
普通小麦 Glu-3位点基因单元型的分离和序列分析
2017-11-08 07:36李淑芬赵献林
麦类作物学报 2017年10期
董 雪,王 增,刘 梦,杨 燕 ,韩 冰,李淑芬,赵献林
(1.内蒙古农业大学生命科学学院植物生物技术功能实验室,内蒙古呼和浩特 010018; 2.河南省农业科学院小麦研究所/河南省小麦生物学重点实验室/小麦国家工程实验室/农业部黄淮中部小麦生物学与遗传育种重点实验室,河南郑州 450002)
普通小麦 Glu-3位点基因单元型的分离和序列分析
董 雪1,王 增1,刘 梦1,杨 燕1,韩 冰1,李淑芬1,赵献林2
(1.内蒙古农业大学生命科学学院植物生物技术功能实验室,内蒙古呼和浩特 010018; 2.河南省农业科学院小麦研究所/河南省小麦生物学重点实验室/小麦国家工程实验室/农业部黄淮中部小麦生物学与遗传育种重点实验室,河南郑州 450002)
为了从分子水平上探讨优质小麦资源中LMW-GS等位基因与小麦品质的关系,以及在改善小麦品质方面的潜在价值,利用小麦 Glu-A3和 Glu-B3基因的特异引物从强筋型、中筋型和弱筋型小麦共计10份材料中分离出LMW-GS基因后进行序列分析。结果表明,共发现14个新的核苷酸变异类型和4个肽链变异类型。其中,14个新的核苷酸变异类型中,4个为 Glu-A3基因变异类型,1个为 Glu-B3基因变异类型,9个为 Glu-D3基因变异类型。值得注意的是,有2个半胱氨酸数目特殊的亚基类型被发现,一个是来自师栾02-1含有9个半胱氨酸残基的 GluA3-18基因,另一个是来自偃展4110含有7个半胱氨酸残基的 GluD3-13基因。
普通小麦;低分子量麦谷蛋白亚基;基因分离; Glu-3位点
根据筋度的不同,小麦可分为强筋、中筋和弱筋三种类型,其中强筋和弱筋小麦一般称为优质麦。强筋型小麦多用于制作面包,弱筋型小麦多用于制作饼干、糕点等。然而,我国的强筋型小麦和弱筋型小麦品种较少,生产上种植的品种多数为中筋型小麦。
小麦贮藏蛋白主要包括麦谷蛋白和醇溶蛋白,而麦谷蛋白的含量对面筋强度有着重要的影响。根据分子量大小,可将麦谷蛋白分为高分子量麦谷蛋白亚基(HMW-GS)和低分子量麦谷蛋白亚基(LMW-GS)[1],其中,LMW-GS约占麦谷蛋白的60%。由于LMW-GS拷贝数较多,加之其分子量与醇溶蛋白分子量相似,传统分离方法SDS-PAGE很难将其区分开来,因此对LMW-GS的研究远不及HMW-GS那么深入。目前,采用特异性引物从小麦栽培品种及其近等基因系和近缘种中发现的LMW-GS基因、基因片断和假基因已经超过200个[1-5],其中在普通小麦中被报道的编码LMW-GS家族的 Glu-3基因位点约有70个[1]。许多研究认为, Glu-3位点编码的LMW-GS等位基因对小麦品质有重要影响,其中 Glu-A3位点上的 Glu-A3b和 Glu-A3d等位基因对增强面团特性和改善烘烤品质方面有着重要贡献[6-9]。大部分LMW-GS通过第一组染色体短臂上的 Glu-3( GluA-3、 Glu-B3和 GluD-3)位点编码[10],在其他位点上也相继有过报道,如在染色体1B上的 Glu-B2和 Glu-B4[11],在染色体1D上的 Glu-D4和7D上的 Glu-D5[12-13]。目前无论是在普通小麦或是近缘种材料中,对D染色体上的LMW-GS基因的研究要比对A和B染色体上的研究多一些。由于3个位点中不同等位基因的亚基间分子量很接近,且有的亚基鉴定需要依赖其浓度进行区分[2,13],所以很难发现新的亚基。由于LMW-GS对小麦加工品质的重要性,鉴定LMW-GS基因的等位变异及开发出相应的分子标记对小麦品质育种具有重要意义[14]。目前已研发出了一些LMW-GS基因的功能标记,如Zhang等[15]根据 Glu-A3位点的1个LMW-GS基因的等位变异开发出一套 Glu-A3位点等位基因的PCR标记,可以有效地鉴定出该位点的等位基因类型,并在小麦资源及育种材料的筛选中加以利用[16-18];Zhao等[19]从 Glu-D3位点扩增出6个LMW-GS基因的12个等位变异,并根据各基因等位变异之间的SNP或Indel开发出了7个单元型的STS标记。
目前,人们较少利用不同筋度的小麦为材料来研究其中的LMW-GS的基因。鉴于此,本试验利用已知不同筋力的普通小麦材料以及 Glu-A3和 Glu-B3位点的特异引物,对相应的LMW-GS基因进行扩增和鉴定,以期发现新的基因单元型,探讨不同筋力小麦品种的LMW-GS基因差异,为小麦品质育种中亲本的选择提供理论依据。
1 材料与方法
1.1 供试材料
供试材料共计10份,包括郑麦366(半冬性)、郑麦9023(弱春性)和师栾02-1(半冬性)3份强筋型,矮抗58(半冬性)、周麦18(半冬性)、石家庄8号(半冬性)和偃展4110(弱春性)4份中筋型,郑麦004(半冬性)、豫麦50(弱春性)和扬麦13号(春性)3份弱筋型,均由河南农业科学院小麦研究所提供。
1.2 DNA的提取
每份小麦材料取3~5粒种子置于洁净的培养皿中,在25 ℃条件下培养5~7 d,取2~3 g叶片,采用CTAB法[20]提取基因组DNA。
1.3 引物的筛选和PCR扩增
LMW-GS基因 Glu-A3位点和 Glu-B3位点上的特异性引物选自王林海[18]的文章,具体见表1。
PCR扩增体系50 μL,包括模板DNA 100 ng、上下游引物(20 μmol·L-1)各1 μL、TaqDNA聚合酶(5 U·μL-1,TaKaRa Ex) 1 μL、10×PCR buffer 5 μL、dNTP Mix (2.5 mmol·L-1)4 μL,其余用ddH2O补足。PCR扩增程序:94 ℃预变性5 min;94 ℃变性45 s,57~61 ℃退火45 s,72 ℃延伸1 min,32个循环;72 ℃延伸10 min,4 ℃保存。
1.4 PCR产物的纯化、克隆及测序
PCR产物电泳检测后,在紫外灯下切取目的条带,用天根公司生产的琼脂糖凝胶DNA回收试剂盒回收纯化(具体步骤见其说明书)。将纯化后的PCR产物连接至PMD19-T载体(北京全式金生物技术有限公司)上(连接体系:SolutionⅠ 5 μL、PMD19-T 1 μL、纯化后的PCR产物4 μL),16 ℃连接12 h,4 ℃保存。连接产物通过热击法进行转化,之后进行克隆子的鉴定,最后送南京金斯瑞生物技术有限公司进行测序,每个样品测3~6次重复。

表1 本研究所用引物Table 1 Primers used in this study
1.5 序列分析
用DNAMAN 6.0汉化版软件进行蛋白质翻译及多序列比对等处理;用Vector NTI Advance 11.5.2软件进行肽链分子量的计算;用MEGA 4.0软件构建基因进化树。
2 结果与分析
2.1Glu-A3基因等位变异的分离和序列分析
利用引物LA1F/LA1R对强筋、中筋、弱筋型10份小麦材料进行PCR扩增,共获得8个条带(图1A),经测序得到3个不同的序列。将这3个序列与GenBank中已知的 GluA3-1基因核苷酸和氨基酸序列进行比对分析,结果(表2)发现,获得的3个 GluA3-1单元型均不是新的等位变异类型,其中在郑麦366、郑麦9023、周麦18、偃展4110、豫麦50和扬麦13号中得到了相同的单元型 GluA3-13,在石家庄8号中得到了单元型 GluA3-17,在师栾02-1中得到了单元型 GluA3-18。不过,从强筋小麦品种师栾02-1中扩增的1个等位变异是首次在普通小麦中发现,且与本人先前从栽培二粒小麦DM2中扩增出的等位变异类型 GluA3-18(GenBank登录号为KX879094)相同。该基因序列长度为1 537 bp,编码区长度为1 164 bp。经与 GluA3-11(GenBank登录号FJ549928.1)进行对比后发现, GluA3-18在编码区的第212、596和965位碱基后分别插入了24、3和9 bp碱基。此外,该序列还存在19个SNP,其中11个为错义突变。值得注意的是,在924 bp处碱基C替换成T,使得对应氨基酸R替换成了C,导致 GluA3-18编码的氨基酸序列AP1-8含有9个半胱氨酸残基,多出的一个半胱氨酸位于半胱氨酸富集区。
利用引物LA2F/LA2R对强筋、中筋、弱筋型10份小麦材料进行PCR扩增,共获得9个条带(图1B),经测序得到4个不同的序列。将这4个序列与GenBank中已知的 GluA3-1基因核苷酸和氨基酸序列进行比对分析,结果(表2)发现,有2个序列为新的等位变异,命名为 GluA3-25和 GluA3-26(GenBank登录号分别为KX879099和KX879100),分别来自品种师栾02-1和豫麦50。 GluA3-25和 GluA3-26基因长度分别为1 100 bp和1 097 bp,编码区长度分别为912 bp和909 bp。在编码区域内,与 GluA3-21(GenBank登录号为FJ549935.1)相比,二单元型在第306个碱基处发生了无义突变,形成了终止密码子,为假基因。
利用引物LA3F/LA3R对强筋、中筋、弱筋型10份小麦材料进行PCR扩增,共获得9个条带(图1C),经测序得到4个不同的序列。其中,来自矮抗58和石家庄8号的序列为新等位变异,分别命名为 GluA3-37和 GluA3-38(GenBank登录号分别为KX879106和KX879107),序列长度分别为1 408 bp和1 393 bp,完整编码区长度分别为1 032 bp和1 017 bp。在编码区内,与 GluA3-31(GenBank登录号:FJ549939.1)相比, GluA3-37和 GluA3-38在第417 bp处发生了无义突变,形成终止密码子,为假基因。

A:引物LA1F/LA1R的扩增产物;B:引物LA2F/LA2R的扩增产物;C:引物LA3F/LA3R的扩增产物。M:DL2000;1:郑麦366;2:郑麦9023;3:师栾02-1;4:矮抗58;5:周麦18;6:石家庄8号;7:偃展4110;8:郑麦004;9:豫麦50;10:扬麦13号。
A:PCR products amplified with the primer LA1F/LA1R; B:PCR products amplified with the primer LA2F/LA2R; C:PCR products amplified with the primer LA3F/LA3R.M:DL2000; 1:Zhengmai 366; 2:Zhengmai 9023; 3:Shiluan 02-1; 4:Aikang 58; 5:Zhoumai 18; 6:Shijiazhuang 8; 7:Yanzhan 4110; 8:Zhengmai 004; 9:Yumai 50; 10:Yangmai 13.
图13对Glu-A3位点基因特异性引物在普通小麦中的PCR扩增产物
Fig.1PCRproductsofGlu-A3amplifiedwiththreegene-specificprimersfromcommonwheat
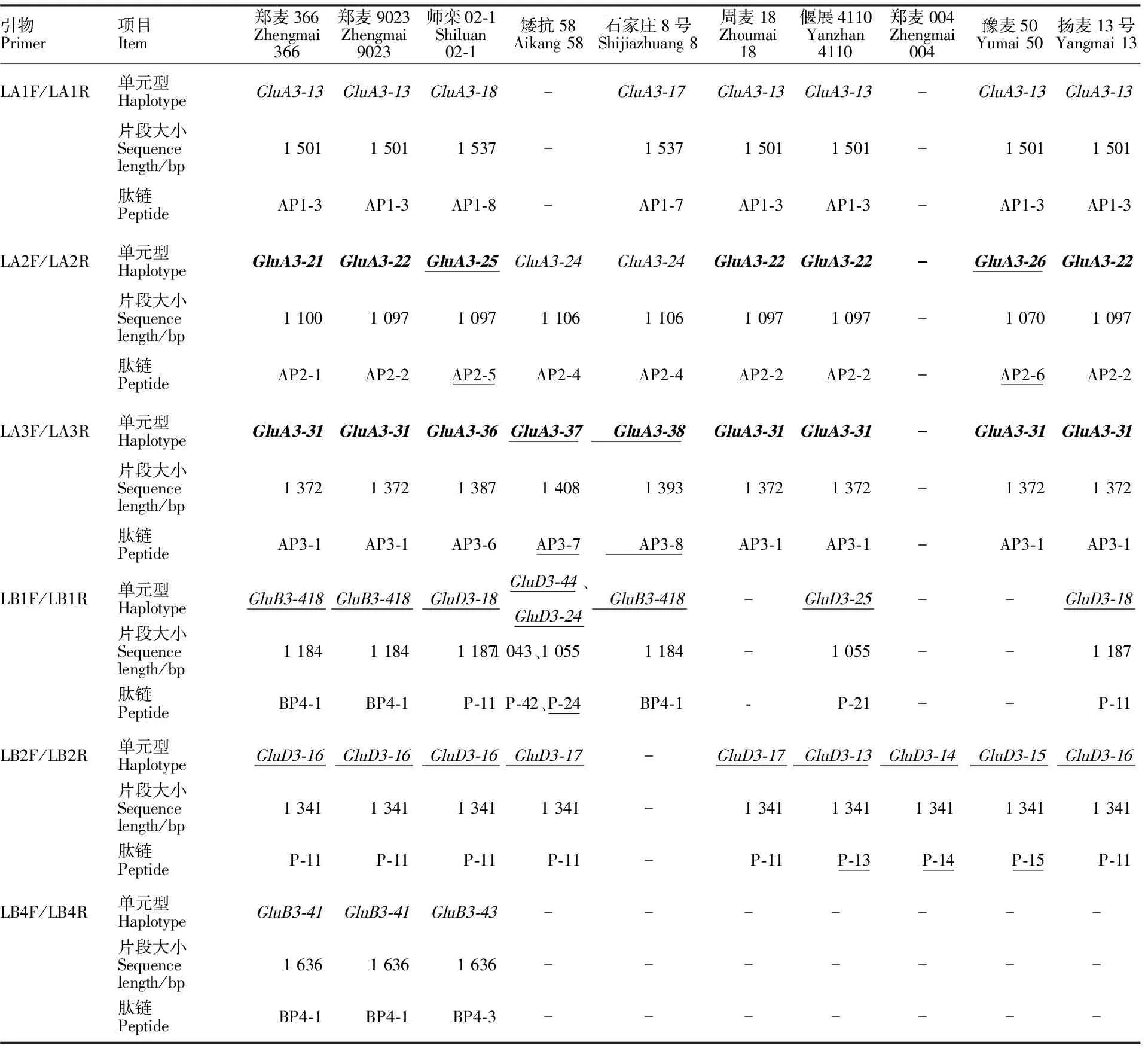
表2 10份普通小麦中扩增获得的单元型Table 2 Haplotypes amplified from the ten common wheats
加粗体表示假基因;“-”表示没有扩增出相应单元型;下划线表示新发现的单元型及肽链。
Pseudogenes are in bold; “-” represents no amplification; The newly discovered haplotypes and peptide chains are underlined.
2.2Glu-B3和Glu-D3基因等位变异的分离和序列分析
利用引物LB1F/LB1R对强筋、中筋、弱筋型10份小麦材料进行PCR扩增,结果(图2A)发现,在郑麦366、郑麦9023和石家庄8号中扩增出了相同的核苷酸序列(1 184 bp),与 GluB3-41(GenBank登录号为EU369724.1)的相似性高达99.9%,且编码区核苷酸序列相同(1 056 bp),仅在3′-UTR有1个变异碱基(T→C),将该序列命名为 GluB3-418(GenBank登录号:KY430288);在师栾02-1和扬麦13号中扩增的核苷酸序列也相同(1 187 bp),与 GluD3-11(GenBank登录号为DQ357052.1)的相似性高达99.8%,且编码区核苷酸序列相同(1 056 bp),仅5′-UTR和3′-UTR分别有1个变异碱基(T→G、T→C),将该核苷酸序列命名为 GluD3-18(GenBank登录号KY430289);在矮抗58中获得2种DNA序列,一种长度为1 043 bp,与 GluD3-42相似性高达99.1%,编码区序列相同(915 bp),仅在3′-UTR有9个变异碱基,命名为 GluD3-44(GenBank登录号:KY430290),另一个长度为1 055 bp,与 GluD3-21(GenBank登录号:DQ357054.1)的相似性为99.8%,编码区序列长度相同(927 bp),但有1个变异碱基(C→T),导致对应氨基酸T替换成I,将该基因命名为 GluD3-24(GenBank登录号:KY436383),并将推导的氨基酸序列命名为P-24;在偃展4110中扩增的核苷酸序列长度为1 055 bp,与 GluD3-21的相似性高达99.9%,编码区序列相同(927 bp),仅在5′-UTR区有1个变异碱基(G→A),命名为 GluD3-25(GenBank登录号:KY430291);而在周麦18、郑麦004和豫麦50中并未扩增出条带。
利用引物LB2F/LB2R在不同筋度的小麦品种中共扩增到9个条带,测序后得到5个新的序列变异类型(图2B)。其中,从郑麦366、郑麦9023、师栾02-1和扬麦13号中得到的核苷酸序列相同,命名为 GluD3-16(GenBank登录号:KY430292);从矮抗58和周麦18中得到的核苷酸序列相同,命名为 GluD3-17(GenBank登录号:KY430293)。 GluD3-16和 GluD3-17的核苷酸序列长度均为1 341 bp,编码区长度为1 056 bp,在第1 324 bp处有一个碱基变异( GluD3-16为C, GluD3-17为T)。 GluD3-16、 GluD3-17与 GluD3-11相似性分别为99.8%和99.7%,且与 GluD3-11的编码区序列相同。从偃展4110、郑麦004和豫麦50中各扩增出一个核苷酸序列,长度均为1 341 bp,编码区长度均为1 056 bp。这3条序列与GenBank中注册的KC716051.1序列的相似性分别为99.9%、99.7%和99.8%,与 GluD3-11的相似性分别为99.6%、99.6%和99.5%。分别将从偃展4110、郑麦004和豫麦50中所得到的3条核苷酸序列命名为 GluD3-13、 GluD3-14和 GluD3-15 (GenBank登录号分别为:KY436384、KY436385 和 KY436386),对应肽链分别命名为P-13、P-14和P-15;P-13、P-14和P-15与KC716051.1编码的氨基酸序列(KC716051.1和 GluD3-11编码的氨基酸序列相同)相比,其相似性分别为99.4%、99.7%和99.4%。
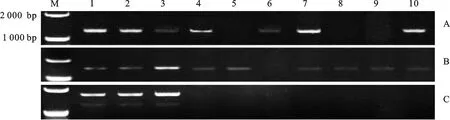
A:引物LB1F/LB1R的扩增产物;B:引物LB2F/LB2R的扩增产物;C:引物LB4F/LB4R的扩增产物。M:DL2000;1:郑麦366;2:郑麦9023;3:师栾02-1;4:矮抗58;5:周麦18;6:石家庄8号;7:偃展4110;8:郑麦004;9:豫麦50;10:扬麦13号。
A:PCR products amplified with the primer LB1F/LB1R; B:PCR products amplified with the primer LB2F/LB2R; C:PCR products amplified with the primer LB4F/LB4R. M:DL2000; 1:Zhengmai 366; 2:Zhengmai 9023; 3:Shiluan 02-1; 4:Aikang 58; 5:Zhoumai 18; 6:Shijiazhuang 8; 7:Yanzhan 4110; 8:Zhengmai 004; 9:Yumai 50; 10:Yangmai 13.
图23对Glu-B3位点基因特异性引物在普通小麦中的PCR扩增产物
Fig.2PCRproductsofGlu-B3amplifiedwiththreegene-specificprimersfromcommonwheat
利用引物LB4F/LB4R对不同筋度的10份小麦品种进行PCR扩增,结果(图2C)发现,仅在强筋小麦品种郑麦366、郑麦9023和师栾02-1中扩增出了PCR产物,其中从郑麦366和郑麦9023中扩增得到的是单元型 GluB3-41,从师栾02-1中得到的是单元型 GluB3-43,即引物LB4F/LB4R并未扩增到新的变异类型。
2.3Glu-A3位点和Glu-D3位点上新等位变异系统发育树的构建
为了明确本研究新发现的 Glu-A3基因和 Glu-D3基因与其他已知的 Glu-A3基因和 Glu-D3基因的进化关系,采用MEGA 4.0软件的NJ法和最大拟自然法对本研究发现的8个新等位变异( GluA3-25、 GluA3-26、 GluA3-37、 GluA3-38、 GluD3-24、 GluD3-13、 GluD3-14和 GluD3-15)与GenBank中搜索到的32个普通小麦 Glu-A3基因、6个小麦近缘种 Glu-A3基因、15个普通小麦 Glu-D3基因及17个小麦近缘种 Glu-D3基因的编码区序列进行了聚类分析。结果(图3)表明,所有序列划分为2个一级分支、5个二级分支和10个三级分支。在二级分支下, GluA3-37和 GluA3-38聚为一类, GluA3-25、 GluA3-26、 GluD3-13、 GluD3-14和 GluD3-15聚为另一类。在三级分支下, GluD3-13、 GluD3-14和 GluD3-15聚为一类, GluA3-25和 GluA3-26聚为另一类;而 GluD3-24在二级分支下就和其余的7个新等位变异分离,与山羊草中的编码LMW-GS的序列DQ287977相似度最高。以上结果说明, GluD3-13、 GluD3-14和 GluD3-15间及 GluA3-25和 GluA3-26间相似度较高。另外,在二级分支下, GluA3-25、 GluA3-26、 GluD3-13、 GluD3-14和 GluD3-15聚为一类,还说明 Glu-A3和 Glu-D3序列间具有较高的相似度。
3 讨 论
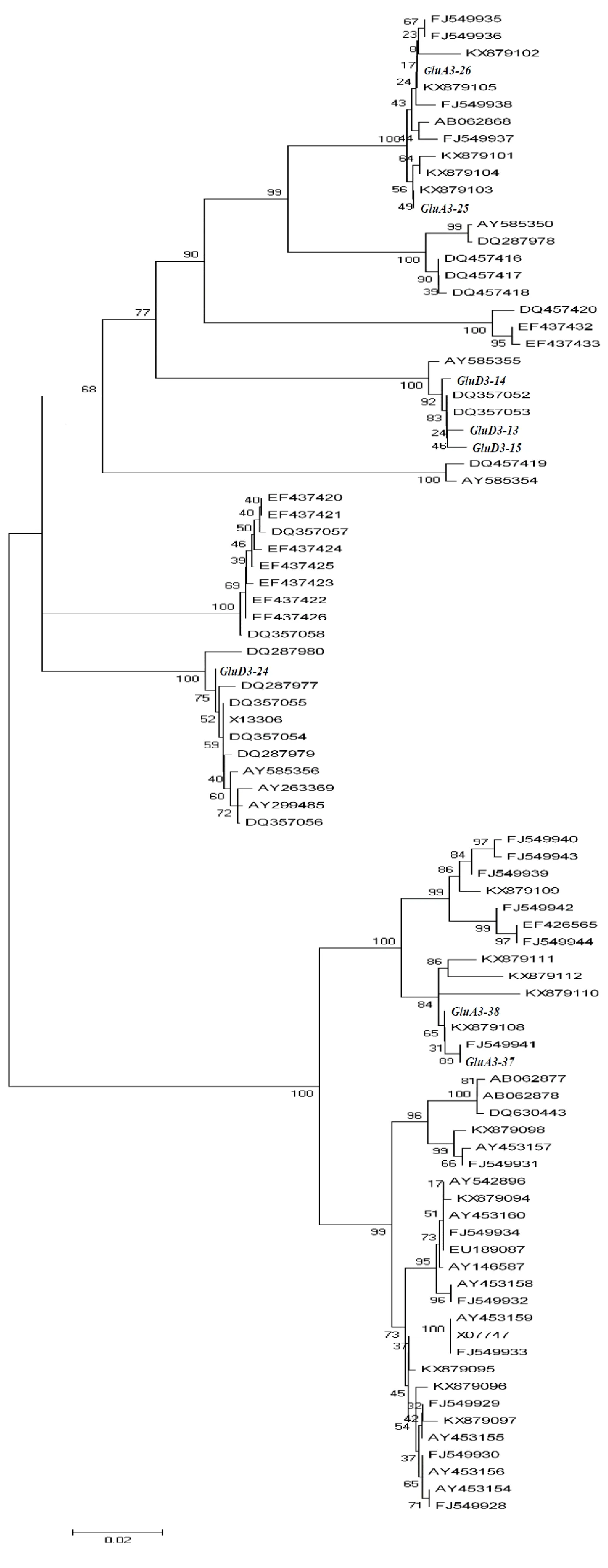
斜体为本研究所得到的在编码区有碱基突变的 Glu-A3和 Glu-D3新单元型。
Italic means the new Glu-A3 and Glu-D3 haplotypes with mutations in coding regions identified in this paper.
图3来源于普通小麦及其近缘种中的78个Glu-A3和Glu-D3基因的进化树分析
Fig.3Phylogenetictreeofseventy-eightGlu-A3andGlu-D3genesincommonwheatandrelatedspecies
LMW-GS基因核苷酸序列的长度主要是由重复区内重复序列的多少决定的,不同等位变异之间的差别则取决于核苷酸序列间存在的SNP和InDel数目。本研究利用 Glu-A3和 Glu-B3基因的特异引物在不同筋力小麦品种中共扩增到14个新的LMW-GS基因序列,但值得注意的是,在 Glu-D3位点上新发现的9个等位变异是由 Glu-B3基因的特异引物扩增的,其原因可能是 Glu-B3 和 Glu-D3位点的碱基序列具有较高的相似性。韩 冉等[3]对 Glu-A3、 Glu-B3和 Glu-D3位点上的LMW-GS基因进行聚类分析,同样发现 Glu-B3 和 Glu-D3位点间基因编码区序列具有较高的相似性, Glu-A3位点基因与 Glu-B3或 Glu-D3位点基因编码区序列差异相对较大。在新扩增到的14个新的LMW-GS基因序列中,在 Glu-A3位点得到4个变异类型,分别是 GluA3-2基因的单元型 GluA3-25和 GluA3-26,以及 GluA3-3基因的单元型 GluA3-37和 GluA3-38,但这4个单元型均为假基因。在 Glu-B3位点得到了1个新的碱基变异序列,但对应氨基酸序列是曾经报道过的肽链。在 Glu-D3位点,共发现了9个新的单元型,分别为 GluD3-18、 GluD3-44、 GluD3-25、 GluD3-16、 GluD3-17、 GluD3-13、 GluD3-14、 GluD3-15和 GluD3-24,其中, GluD3-18、 GluD3-44、 (GluD3-25、) GluD3-16和 GluD3-17在编码区与已经报道的相关基因核苷酸序列无差异,但在5′-UTR或3′-UTR区有1至9个碱基变异。有研究认为,UTR区的等位变异位点对基因的转录和翻译具有非常重要的影响,包括mRNA的半衰期、转录本的剪切、转录本的表达水平和蛋白质的翻译等[21-22]。而 GluD3-13、 GluD3-14、 GluD3-15和 GluD3-24单元型对应的肽链P-13、P-14、P-15和 P-24为本次新发现的肽链变异类型。在普通小麦中发现的LMW-GS基因与本人先前在小麦近缘种中发现的LMW-GS基因相比,普通小麦的LMW-GS基因相对稳定,而在小麦近缘种中的LMW-GS基因多样性更为广泛,是小麦品质遗传改良不可忽视的基因资源。
根据N-末端起始氨基酸的不同,分别为甲硫氨酸、丝氨酸和异亮氨酸,又可将低分子量麦谷蛋白亚基分为LMW-m、LMW-s和LMW-i 3种类型[23-24]。据此可以确定,本研究获得的氨基酸序列AP1和AP3的N-末端为ISQQQQ-,属于LMW-i类型;AP2的N-末端为 MDTSYIP-和MDTSCIP-,属于LMW-m类型;P-1和P-2的N-末端为METSRVP-和 METRCIP-,也属于 LMW-m类型。由于在 GluA-3位点上新发现的4个单元型均为假基因,在理论上不能编码蛋白,而在 GluD-3位点上新发现的4个单元型均含有完整的开放阅读框,编码的肽链P-13、P-14、P-15 和P-24的分子量依次约为39.83、39.81、39.81和34.91 kDa,这与王林海等的研究结果基本一致[18,25]。
根据多数人对LMW-GS基因的研究结果, GluA3-3基因的等位变异类型大多数为假基因[26],这在本研中也得到了验证,一共发现了2个 Glu-A3-3等位变异,都为假基因。由于3个 Glu-3位点的LMW-GS基因之间有着极高的相似性,特异引物的特异性受到限制。日本的Ikeda等[27]曾用1对引物扩增30多个 Glu-3基因。在本研究中,原本用于扩增 Glu-B3基因的引物,从10份小麦材料中分别扩增出 Glu-B3位点的 GluB3-418以及 Glu-D3位点的 GluD3-18、 GluD3-44、 GluD3-25、 GluD3-16、 GluD3-17、 GluD3-13、 GluD3-14、 GluD3-15和 GluD3-24基因序列。在 Glu-D3位点上扩增到9个新的等位变异,这说明 Glu-D3基因与 Glu-B3基因的核苷酸序列有着较高的相似性。其中,在偃展4110小麦品种中扩增到的 GluD3-13对应的肽链P-13含有7个半胱氨酸,其缺少的氨基酸位于C末端保守区上的第8个半胱氨酸的位置上。从师栾02-1小麦品种中扩增到一个含有9个半胱氨酸残基的单元型,是首次在小麦品种中发现,但与本人曾在栽培二粒小麦中得到的 GluA3-18单元型相同,进一步说明一粒小麦就是普通小麦的祖先[28]。
大多数LMW-GS均含有8个半胱氨酸残基,在第1和第7位上的半胱氨酸残基形成分子间二硫键,其余6个形成分子内二硫键[29]。AP1-8是在第6位和第7位半胱氨酸残基之间多出一个半胱氨酸残基。Zhao等[19]曾在小偃6号小麦品种的1D染色体上首次发现含有9个半胱氨酸残基的LMW-GS,其多出的半胱氨酸残基在第3和第4位半胱氨酸残基之间;Si等[5]在1B染色体上也发现了含有9个半胱氨酸残基的LMW-GS,其多出的半胱氨酸残基在第7和第8位半胱氨酸残基之间。根据Zhao等[30]和Xu等[31]的报道,与含有8个半胱氨酸残基LMW-GS材料的面粉相比,含有9个半胱氨酸残基的LMW-GS更有助于形成分子间二硫键,进而提高了和面时间,显著增强了面团强度,增多了蛋白聚合体,减弱了抗延阻力。此外,一些含有7个半胱氨酸的LMW-GS相继被发现,例如Lan等[32]曾在小麦品种Jiachazharen、Jiachaaigan和Maoyintumai中分别发现了含有7个半胱氨酸的LMW-GS,韩 冉等[3]在从小麦品种Cook中也发现了含有7个半胱氨酸的LMW-GS。经对本研究获得的含有7个半胱氨酸残基的LMW-GS进行二级结构预测,发现 P-13蛋白二级结构中α-螺旋和β-折叠所占的比例之和为24.86%,这明显高于 GluD3-1基因的其他等位变异(24.00%~24.57%),这种二级结构的变化可能与缺少一个半胱氨酸残基有关,半胱氨酸残基数量的变化会影响LMW-GS分子间二硫键的形成,从而影响到其三级结构的构型,进而会影响到蛋白的功能及面粉品质。据此推测, GluA3-18和 GluD3-13可能是小麦改良的优质基因资源。另外值得注意的是,本研究用了6对特异引物对不同筋力的10份小麦材料进行LMW-GS基因扩增,其中1对引物LB4F/LB4R只在三个强筋型小麦品种中扩增出了条带,其他7个品种没能扩增出PCR产物,说明LB4F/LB4R有可能对强筋小麦的LMW-GS基因具有特异性,这有待进一步研究。
[1]VERAVERBEKEW S,DELCOURJ A.Wheat protein composition and properties of wheat glutenin in relation to breadmaking functionality [J].CriticalReviewsinFoodScienceandNutrition,2002,42(3):184.
[2]LUO G B,ZHANG X F,ZHANG Y L,etal.Composition,variation,expression and evolution of low-molecular-weight glutenin subunit genes inTriticumurartu[J].BMCPlantBiology,2015,15(1):7.
[3] 韩 冉,马 猛,魏燕燕,等.优质小麦品种 Glu-A3位点LMW-GS基因的克隆及分子特征[J].中国农业科学,2010,43(5):884.
HAN R,MA M,WEI Y Y,etal.Cloning and characterization of LMW-GS gene at Glu-A3 complex locus in different wheat cultivars [J].ScientiaAgriculturaSinica,2010,43(5):884.
[4]WANG Y P,ZHEN S M,LUO N N,etal.Low molecular weight glutenin subunit gene Glu-B3h confers superior dough strength and breadmaking quality in wheat (TriticumaestivumL.) [J].ScientificReports,2016,6:27182.
[5]SI H Q,ZHAO M L,ZHANG X,etal.Cloning and characterization of low-molecular-weight glutenin subunit alleles from Chinese wheat landraces (TriticumaestivumL.) [J].ScientificWorldJournal,2014,2014(1):371045.
[6] 程西永,吴少辉,李海霞,等.小麦高、低分子量麦谷蛋白亚基对品质性状的影响[J].麦类作物学报,2014,34(4):483.
CHENG X Y,WU S H,LI H X,etal.Effects of HMW and LMW glutenin subunits on wheat quality traits [J].JournalofTriticeaeCrops,2014,34(4):483.
[7]ITO M,FUSHIE S,FUNATSUKI W M,etal.Effect of allelic variation in three glutenin loci on dough properties and bread-making qualities of winter wheat [J].BreedingScience,2011,61(3):285.
[8]JIN H,ZHANG Y,LI G,etal.Effects of allelic variation of HMW-GS and LMW-GS on mixograph properties and Chinese noodle and steamed bread qualities in a set of Aroona near-isogenic wheat lines [J].JournalofCerealScience,2013,57(1):150.
[9]ZHANG X,JIN H,ZHANG Y,etal.Composition and functional analysis of low-molecular-weight glutenin alleles with Aroona near-isogenic lines of bread wheat [J].BMCPlantBiology,2011,12(1):14.
[10]ZHEN S M,HAN C X,MA C Y,etal.Deletion of the low-molecular-weight glutenin subunit allele Glu-A3a of wheat (TriticumaestivumL.) significantly reduces dough strength and breadmaking quality [J].BMCPlantBiology,2014,14(1):13.
[11]SREERAMULU G,SINGH N K.Genetic and biochemical characterization of novel low molecular weight glutenin subunits in wheat [J].GenomeBiology,1997,40(1):416.
[12]LIU C Y,SHEPHERD K W.Inheritance of B subunits of glutenin and ω-and γ-gliadins in tetraploid wheats [J].TheoreticalandAppliedGenetics,1995,90(7):1154.
[13]D′OVIDIO R,MASCI S.The low-molecular-weight glutenin subunits of wheat gluten [J].JournalofCerealScience,2004,39(3):336.
[14]ANDERSEN J R,LBBERSTEDT T.Functional markers in plants [J].TrendsinPlantScience,2003,8(11):558.
[15]ZHANG W,GIANIBELLI M C,RAMPLING L R,etal.Characterisation and marker development for low molecular weight glutenin genes from Glu-A3 alleles of bread wheat (TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2004,108(7):1412.
[16] 孙学永,马传喜,司红起,等.中国小麦微核心种质低分子量麦谷蛋白 Glu-A3位点等位基因的PCR检测[J].分子植物育种,2006,4(4):480.
SUN X Y,MA C X,SI H Q,etal.Screening Glu-A3 alleles of LMW-GS genes from mini-core collections of Chinese wheat varieties using alleles-specific PCR markers [J].MolecularPlantBreeding,2006,4(4):480.
[17] 赵曼丽.低分子量麦谷蛋白亚基基因的克隆及序列分析[D].合肥:安徽农业大学,2014:13.
ZHAO M L.Cloning and sequence analysis of low molecular weight glutein subunit gene [D].Hefei:Anhui Agricultural University,2014:13.
[18] 王林海.普通小麦及其近缘种低分子量麦谷蛋白基因克隆与STS标记开发[D].北京:中国农业科学院,2009:13-24.
WANG L H.Characterisation of low-molecular-weight glutenin genes and development of STS markers in common wheat and its relatives [D].Beijing:Chinese Academy of Agricultural Sciences,2009:13-24.
[19]ZHAO X L,XIA X C,HE Z H,etal.Characterization of three low-molecular-weight Glu-D3 subunit genes in common wheat [J].TheoreticalandApplliedGenetics,2006,113(7):1247.
[20] 刘进英,董 雪,冯玉梅,等.不同休眠特性小麦中TaGAMyb-B基因的等位变异及其表达特性[J].麦类作物学报,2016,36(6):709.
LIU J Y,DONG X,FENG Y M,etal.Identification and expression ofTaGAMyb-Bin wheat with different depth of dormancy [J].JournalofTriticeaeCrops,2016,36(6):709.
[21]BAEV V,DASKALOVA E,MINKOV I.Computational identification of novel micro RNA homologs in the chimpanzee genome [J].ComputationalBiology&Chemistry,2009,33(1):67.
[22] 杨具田,徐红伟,臧荣鑫,等.五个地方绵羊品种 FAS基因 3′-UTR区单核苷酸多态性研究[J].中国农业科学,2010,43(13):2790.
YANG J T,XU H W,ZANG R X,etal.Polymorphism of 3′-UTR of fatty acid synthase (FAS) gene in five chinese sheep breeds [J].ScientiaAgriculturaSinica,2010,43(13):2790.
[23]QIN L M,LIANG Y,YANG D Z,etal.Novel LMW glutenin subunit genes from wild emmer wheat(Triticumturgidumssp.dicoccoides) in relation to Glu-3 evolution [J].DevelopmentGenesandEvolution,2015,225(1):315.
[24]LONG H,WEI Y M,YAN Z H,etal.Classification of wheat low-molecular-weight glutenin subunit genes and its chromosome assignment by developing LMW-GS group-specific primers [J].TheoreticalandAppliedGenetics,2005,111(7):1256.
[25]WANG L H,ZHAO X L,HE Z H,etal.Characterization of low-molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat (TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2009,118(3):533.
[26] 董 雪,赵献林,刘 梦,等.普通小麦近缘种低分子量麦谷蛋白亚基 Glu-A3基因的分离和鉴定[J].作物学报,2017,43(6):831.
DONG X,ZHAO X L,LIU M,etal.Isolation and characterization of LMW-GS Glu-A3 in common wheat related species [J].ActaAgronomicaSinica,2017,43(6):831.
[27]IKEDA T M,NAGAMINE T,FUKUOKA H,etal.Characterization of new low-molecular-weight glutenin subunit genes in wheat [J].TheoreticalandAppliedGenetics,2002,104(4):685.
[28]STEPIEN L,HOLUBEC V,CHELKOWSKI J.Resistance genes in wild accessions of triticeae-inoculation test and STS marker analyses [J].JournalofAppliedGenetics,2002,43(4):431.
[29] 虎梦霞.小麦低分子量麦谷蛋白亚基(LMW-GS)研究进展[J].甘肃农业科技,2014(1):43.
HU M X.Research progress on low molecular weight glutenin subunits (LMW-GS) in wheat [J].GansuAgriculturalScienceandTechnology,2014(1):43.
[30]ZHAO H X,WANG R J,GUO A G,etal.Development of primers specific for LMW-GS genes located on chromosome 1D and molecular characterization of a gene from Glu-D3 complex locus in breed wheat [J].Hereditas,2004,141(3):19.
[31]XU H,WANG R J,SHEN X,etal.Functional properties of a new low-molecular-weight glutenin subunit gene from a bread wheat cultivar [J].TheoreticalandAppliedGenetics,2006,113(7):1299.
[32]LAN Q X,FENG B,XU Z B,etal.Molecular cloning and characterization of five novel low molecular weight glutenin subunit genes from Tibetan wheat landraces (TriticumaestivumL.) [J].GeneticResourcesandCropEvolution,2013,60(2):80.
IsolationandSequenceAnalysisofLMW-GSGeneattheGlu-3LociinCommonWheat
DONGXue1,WANGZeng1,LIUMeng1,YANGYan1,HANBing1,LIShufen1,ZHAOXianlin2
(1.The Functional Laboratory of Plant Biotechnology,College of Life Sciences,Inner Mongolia Agricultural University,Huhhot,Inner Mongolia 010018,China; 2.Wheat Research Institute,Henan Academy of Agricultural Sciences/Henan Key Laboratory of Wheat Biology/National Engineering Laboratory for Wheat/Key Laboratory of Wheat Biology and Genetic Breeding in Central Huang-Huai Region,Ministry of Agriculture,Zhengzhou,Henan 450002,China)
Low-molecular-weight glutenin subunits(LMW-GS) are major fractions of wheat gluten,and have great effects on dough extensibility and food processing quality. In this study,the specific primers of Glu-A3 and Glu-B3 genes were used to identify and amplify the allelic variations of LMW-GS genes from ten wheat varieties with different gluten content. A total of 14 new DNA haplotypes were identified,including four haplotypes at Glu-A3,one haplotype at Glu-B3 and nine haplotypes at (Glu-D3.) Among them,the corresponding peptide of GluA3-18 from wheat variety Shiluan 02-1 contains nine cysteine residues,and the corresponding peptide of GluD3-13 from Yanzhan 4110 has seven cysteine residues,both of which belong to the specific subunits compared with others with eight cysteine residues. The results provide the basis for further study of LMW-GS genes and quality improvement of wheat.
Common wheat; LMW-GS; Gene isolation; Glu-3 loci
时间:2017-10-11
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20171011.1601.002.html
2017-02-13
2017-07-25
国家自然科学基金项目(31271726);内蒙古农业大学优秀青年科学基金项目(2014XYQ-18);内蒙古自治区自然科学基金项目(2014MS0338);内蒙古自治区产业创新创业人才团队项目(201503004)
E-mail:dongxue225@126.com
赵献林(E-mail:xlin918@126.com);杨 燕(E-mail:yangyanchutao@126.com)
S512.1;S330
A
1009-1041(2017)10-1276-09
网络出版时间:2017-10-11
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20171011.1601.004.html
猜你喜欢
作物学报(2022年6期)2022-04-08
麦类作物学报(2022年1期)2022-03-01
国际医学放射学杂志(2021年5期)2021-10-22
中华养生保健(2020年5期)2020-11-16
农民致富之友(2019年33期)2019-12-20
河南农业(2018年12期)2018-12-05
麦类作物学报(2018年4期)2018-05-11
中国医药指南(2017年3期)2017-11-13
第一财经(2017年36期)2017-09-25
