1d、5d、9d后,Ls分别为0.61、0.43、0.57。葡萄叶片水分利用效率(WUE)在寡照处理初期无明显变化规律(图3d),T1、T3、T5处理与CK无明显差异,T7、T9处理低于CK。L1辐射水平下WUE一直保持在较低水平,L2辐射水平下表现为先增加后减小。寡照处理9d后,L1、L2辐射水平下的WUE分别降至CK的77.4%、57.2%,表明寡照胁迫9d可使葡萄叶片水分利用效率降低。整体上L2处理与L1处理变化趋势相同,相同胁迫天数下,气体交换参数的增减幅度小于L1。

图3 两种寡照处理及恢复期间叶片气体交换参数的动态变化
恢复处理阶段,L1组T9处理的Gs在0~4d内由0.0065mmol·m-2·s-1迅速增加至0.0182mmol·m-2·s-1,之后保持相对稳定。恢复16d后,L1、L2处理Gs分别为CK的86.1%和50.3%。Ls在恢复阶段随处理时间持续升高,Tr则先增加后减小,恢复8d时达最大值。WUE于恢复4d时突增,后又迅速降低,恢复期结束时L1和L2组的WUE无明显差异,且均低于CK。T9处理在恢复阶段气体交换参数随时间变化情况表明,0~8d为寡照胁迫后葡萄叶片气体参数快速恢复时期,其后各参数将趋于稳定。
2.4 寡照胁迫对葡萄叶片叶绿素荧光参数的影响
由图4a和4b可知,短期寡照胁迫(1d,3d)对葡萄叶片的PSII最大量子效率(Fv/Fm)影响较弱,各处理与CK无显著差异。L1辐射水平下,Fv/Fm随寡照时间先增加后减小,在寡照处理5d时增至0.79,9d后为0.76。L2辐射水平下则随寡照时间递增,9d时为0.80,辐射水平越低,葡萄叶片Fv/Fm对寡照胁迫的响应越迅速;寡照处理使光化学淬灭系数(qP)和光合电子传递速率(ETR)显著降低(图4c、4g),L1辐射水平下处理1、3、5、7、9d后qP为CK的58.5%、37.8%、35.2%、23.2%、22.6%,ETR为CK的62.2%、52.7%、42.9%、28.4%、24.2%;非光化学淬灭(qN)随着寡照处理时间的增加而升高,与光化学淬灭系数(qP)变化趋势相反(图4e、4f),L1辐射水平下,CK处理均值最小,为0.46,且与各组处理差异显著,T9最大,为0.87。T1、T3组在同一水平,达到对照的1.76倍。T5、T7、T9组在同一水平,为CK的1.89倍。L2辐射水平下,T1与CK无明显差异,qN在T3时迅速增大,为CK的1.47倍。T5、T7、T9无明显差异,达到CK的1.59倍。相同处理天数下,L1辐射水平下葡萄叶片的qN均高于L2。
在恢复阶段,L1辐射水平处理下的T1、T3、T5组,Fv/Fm在恢复4d时达CK水平。T7、T9处理随着恢复天数的增加而减小,恢复16d时T7、T9处理分别为CK的96.3%和80%(图4a、4b);qP恢复较快,L1辐射水平下除T9处理外均恢复至CK的75%以上。恢复16d后,L2辐射水平下各处理均达到CK的90%以上(图4c、4d);L1辐射水平下各处理qN值随恢复时间有增加的趋势,仅T1处理能恢复至CK水平。L2辐射水平下T1、T3、T5组恢复12d时达CK水平。恢复16d后T1、T3、T5均小于同期CK值(图4e、4f),表明一定程度的寡照刺激使葡萄叶片在适宜光照下热耗散降低;L2辐射水平处理下各组的ETR恢复程度要高于L1(图4g、4h),恢复期结束时L2辐射水平下各处理均恢复至CK的85%以上,L1辐射水平下T1恢复至对照水平,T3、T5、T7、T9组分别为CK的91.3%、85.1%、67.4%、59.1%。
2.5 寡照胁迫对葡萄叶片光合参数影响的综合评价
对本研究的所有指标进行主成分分析结果表明(表2),第一主成分以LSP、AQE、Pnmax、qP、qN、ETR、Gs、Tr为主,特征根向量最大,贡献率达64.2%。第二主成分中LCP和WUE具有绝对值较大的特征向量,贡献率为14.3%。第三主成分主要包括Fv/Fm和Ls,贡献率为10.2%。第四主成分中SPAD、Fv/Fm和Ls特征向量绝对值较大,贡献率为6.4%。前四主成分累计贡献率达95.0%,足够描述所有数据。同时,由于众指标存在较高的相关性,所测定数据反映的信息存在一定的重叠性。因此,对前四个主成分里的信息进行筛选可以得到简化的寡照胁迫指标。

图4 两种寡照处理持续不同天数后叶片叶绿素荧光参数随恢复天数变化的比较
叶绿素作为植物光合作用过程中最重要的色素,是光合作用的物质基础和光敏化剂,与植被的光能利用及转化效率密切相关,SPAD在不同寡照处理下差异显著,恢复期各处理依然保持一定的差异性,且与其它指标相关性较高,有一定的代表性,可作为寡照胁迫的指标;在光合参数中,光补偿点LCP能够反映出植物对弱光的适应能力[20],但因为葡萄叶片对寡照胁迫有一定的适应性,短期不同寡照处理下葡萄叶片的LCP变化规律不明显。同时LCP不能直接测量,需要通过光响应曲线拟合得出,存在一定误差。其在主成分分析中特征向量较小,所以不能作为寡照胁迫的指标。光饱和点LSP表示植物对强光的适应能力,表观量子效率AQE是光响应曲线0~200μmol·m-2·s-1区间的斜率,反映对弱光的利用能力。最大光合速率Pnmax反映植物的光合能力上限,气孔限制值Ls与Pnmax相关性为-0.22(未通过显著性检验),表明限制光合速率的并不是气孔因素。LSP、AQE、Pnmax三者之间相关性极高,且特征向量近似,故选择Pnmax作为寡照胁迫指标,来表达寡照胁迫对葡萄叶片光合潜能的伤害程度;选择最大量子效率Fv/Fm作为寡照胁迫指标。战吉宬对葡萄的研究表明[11],弱光环境中的葡萄叶片在强光下光合速率降低与暗适应下的叶绿素荧光参数Fv/Fm有密切关系。Fv/Fm能够反应出植物受胁迫程度,且在第一主成分和第四主成分有一定的贡献率。光化学淬灭qP与非光化学淬灭qN 极显著相关(r=-0.88**),qN反映光系统Ⅱ吸收的光能以热能形式耗散的部分[21],可表征寡照胁迫后葡萄对光能的利用能力,且特征向量绝对值要高于qP,所以选择qN作为胁迫指标;ETR反映实际辐射水平下的表观电子传递效率,在第一主成分中占到0.33,由于其受环境影响波动较大,且与Pnmax、Fv/Fm和qN极显著相关(r=0.91**,-0.94**,-0.90**)存在重叠,故不作为胁迫指标。
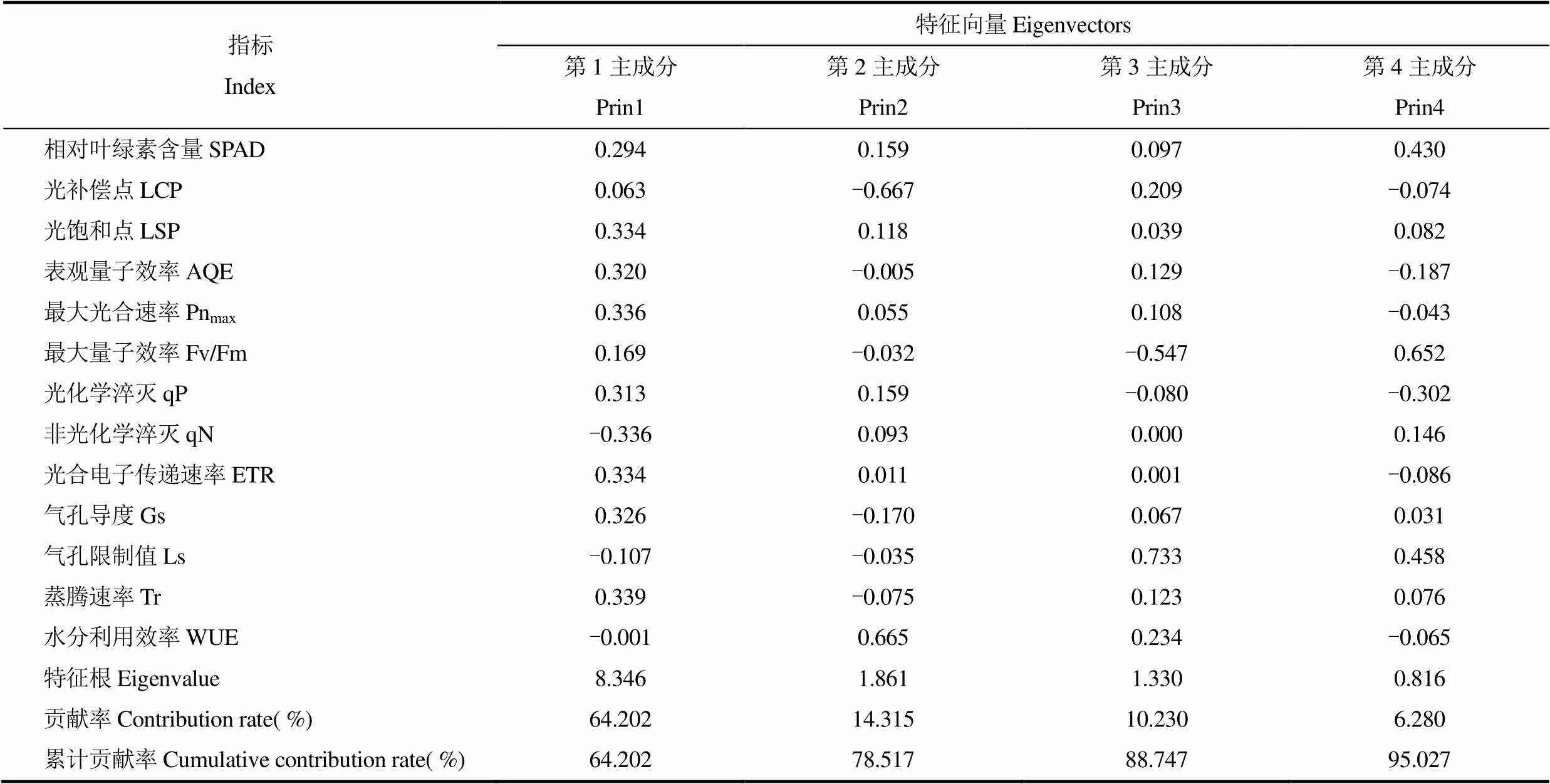
表2 葡萄叶片光合参数的主成分分析结果
综上所述,筛选出SPAD、Pnmax、Fv/Fm和qN 4个指标来定义葡萄叶片受寡照胁迫程度,再次对筛选出的4个指标进行主成分分析,根据每个指标的特征向量和贡献率计算得到每个指标在寡照胁迫程度指数(LSI)中的贡献度,代入式(3)得到
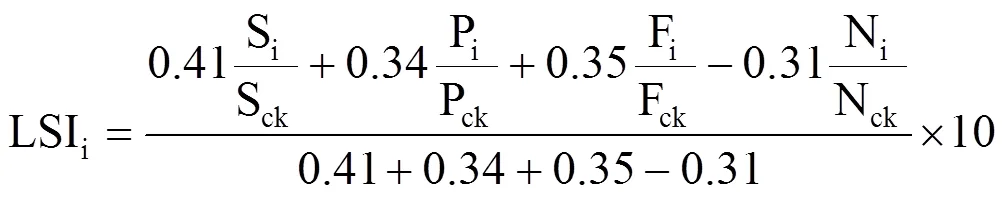
2.6 寡照胁迫程度指数分析
由表3可知,胁迫较轻的L1(T1、T3)处理和L2(T1、T3、T5、T7)处理,其LSI随恢复时间表现为增加的趋势,恢复8d后LSI趋于稳定,部分处理(L1T1、L2T1、L2T3)在恢复期间LSI高于CK。L1(T5、T7、T9)和L2(T9)处理LSI随恢复时间先增加后减小,除受胁迫最严重的L1T9处理最大值出现时间为4d外,其余均为8d。可见,0~8d为寡照胁迫后葡萄叶片快速恢复阶段,若胁迫程度过于严重,其LSI在快速恢复阶段后持续降低或稳定于较低水平,不能恢复至CK。故可推断寡照胁迫后,葡萄叶片恢复至最佳水平需要8~12d。采用恢复生长法[22]对设施葡萄寡照胁迫程度进行等级划分,将恢复12d时LSI值大于9.5即恢复至CK的95%以上的处理,称无灾,8.5~9.5为轻度灾害,6~8.5为中度灾害,小于6为重度灾害。为方便实际应用,按寡照胁迫天数可以划分为无灾0级(L1寡照1d,L2寡照1~3d),轻度灾害Ⅰ级(L1寡照2~3d,L2寡照4~7d),中度灾害Ⅱ级(L1寡照4~7d,L2寡照8~9d),重度灾害Ⅲ级(L1寡照7d以上,L2寡照9d以上)。

表3 两种寡照处理持续不同天数后的胁迫程度指数评价
3 结论与讨论
设施作物的生长和发育对光照有很强的依赖性,赵光强[23]在弱光对葡萄生长发育影响机理的研究中发现,弱光环境下,葡萄叶片单位重量叶绿素含量升高,但单位面积叶绿素含量较对照低。SPAD在一定程度上代表着单位面积叶绿素含量。本试验中,SPAD随着寡照时间的增加而降低,与赵光强研究结果一致。其中,葡萄叶片SPAD对寡照胁迫响应迅速。SPAD与最大光合速率呈现极显著正相关(r=0.81**),与吴月燕[17]对葡萄的研究结果相互印证。在光照恢复过程中,短期(T1,T3)轻度寡照处理的SPAD高于CK,原因可能是适宜程度的寡照胁迫锻炼引起植株应激反应,对葡萄叶片的叶绿素合成产生促进作用。严重寡照胁迫可能对葡萄叶片叶绿素合成系统产生不利影响,L1、L2分别寡照胁迫5d和7d即可使葡萄叶片SPAD在恢复期结束时远低于CK,对光响应曲线参数的分析表明,寡照胁迫使葡萄叶片LSP降低,AQE减小,LCP随胁迫天数先增加后减小。短期(T1,T3)轻度寡照胁迫对葡萄叶片LSP和AQE影响较弱,胁迫5d时则显著减小。LCP对寡照胁迫响应较为迅速,胁迫3d即显著降低,是葡萄叶片主动适应弱光环境的表现,以利用较弱的光合有效辐射。
最大净光合速率与气孔导度、蒸腾速率显著正相关,与气孔限制值无明显相关性(r=-0.22),非气孔因素造成净光合速率下降而胞间CO2浓度上升[24-25],说明寡照处理期间限制光合速率的主要因素并不是气孔因素而是光合系统本身,本结论与付涛等[26]对幼年鄞红葡萄的研究结果一致。在寡照胁迫条件下水分利用效率先增加后减小,且与光补偿点呈负相关,说明在寡照光抑制阶段葡萄叶片通过提高水分利用效率和降低光补偿点来提高光能利用率。
叶绿素荧光参数被认为是光合作用和环境关系的内在探针,能够反映各种胁迫引起的光合系统损伤或破坏[27]。葡萄叶片光系统II最大量子效率Fv/Fm可反映植物受光抑制程度,本试验中,Fv/Fm随寡照胁迫程度加深总体波动并不显著,但呈现先增加后减小的趋势,寡照胁迫使光化学淬灭系数qP和光和电子传递速率ETR降低,非光化学淬灭系数qN升高,用于光化学猝灭的比例减少,热耗散的比例增加,与武辉等[28]对棉花叶片的研究结果一致。分析葡萄叶片叶绿素荧光参数恢复状况可知,轻度寡照胁迫的叶绿素荧光参数可在恢复4d时达CK水平,中度胁迫恢复速度减慢,重度胁迫恢复16d后Fv/Fm、qP、qN、ETR均不能恢复至CK水平,推测葡萄叶片光系统II受到了不可逆的损伤。
本试验以新梢生长期设施盆栽红提葡萄为试材,定量研究不同辐射水平和天数的寡照胁迫对葡萄叶片光合特性的影响,及胁迫解除后光合特性的恢复情况。本研究证实,寡照胁迫使葡萄叶片光合作用受到抑制,0~8d为葡萄叶片光合参数快速恢复阶段,要达到相对稳定状态需要12d左右。短时轻度寡照胁迫对葡萄叶片光合作用无明显抑制,甚至在恢复光照后表现出有一定的刺激作用。长时重度寡照胁迫会对葡萄叶片光合系统造成不可逆损伤。根据不同胁迫水平后植株光合参数的恢复情况,将设施葡萄寡照灾害分无灾0级(L1寡照1d,L2寡照1~3d),轻度灾害Ⅰ级(L1寡照2~3d,L2寡照4~7d),中度灾害Ⅱ级(L1寡照4~7d,L2寡照8~9d),重度灾害Ⅲ级(L1寡照7d以上,L2寡照9d以上)4个等级。该结果可为设施葡萄的寡照灾害防御、农气服务及光环境调控提供科学依据。寡照胁迫对设施葡萄果实生长发育的影响更为重要,有待进一步研究。不同品种对光照条件的要求不同,该结论的适用性有待进一步检验。
[1]王玉忠,邵军辉,黄步青.设施葡萄产业发展现状与对策[J].农业科技与信息,2011,(9):36-37.
Wang Y Z,Shao J H,Huang B Q.Current situation andcountermeasures of grape industry development[J].Information of Agricultural Science and Technology,2011,(9):36-37.(in Chinese)
[2]穆维松,高阳,王秀娟,等.我国设施葡萄生产的成本收益比较研究[J].中外葡萄与葡萄酒,2014,(3):20-24.
Mu W S,Gao Y,Wang X J,et al.Comparative cost-benefit analysis on grape protected cultivation[J].Sino-Overseas Grapevine & Wine,2014, (3):20-24.(in Chinese)
[3]王海波,王宝亮,王孝娣,等.我国葡萄产业现状与存在问题及发展对策[J].中国果树,2010,(6):69-71.
Wang H B,Wang B L,Wang X T,et al.The present situation and existing problems of grape industry in China and the countermeasures[J].China Fruits,2010,(6):69-71.(in Chinese)
[4]陈丽,艾军,王振兴,等.干旱胁迫对山葡萄光合作用及光响应特性的影响[J].北方园艺,2011,(6):5-8.
Chen L,Ai J,Wang Z X,et al.Effect of drought stress on photosynthetic light response curve of[J].Northern Horticulture,2011,(6):5-8.(in Chinese)
[5]曹慧,赵升,王孝威,等.低温胁迫对葡萄幼苗相关生理指标的影响[J].安徽农业科学,2008,36(4):1351-1352.
Cao H,Zhao S,Wang X W,et al.Effects of low temperature stress on the related physiological indexes in grope seedlings[J].Journal of Anhui Agricultural Sciences,2008,36(4):1351-1352. (in Chinese)
[6]吴月燕,杨祚胜,丁伟红.高湿弱光对葡萄叶片光合生理生化指标的影响[J].浙江农业学报,2005,17(1):7-10.
Wu Y Y,Yang Q S,Ding W H.Effects of high humidity and poor light on photosynthetic physiological and biochemical indexes of grapevine leaves[J].Acta Agriculturae Zhejiangensis, 2005,17(1):7-10.(in Chinese)
[7]王艳杰,薛达元,彭羽.盐碱胁迫对两个葡萄品种光合作用-光响应特性的影响[J].北方园艺,2011,(16):30-36.
Wang Y J,Xue D Y,Peng Y.The photosynthesis-light response characteristics influence of two grape varieties under the stress of soil salinity-alkalinity[J]. Northern Horticulture,2011,(16): 30-36.(in Chinese)
[8]Jackson J E,Palmer J W.Effects of shade on the growth and cropping of apple trees I:experimental Détails and effects on vegetative growth[J].Journal of Horticultural Science,1977,52(2): 253-266.
[9]Kappel F,Flore J A.Effect of shade on photosynthesis,specific leaf weight,leaf chlorophyll content,and morphology of young peach trees[J].Journal American Society for Horticultural Science,1983,108:541-544.
[10]Bertamini M,Muthuchelian K,Nedunchezhian N.Iron deficiency induced changes on the donor side of PS II in field grown grapevine (L. cv.) leaves[J].Plant Science,2002,162(4):599-605.
[11]战吉成,黄卫东,王志龙,等.葡萄幼苗对弱光环境的形态和生长反应[J].中国农学通报,2002,18(2):1-2.
Zhan J C,Wang W D,Wang Z L,et al.Morphological and growth response of young grape plants to low light environment[J]. Chinese Agricultural Science Bulletin,2002,18(2):1-2.(in Chinese)
[12]武高林,陈敏,杜国祯.三种高寒植物幼苗生物量分配及性状特征对光照和养分的响应[J].生态学报,2010,30(1):60-66.
Wu G L,Chen M,Du G Z.Response of biomass allocation and morphological characteristics to light and nutrient resourcesfor seedlings of three alpine species[J].Acta Ecologica Sinica, 2010,30(1):60-66.(in Chinese)
[13]韩霜,陈发棣.植物对弱光的响应研究进展[J].植物生理学报,2013,(4):309-316.
Han S,Chen F D.Research progress in plant response to weak light[J].Plant Physiology Journal,2013,(4):309-316. (in Chinese)
[14]王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报,2011,31(7):1811-1817.
Wang J H,Ren S F,Shi B S,et al.Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters of[J].Acta Ecologica Sinica, 2011,31(7):1811-1817.(in Chinese)
[15]熊宇,杨再强,薛晓萍,等.遮光处理对温室黄瓜幼龄植株叶片光合参数的影响[J].中国农业气象,2016,37(2):222-230.
Xiong Y,Yang Z Q,Xue X P,et al.Effect of shading on photosynthetic parameters in greenhouse cucumber leaves[J].Chinese Journal of Agrometeorology,2016,37(2):222-230. (in Chinese)
[16]Deng Y,Li C,Shao Q,et al.Differential responses of double petal and multi petal jasmine to shading I: photosynthetic characteristics and chloroplast ultrastructure[J].Plant Physiology & Biochemistry,2012,55(2):93-102.
[17]吴月燕.两个不同葡萄种对高湿弱光气候的表现[J].生态学报,2004,24(1):156-161.
Wu Y Y.Response of grapes to low light intensity and high humidity[J].Acta Ecologica Sinica,2004,24(1):156-161.(in Chinese)
[18]韩春丽,孙中海,王艳,等.不同辐射水平对纽荷尔脐橙叶片PSⅡ功能和光能分配的影响[J].果树学报,2008, 25(1):40-44.
Han L C,Sun Z H,Wang Y,et al.Effects of different light intensity on the PS Ⅱ functions and absorbed light allocation in Newhall navel orange leaves[J].Journal of Fruit Science, 2008,25(1):40-44.(in Chinese)
[19]Bassman J B,Zwier J C.Gas exchange characteristics of,andxclones[J].Tree Physiology,1991,8(2):145-149.
[20]张振文,张保玉,童海峰,等.葡萄开花期光合作用光补偿点和光饱和点的研究[J].西北林学院学报,2010,25(1):24-29.
Zhang Z W,Zhang B Y,Tong H F,et al.Photosynthetic LCP and LSP of different grapevine cultivars[J].Journal of Northwest Forestry University,2010,25(1):24-29.(in Chinese)
[21]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
Zhang S R.Significance and discussion of chlorophyll fluorescence kinetic parameters[J].Chinese Bulletin of Botany,1999,16(4): 444-448.(in Chinese)
[22]许宏,王孝娣,邹英宁,等.葡萄砧木及酿酒品种抗寒性比较[J].中外葡萄与葡萄酒,2003,(6):20-23.
Xu H,Wang X T,Zou Y N,et al.Study on the cold resistance of grape rootstocks and wine grape cultivars[J].Sino-Overseas Grapevine & Wine,2003,(6):20-23.(in Chinese)
[23]赵光强.辐射水平、弱光对葡萄生长发育影响机理的研究[J].潍坊学院学报,2006,6(6):99-101.
Zhao G Q.Effects of weak light on the growth and development of grape[J].Journal of Weifang University,2006,6(6):99- 101.(in Chinese)
[24]韩艳,韩晨光,崔荣华,等.外源水杨酸对UV-B增强下花生叶片光合特性的影响[J].中国农业气象,2016,37(4):437-444.
Han Y,Han C G,Cui R H,et al.Effects of exogenous salicylic acid on photosynthetic characteristics of peanut leaves under elevated UV-B radiation[J].Chinese Journal of Agrometeorology,2016,37(4):437-444.(in Chinese)
[25]张曼义,杨再强,侯梦媛.土壤水分胁迫对设施黄瓜叶片光合及抗氧化酶系统的影响[J].中国农业气象,2017,38(1):21-30.
Zhang M Y,Yang Z Q,Hou M Y.Effects of soft water stress on photosynthetic characteristics and antioxidant enzyme system of cucumber leaves in greenhouse[J].Chinese Journal of Agrometeorology,2017,38(1):21-30.
[26]付涛,吴月燕,陶巧静,等.幼年鄞红葡萄对短期弱光胁迫的生理生化响应[J].江苏农业学报,2014,(2):200-204.
Fu T,Wu Y Y,Tao Q J,et al.Physiological and biochemical responsesof grape Yinhong seedlings to short-term weak-light stress[J].Jiangsu Journal of Agricultural Sciences,2014,(2): 200-204.(in Chinese)
[27]Baker N R.Chlorophyll fluorescence:a probe of photosynthesis in vivo[J].Plant Biology,2008,59(59):89-113.
[28]武辉,周艳飞,侯丽丽,等.低温弱光胁迫对棉花幼苗叶绿素荧光特性及能量分配的影响[J].新疆农业科学,2012,49(3): 393-399.
Wu H,Zhou Y F,Hou L L,et al.Effects of low temperature and weak light stress on chlorophyll fluorescence characteristicsand energy allocation in cotton seedlings[J].Xinjiang Agricultural Sciences,2012,49(3):393-399.(in Chinese)
Effects and Evaluation of Low Irradiation Stress on Photosynthetic Characteristics of Grapevine Leaves in Greenhouse
LI Kai-wei1, YANG Zai-qiang1, 2, XIAO Fang1, WANG Lin1, YANG Shi-qiong1
(1.Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters, Nanjing University of Information Science & Technology, Nanjing 210044, China; 2.Jiangsu Key Laboratory of Agricultural Meteorology, Nanjing University of Information Science & Technology, Nanjing 210044)
To study the effects of low irradiation (LI) on grape (L., cv) growth, a controlled experiment was carried out in growth chambers. Three levels of photosynthetic active radiation (PAR) were set for grapes:normal irradiation [L0, 600μmol(photon)·m-2·s-1] and low irradiation [L1, 80μmol(photon)·m-2·s-1; L2, 100μmol(photon)·m-2·s-1].The experiment lasted for 9d, then all groups of grapes were transferred to L0 to recover for 16d. The principal component analysis was used to screen the relative chlorophyll content, photosynthetic parameters, and chlorophyll fluorescence dynamics parameters; light stress index (LSI) was defined, and the stress level was classified combined with recovery growth. The results showed that, (1) the photosynthesis was inhibited by LI, relative chlorophyll content (SPAD), light saturation point (LSP), photosynthetic rate at irradiation saturation (Pnmax), apparent quantum efficiency (AQE), stomatal conductance (Gs), transpiration rate (Tr), photochemical quenching coefficient (qP), and electron transport rate (ETR) decreased under LI, while the non-photochemical quenching coefficient (qN) was opposite. The change of all indices under L1 was bigger than that of L2, after 9 d, Pnmaxdecreased to 26.5%, while qN increased to 189%, compared to CK. Maximal photochemical efficiency of PSII (Fv/Fm) increased at first, and then decreased with an increase of stress time, it peaked on the 5thd under L1. (2) The quick recovery stage of photosynthetic parameters was 0-8d, and it took about 12d to resume to CK or a relatively stable state.(3) Taking the recovery state of photosynthetic parameters into consideration, and combining with the LSI at 12d of recovery, the LI disaster level was divided into four grade, that is level 0 (1d under L1, and 1-3d under L2), levelⅠ (2-3d under L1, and 4-7d under L2), level Ⅱ (4-7d under L1, and 8-9d under L2), and level Ⅲ(more than 7d under L1, and more than 9d under L2).
Low irradiation stress and recovery; Light stress index; Regrowth method
10.3969/j.issn.1000-6362.2017.12.007
李凯伟,杨再强,肖芳,等.寡照胁迫对设施葡萄叶片光合特性的影响及评价[J].中国农业气象,2017,38(12):801-811
2017-04-27
。E-mail:yzq@nuist.edu.cn
江苏省科技支撑项目(BE2015693)
李凯伟(1993-),硕士生,主要从事设施葡萄气象灾害机理分析监测预警及服务技术应用。E-mail:564143539@qq.com
猜你喜欢
江西通信科技(2022年2期)2022-08-08
现代仪器与医疗(2021年4期)2021-11-05
保健医苑(2020年11期)2020-12-04
阅读(科学探秘)(2020年8期)2020-11-06
数学大王·中高年级(2020年5期)2020-05-25
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
现代园艺(2017年23期)2018-01-18
小天使·六年级语数英综合(2017年8期)2017-08-04
福建农业科技(2016年10期)2016-03-07
