
脊尾白虾全人工繁育技术初步研究
2017-12-18 11:23梁俊平李吉涛刘德月戴芳钰
水产科学 2017年3期
梁俊平,李 健,李吉涛,刘 萍,刘德月,戴芳钰
( 1.河南师范大学 水产学院,河南省水产动物养殖工程技术研究中心,河南 新乡453007;2. 中国水产科学研究院 黄海水产研究所,农业部海洋渔业可持续发展重点实验室,山东 青岛 266071;3. 青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛 266071 )
脊尾白虾全人工繁育技术初步研究
梁俊平1,2,李 健2,3,李吉涛2,3,刘 萍2,3,刘德月2,3,戴芳钰2,3
( 1.河南师范大学 水产学院,河南省水产动物养殖工程技术研究中心,河南 新乡453007;2. 中国水产科学研究院 黄海水产研究所,农业部海洋渔业可持续发展重点实验室,山东 青岛 266071;3. 青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛 266071 )
脊尾白虾是我国沿海地区池塘养殖重要品种,但目前尚未实现全人工繁育,严重制约了产业进一步发展。选取野生脊尾白虾作为亲虾,经过越冬培育、促熟交尾、幼体孵化培育、仔虾选育等手段,进行了脊尾白虾全人工繁育技术研究。试验结果显示,在盐度31.3、水温18.1~26.2 ℃、pH 8.1条件下,室内越冬脊尾白虾亲虾可成功培养至性腺成熟,并交尾抱卵,平均抱卵率达70%,平均孵化率为61.02%。幼体最佳培育密度20尾/L,仔虾适宜养殖密度0.13~0.53尾/L,培育至2 cm时存活率可达80%,继续培育3个月后发育至性成熟。一年可繁殖2~3代。在此基础上建立了脊尾白虾近交家系,现已培育至第6代,各家系繁殖力、生长速度和存活率均未出现近交衰退现象。本研究初步解决了脊尾白虾室内全人工繁育技术,并为试验动物培育奠定了良好的技术基础。
脊尾白虾;人工繁育;近交家系
脊尾白虾(Exopalaemoncarinicauda) 主要分布于我国整个大陆沿岸和朝鲜半岛西岸的浅海低盐水域[1-2],在我国沿海人工养殖规模超过约1.0×104hm2,是沿海池塘生态养殖主要特色虾种[3]。脊尾白虾作为海水长臂虾科白虾属的代表种,具有个体小、繁殖周期短、生长速度快、对毒性敏感以及容易饲养等特点,可作为甲壳动物发育学、遗传学、药理学、毒理学和疾病学等研究的理想试验动物[4-5]。然而,目前脊尾白虾养殖主要依靠捕捞天然苗种或投放野生抱卵亲虾在池塘自行繁殖,野生亲虾易携带病原,亲本遗传背景不清,苗种来源不稳定。随着养殖规模逐年扩大,各种病害也接踵而至,大规模发生和爆发性流行的趋势严峻,制约了脊尾白虾规模化养殖业的发展[6-8],也阻碍了其作为试验动物的培育进程。自20世纪80年代就开展了脊尾白虾人工繁育的相关研究,主要从野外捕获抱卵虾,在实验室条件下孵化、育出虾苗[9-10],但关于脊尾白虾的全人工繁育技术尚未解决。因此,开展脊尾白虾全人工繁育,切断其病原传播途径,可为脊尾白虾规模化人工养殖及试验动物化研究奠定良好的技术基础。
1 材料与方法
1.1 材料
野生脊尾白虾捕自莱州湾自然海区,共10 000尾,平均体长(50.8±6.3) cm,平均体质量(2.07±0.87) g。
1.2 方法
1.2.1 野生脊尾白虾室内越冬与培育
野生成虾运至实验室后,养殖在200 L PVC桶内,水体150 L,盐度31.3,水温18.1~26.2 ℃,pH 8.1,充气,每日8:00和17:00投喂配合饲料,定时吸污换水。历时6个月,至翌年3月,挑选体质健壮、附肢健全的性成熟亲虾200对,以雌雄比1∶1放入50 L PVC桶内进行交尾、产卵。
1.2.2 野生脊尾白虾幼体孵化和培育
孵化水质条件同上,幼体孵出后,亲虾捞出、测量体长、体质量后编号冷冻保存,计数幼体数量,计算受精卵孵化率。
光诱挑选活力强、胃肠饱满的幼体,设置5、10、20、30、40、50、60尾/L共7个培育密度梯度,养殖于50 L PVC桶内。水温22~26 ℃,微充气,投喂卤虫无节幼体。
1.2.3 F1代仔虾挑选及培育
挑选规格整齐、活力强、胃肠饱满、肝胰腺呈黄褐色的脊尾白虾仔虾,检测无病原携带。设置0.13、0.20、0.27、0.33、0.40、0.47、0.53、0.60、0.67 尾/L共9个密度,养殖于200 L PVC桶内,水体150 L,水温22~26 ℃,投喂人工配合饲料。
1.2.4 F1代脊尾白虾性成熟调控
选择体长2 cm的健康脊尾白虾,根据1.2.3的结果选择最佳养殖密度,雌、雄分养。设置24、26、28 ℃ 3个温度。投喂蛤蜊肉和鲜活沙蚕(Nereissuccinea),投饵量为虾体质量的10%~20%。蛤蜊肉切碎后置入150目筛绢网用消毒海水冲洗,除去油脂;沙蚕剪成0.5 cm长的小段,用消毒海水冲洗干净。早晚各投喂一次,蛤蜊肉和沙蚕隔天交替投喂。定时吸污换水,日换水量为20%~30%。
1.2.5 脊尾白虾近交家系建立
建立脊尾白虾近交家系(图1)。自第1代开始,依幼体孵化数量为选择指标,建立家系,各家系以英文字母编号,每个家系分4~5组培育,每代保持4~5个组,繁殖力下降的试验组则从其他组补充。统计每代幼体的孵化数量、存活率及生长速度。
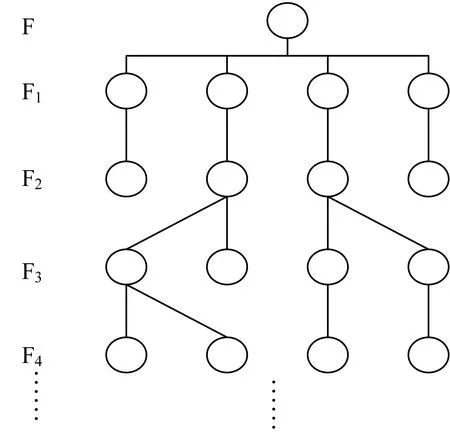
图1 脊尾白虾单个近交家系建立图谱
2 结 果
2.1 脊尾白虾亲虾的繁殖力
2.1.1 越冬脊尾白虾抱卵亲虾的规格
经越冬后,性成熟脊尾白虾平均抱卵率约70%,抱卵亲虾平均体长52.82 mm,平均体质量2.26 g,体长45~57 mm的抱卵虾占总抱卵虾的78.26%,体质量1.55~2.62 g的抱卵虾占总抱卵虾的72.73%(图2)。
2.1.2 越冬脊尾白虾亲虾孵化幼体数量和孵化率
每尾越冬亲虾孵化幼体数量差异较大,可孵化300~600尾幼体的亲虾占抱卵亲虾总数的52.46%,可孵化700~1000尾幼体的亲虾占抱卵亲虾总数的36.07%,孵化幼体数量低于100尾或高于1900尾的亲虾均只占抱卵亲虾总数的1.64%(图3)。

图2 室内越冬抱卵脊尾白虾体长和体质量的分布频率

图3 室内越冬脊尾白虾幼体孵化数量分布频率
根据体质量(m,g)与抱卵数量(y,粒)关系方程y=466.33m+133.9 (r2=0.9526),估算每尾亲虾理论抱卵量,再统计实际幼体孵化数量,计算出抱卵亲虾孵出幼体的孵化率为37.15%~94.85%,平均为61.02%(图4)。
2.1.3 不同密度下幼体及仔虾的存活率
水温22~26 ℃下,脊尾白虾溞状幼体投喂卤虫无节幼体,幼体培育密度为2、10、20尾/L时,变态为仔虾时的存活率均在80%以上(图5),3组间无显著差异(P>0.05),当密度超过30尾/L时,随着密度升高存活率显著下降(P<0.05)。自仔虾开始投喂人工配合饵料,培育50 d后,仔虾平均体长达2 cm,各组存活率见图6。仔虾适宜养殖密度为0.13~0.53尾/L。

图4 室内越冬脊尾白虾孵出幼体的孵化率

图5 不同养殖密度下幼体变态为仔虾时的存活率
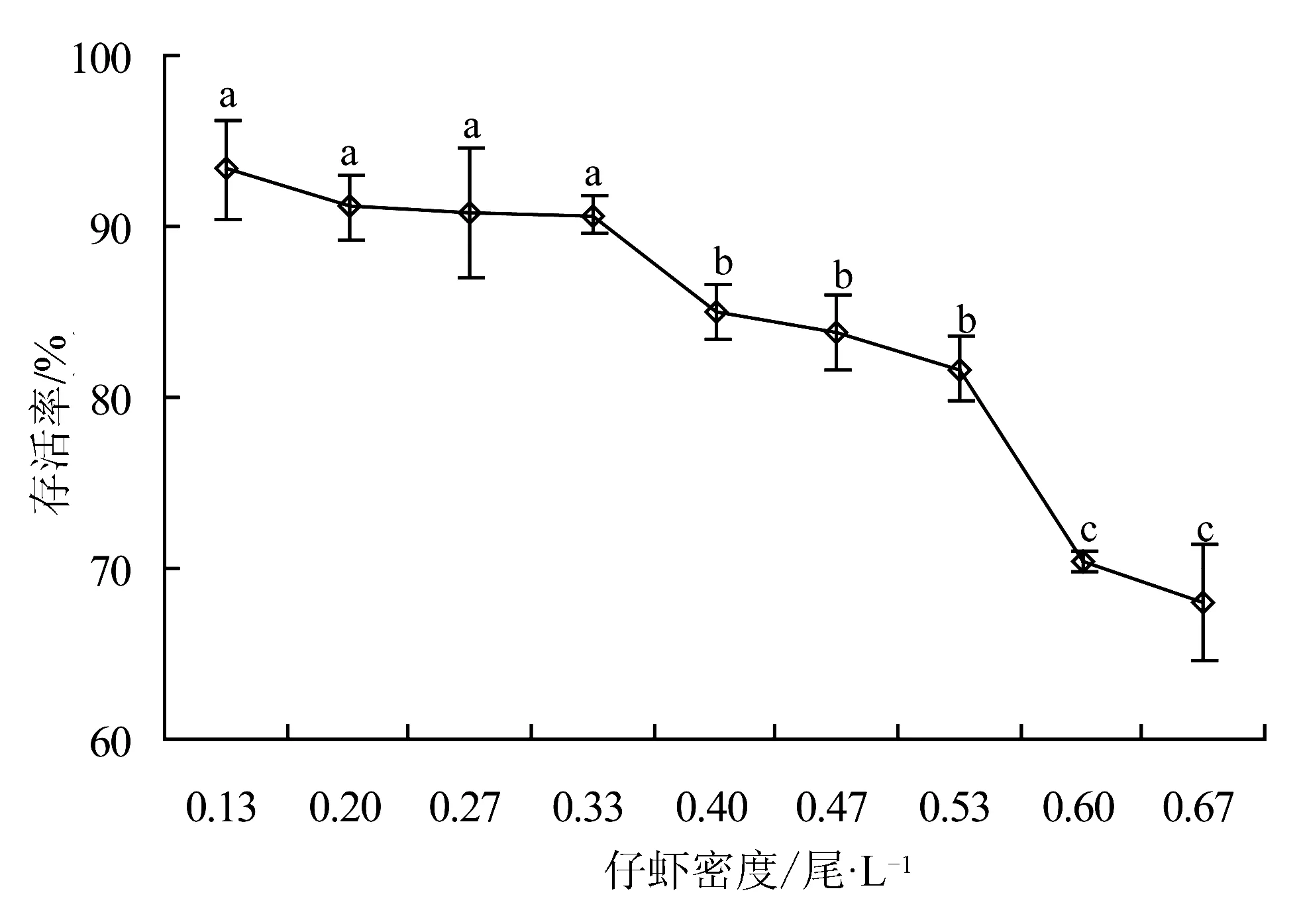
图6 不同养殖密度下仔虾的存活率
2.1.4 不同温度对脊尾白虾性腺发育的影响
脊尾白虾卵巢一对,前后端愈合,中央分离,前端游离,后端伸向头胸甲腹面,一对输卵管分别在卵巢两侧伸出,末端开口于第三步足基部雌性生殖孔。发育初期的卵巢呈半透明乳白色,中央分离区明显;成熟的卵巢呈棕黄色或淡绿色,卵膜表面分布有褐色斑点,卵巢布满了整个心区、肝区和胃区,外观不易观察到中央分离区(图7)。体长2 cm的健康脊尾白虾在24、26、28 ℃下培育80 d后,分别有18%、55%和61%的个体性腺发育至Ⅲ期(图8)。将发育到Ⅲ期的雌虾培育温度调至24 ℃进行培育,将发育到Ⅰ、Ⅱ期的雌虾培育温度调至28 ℃进行培育,继续培育30 d后,两温度组80%以上亲虾性腺发育到了Ⅳ期。

图7 脊尾白虾成熟卵巢外部形态特征

图8 养殖80 d后性腺发育到不同时期的雌虾的百分率
2.2 脊尾白虾近交家系建立
2.2.1 脊尾白虾近交家系系谱及培育
现已建立4个近交家系,培育代数见表1,培育速度最快家系已达6代。

表1 脊尾白虾近交家系培育代数
2.2.2 脊尾白虾近交家系幼体孵化数量及存活率
脊尾白虾近交家系每代亲虾孵出幼体数量及存活率见图9、图10,各代亲虾平均孵出幼体尾数在400尾以上,最高可达800尾。幼体发育到仔虾的存活率70%~80%,随着培育代数增加仔虾存活率均未明显下降。

图9 各家系平均每尾亲虾孵出幼体数量

图10 各家系每代幼体变态为仔虾时的存活率
2.2.3 脊尾白虾近交家系生长率
各家系每代的个体生长率见图11,K和JZW家系的生长率相对高于A和H家系,各家系的生长率随着培育代数增加均未下降。

图11 各家系每代个体体质量月增长速率
3 讨 论
3.1 脊尾白虾室内越冬培育
长臂虾科是十足目长臂虾总科中种类最多、分布最广的科,包括2亚科102 属,在海水、淡水中均有分布。近年来,关于脊尾白虾的研究逐渐增多,包括免疫、疾病、繁殖等[11-14],其中关于脊尾白虾人工繁育研究也有零星报道,但均为捕获野生抱卵亲虾进行繁殖,尚未达到全人工控制。徐嘉波等[15]进行了脊尾白虾家系育苗试验,但其亲虾培育仍在水泥池内进行,越冬温度低于10 ℃,摄食较少。本研究选择野生脊尾白虾,室内养殖于200 L桶内,越冬温度控制在18~26 ℃,为脊尾白虾适宜生长温度,保证了亲虾正常摄食,加快了性腺发育,越冬培育后,亲虾性腺均可发育成熟。通过一对一配对交尾,抱卵率可达70%,远远高于池塘养殖脊尾白虾的抱卵率29.31%[16]。为避免雄虾干扰,抱卵后亲虾单独养殖,幼体孵化率平均为61.02%,高于室外池塘(46%)[16]和水泥池孵化率(47%)[17]。
3.2 脊尾白虾溞状幼体培育
脊尾白虾刚孵出的幼体形态类似对虾糠虾幼体。武文魁等[18-19]最早将脊尾白虾幼体划分为了溞状幼体Ⅰ~Ⅵ期,而栗治国等[20]将脊尾白虾幼体发育划分为了糠虾幼体Ⅰ~Ⅴ期。笔者以6个时期溞状幼体发育指数和成活率为指标,研究了不同温度对幼体发育的影响,得出溞状幼体最佳培育温度为22~26 ℃[2]。脊尾白虾溞状幼体Ⅱ~Ⅲ期是食性转变期,也是死亡率较高时期,因此饵料选择及投饵量是提高幼体成活率的重要因素[21]。施祥元等[9,22]发现,脊尾白虾溞状幼体摄食蛋黄和豆浆时,发育至仔虾的成活率显著低于摄食卤虫(Artemiasp.)无节幼体的幼体。本研究中,在22~26 ℃水温下,全程投喂卤虫无节幼体,脊尾白虾溞状幼体可顺利发育至仔虾;当培育密度不高于20尾/L时,培育至仔虾的成活率可达80%,高于施祥元等[9,15,17]的试验结果。
3.3 脊尾白虾仔虾后培育
本研究采用200 L PVC桶培育仔虾,全程投喂人工配合饵料,培育密度为0.13~0.53尾/L,培育至2 cm时,成活率可达80%以上。2 cm脊尾白虾经过3个月培育,性腺开始发育成熟,不同温度显著影响性成熟速度。王春琳等[23]研究了脊尾白虾人工促熟技术,切除眼柄和升温均可促进性腺发育,虽然切除眼柄促熟速度快于升温,但由于脊尾白虾个体较小,容易受伤,有较高的死亡率;而升温促熟获得的抱卵虾健康活泼,体质不会变弱,与自然抱卵虾无差异,因此就脊尾白虾而言,升温促熟更符合实际。本研究采用升温技术,可使80%脊尾白虾性腺同步发育成熟,解决了性成熟不同步的问题[3]。
3.4 脊尾白虾近交家系建立及试验动物开发前景
根据遗传背景,试验动物可分为近交系、突变系、杂交系等,通过连续20代以上的近交繁殖,基因纯合度可达98.6%以上,可为试验获得更加精确、理想的数据。而在试验动物化过程中的最大难题是克服近交衰退[24]。近交衰退在鱼类中研究较多,关于甲壳动物近交衰退研究相对较少。研究发现,近交对凡纳滨对虾(Litopenaeusvannamei)幼体存活率有显著影响,而对成体存活无显著性影响[25-26];对中华卤虫(A.sinica)的研究发现,近交对存活率影响很小[27]。本研究通过定向交尾,已建立了4个脊尾白虾近交家系,为避免近交繁殖力衰退,每个家系内又建立了若干组(图1),以保证每代选择繁殖力强的个体。目前,各家系繁殖力、幼体存活率及生长速度虽有差异,但随着代数的增加并未出现下降趋势,这为脊尾白虾试验动物化奠定了基础。

[1] 王庚申, 谢建军, 施慧, 等. 盐度对脊尾白虾生长及肝胰腺消化酶活力的影响[J]. 水产科学, 2014, 33(7):451-454.
[2] 梁俊平, 李健, 李吉涛, 等. 不同温度对脊尾白虾胚胎发育与幼体变态存活的影响[J]. 生态学报, 2013, 33(4):1142-1152.
[3] 梁俊平, 李健, 刘萍, 等. 脊尾白虾生物学特性与人工繁育的研究进展[J]. 中国农学通报, 2012, 28(17):109-116.
[4] 郑琰晶, 魏社林, 吴进孝, 等. Cu2+、Zn2+、SDS、DBS 对脊尾白虾的毒性试验[J]. 热带海洋学报, 2006, 25(5):87-90.
[5] 王李宝, 万夕和, 朱彬, 等. 二硫氰基甲烷对脊尾白虾的急性致毒实验[J]. 海洋水产研究, 2008, 29(5):120-123.
[6] 许文军, 谢建军, 施慧, 等. 池塘养殖脊尾白虾(Exopalaemoncarinicauda)感染血卵涡鞭虫的研究[J]. 海洋与湖沼, 2010, 41(3):396-402.
[7] 沈辉, 万夕和, 王李宝, 等. 白斑综合征病毒对脊尾白虾的致病性研究[J]. 海洋科学, 2013, 37(5):55-60.
[8] 王元, 房文红, 周俊芳, 等. 脊尾白虾肌肉微孢子虫病的病原和病理分析[J]. 上海海洋大学学报, 2013, 22(5):726-733.
[9] 施祥元, 冯兴钱. 脊尾白虾人工育苗试验[J]. 海洋渔业, 1987, 9(2):73-74.
[10] 肖化卿. 白虾人工育苗初步研究[J]. 海洋科学, 1988,10(1):42-44.
[11] 王有昆, 刘萍, 李吉涛, 等. 脊尾白虾14-3-3基因cDNA全长的克隆和表达分析[J]. 中国水产科学, 2016, 23(1):44-52.
[12] Xu W, Xie J, Shi H, et al. Hematodinium infections in cultured ridgetail white prawns,Exopalaemoncarinicauda, in eastern China [J]. Aquaculture, 2010, 300(1):25-31.
[13] Duan Y, Liu P, Li J,et al. Expression profiles of selenium dependent glutathione peroxidase and glutathione S-transferase fromExopalaemoncarinicaudain response toVibrioanguillarumand WSSV challenge [J]. Fish & Shellfish Immunology, 2013, 35(3):661-670.
[14] 段亚飞, 梁俊平, 李吉涛, 等. 鳗弧菌感染对脊尾白虾鳃组织抗氧化酶系统的影响[J]. 水产科学, 2015, 34(11):667-672.
[15] 徐嘉波, 严银龙, 朱雅珠. 脊尾白虾家系育苗试验[J]. 水产科技情报, 2009(6):301-303.
[16] 于天基, 李健, 李吉涛, 等. 池养脊尾白虾的生长与繁殖特性[J]. 中国渔业质量与标准, 2014, 4 (2) :16-25.
[17] 李国峰, 张振华, 严玉洲. 脊尾白虾在低盐度水体中的人工繁育试验[J]. 水产养殖, 2000(1):6-7.
[18] 武文魁. 脊尾白虾幼体发育的研究[J]. 海洋学报, 1984, 5(增刊):953-964.
[19] 梁象秋, 李亚娟, 周昭曼. 脊尾白虾的幼体发育[J]. 水产学报, 1988, 12(2):157-168.
[20] 栗治国, 张成松, 张岩, 等. 温度对脊尾白虾胚胎及幼体发育的影响研究[J]. 海洋科学, 2013, 37(10):9-16.
[21] 罗会明, 黄厚哲. 脊尾白虾幼体对饵料的摄食与吸收[J]. 厦门大学学报:自然科学版, 1980, 19(4):100-107.
[22] 陆开宏, 华建权, 陈贤龙. 人工培育脊尾白虾蚤状幼体的饵料基础研究[J]. 黄渤海海洋, 2001, 19 (4):63-70.
[23] 王春琳, 陈云彬, 陈阳明, 等. 脊尾白虾(Palaemoncarinicauda)人工促熟的初步研究[J]. 浙江海洋学院学报, 1995, 14(4):243-246.
[24] 胡建华, 姚明, 崔淑芳. 实验动物学教程[M]. 上海:上海科学技术出版社, 2009:24-30.
[25] Moss D R,Arce S M, Otoshi C A, et al. Effects of inbreeding on survival and growth of Pacific white shrimpPenaeus(Litopenaeus)vannamei[J]. Aquaculture, 2007, 272(S1):30-37.
[26] De los Ríos-Pérez L, Campos-Montes G R,Martínez-Ortega A, et al. Inbreeding effects on body weight at harvest size and grow-out survival rate in a genetic selected population of Pacific white shrimpPenaeus(Litopenaeus)vannamei[J]. Journal of the World Aquaculture Society, 2015, 46(1):53-60.
[27] 喻驰方, 栾生, 张志伟, 等. 中华卤虫不同近交水平家系生长性能比较[J]. 渔业科学进展, 2013, 34 (6):75-79.
[28] 吴志强, 邵燕, 袁乐洋. 鱼类实验动物[J]. 生物学通报, 2004, 38(11):20-22.
[29] 吴淑勤, 黄志斌, 石存斌, 等. 鱼类实验动物与剑尾鱼水生实验动物化[J]. 实验动物科学与管理, 2003, 20(增刊):20-21.
[31] 吴端生, 郑家铨, 刘冬娥, 等. 红鲫实验动物生物学特性的研究[J]. 中国实验动物学杂志, 1997, 7(2):79-81.
ControlledPropagationofRidgetailWhitePrawnExopalaemoncarinicauda
LIANG Junping1,2, LI Jian2,3, LI Jitao2,3, LIU Ping2,3, LIU Deyue2,3, DAI Fangyu2,3
( 1. Engineering Technology Research Center of Henan Province for Aquatic Animal Cultivation, College of Fisheries, Henan Normal University, Xinxiang 453007, China;2. Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China;3. Function Laboratory of Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China )
The aim of the present experiment is to provide the artificial breeding technology of ridgetail white prawnExopalaemoncarinicauda, using comprehensive measures such as overwinter culture, promotion of gonadal maturation, larval cultivation, and rearing of post-larvae. The results showed that the ridgetail white prawn were cultivated to be eggs-carrying broodstock at water temperature of 18.1—26.2 ℃, a salinity of 31.3 and pH 8.1, and induced to mate and spawn, with average success rate of 70%, and average hatching rate of 61.02%. The larvae were suitablly developed at density of 20 individual/L, and the post-larvae had suitable density of 0.13—0.53 individual/L, with the survival rate of 80% in the juveniles. The juveniles were cultivated for 80 days when the ovaries were mature. The artificial breeding technology of ridgetail white prawn was first succeeded, breeding 2—3 generations each year. Based on the artificial breeding technology, the 6 generations of ridgetail white prawn were bred, without significant recession of fecundity, growth rate and survival rate in the inbreeding families of ridgetail white prawn. The findings provide a basis for cultivating of ridgetail white prawn as a laboratory animal.
Exopalaemoncarinicauda; artificial breeding; inbred family
10.16378/j.cnki.1003-1111.2017.03.006
S968.22
A
1003-1111(2017)03-0290-06
2016-02-01;
2016-04-08.
国家自然科学基金资助项目(31472275);国家虾产业技术体系项目(CARS-47);泰山产业领军人才工程项目(LJNY2015002);青岛海洋科学与技术国家实验室鳌山科技创新计划项目(2015ASKJ02).
梁俊平(1981-),男,副教授,博士;研究方向:水产动物营养与健康养殖. E-mail: liangjunpinglover@163.com.通讯作者: 李健(1961-),男,研究员,博士生导师;研究方向:海水健康养殖技术. E-mail: lijian@ysfri.ac.cn.
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
渔业致富指南(2021年16期)2021-12-07
生物学通报(2021年5期)2021-03-16
海洋通报(2020年3期)2020-10-20
水产养殖(2020年9期)2020-09-24
当代水产(2020年3期)2020-06-15
渔业致富指南(2018年19期)2018-10-18
渔业致富指南(2018年18期)2018-10-10
河北渔业(2015年5期)2016-03-01
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
