
鱼类粒径谱研究进展
2017-11-07 02:34:58郭建忠陈作志徐姗楠
海洋渔业 2017年5期
郭建忠,陈作志,徐姗楠
鱼类粒径谱研究进展
郭建忠1,2,陈作志1,徐姗楠1
(1.中国水产科学研究院南海水产研究所,农业部南海渔业资源开发利用重点实验室,广东省渔业生态环境重点实验室,广东广州 510300;2.上海海洋大学水产与生命学院,上海 201306)
粒径谱表示的是生物量或者生物数量与粒径大小关系的曲线,能够反映生态系统结构和功能以及系统内部的动态联系,是当前水生生态学研究的热点。综述了鱼类粒径谱的理论、研究方法及影响因子等,重点介绍了鱼类粒径谱在鱼类生产量评估,开展生态系统监测,以及预测各种人类活动对生态系统的影响等方面的研究进展。针对目前研究中所存在的不足与问题,提出了新观测和采样技术的应用以及基于粒径谱评估鱼类生物量的通用理论模式研发等是今后鱼类粒径谱研究的趋势。
粒径谱;鱼类粒径谱;生态系统
粒径谱表示的是生物量或者生物数量与粒径大小关系的曲线,是研究生态系统特征的一个重要途径。它不仅能够从宏观上研究海洋生态系统的结构、动态和影响机制,而且能够比较不同生态系统的特征、评估生物群落的生产力与生物量,对生态系统的研究起着重要的指示作用[1]。在水生生态系统中[2],鱼类粒径谱被广泛应用于评估不同营养水平的生产状况,尤其是鱼类的生产、丰度和物种的丰度[3-9],以及预测各种人类活动对生态系统的影响等[10-11]。因此,研究鱼类粒径谱对于渔业的管理、开发和生态系统的可持续健康发展有着重要的意义。
1 粒径谱
1.1 粒径谱概念及发展
1967年 SHELDON等[12-13]首次提出粒径谱的概念,即粒径谱是在相等的对数粒径间隔上测量生物体总的丰度或生物量;将某一特定的生态系统中的生物按照统一颗粒粒径大小(equivalent spherical diameter,ESD)划分为不同的粒径范围,计算各粒径范围上的生物量,并将其对数转化,那么这种生物量在对数粒径范围上的分布称为生物量粒径谱。之后又提出了粒径谱理论,即相等对数化粒径间隔的颗粒物质的生物量大致相同。之后的40年间,该理论得到了迅速的发展[14]。20世纪 70年代,SHELDON等[8]以捕食者-被捕食者相互作用关系建立模型来研究海洋生态系统;KERR[15]在 SHELDON模型基础上以能量流动与营养级关系来构建模型;THIEBAUX等[16-17]以双对数坐标绘出非正态化生物量谱,并提出正态化生物量的概念[13];SILVERT等[18]以时间为基础建立模型。80年代,BORGMANN[3]提出粒径转换效率“ε”概念,并用来评估生物体粒径组的生产力[10]。90年代,BOUDREAU等[19]用生态动力学理论来分析生物量谱穹顶与营养级、粒径的关系;THIEBAUX等[16]在P-D模型基础上建立完全异速生长模型;ZHOU等[20]建立正态化生物量谱来平衡生物体出生率、生长率、死亡率与捕食的关系。21世纪初期,ROSEN[21]建立粒径模型来预测任意粒级范围内不同粒径的物种比例;LAW等[22]以特定捕食者粒径为基础建立参数化模型;ROSSBERG[23]通过摄食、新陈代谢和生长的能量消耗等关系将不同规格的性成熟鱼类物种连接起来建立模型。
1.2 粒径谱研究方法
对于水生生态系统,粒径谱有两种主要的研究方法[24]。第一种是假定生物体由小型生长到大型的生物量是连续的,集中分析生物量粒径谱的斜率,采用动态方程式描述水生生物群落特征[25]。第二种是在以粒径为基础的生态系统结构一致的前提下分析生物量粒径谱的不规则性和分布结构[26]。两种方法形成对照,第一种强调生长,第二种强调捕食行为,且第二种已经广泛用于分析粒径谱形成的“峰型”曲线上穹顶形成的原因和研究海洋底栖无脊椎动物、鱼类粒径谱的斜率[27]。
2 鱼类粒径谱
2.1 鱼类粒径谱基本原理
1991年,SPRULES等[9]对美国密歇根湖海洋生物进行研究,首次完成了生物量粒径谱从细菌到大型鱼类的应用。自此,国内外对于鱼类粒径谱的研究逐步开展起来。鱼类粒径谱曲线所形成的“峰型”形状反映鱼类群落的结构特征,如图1安大略湖鱼类粒径谱呈现“单峰”型。正态化鱼类粒径谱表面上反映鱼类群落生物量的转化分布,而本质上代表鱼类群落的粒级丰度;方法上强调异速生长比例,其理论斜率为-1,且斜率与海域的生产状况、鱼类粒径大小、捕捞强度、栖息地环境等因素有关,表示生物量与粒径之间的关系,如图1安大略湖鱼类正态化粒径谱斜率为-1.249 8,小于-1,说明生物量随粒径的增大而减小。
2.2 鱼类粒径谱类型
2.2.1 SHELDON型生物量粒径谱
SHELDON型生物量粒径谱是用以2为底对数转换的个体干重生物量划分的粒径间隔为横坐标,用以2为底对数转换的单位面积上对应粒径间隔的干重生物量为纵坐标[26]。SHELDON型生物量粒径谱曲线上穹顶与波谷构成的“峰型”能够反映不同粒径范围内鱼类群落的结构特征。
2.2.2 正态化生物量粒径谱
正态化生物量粒径谱(normalized biomass size spectrum,NBSS)是用以2为底对数转换的个体干重生物量划分的粒级为横坐标,用以2为底对数转换后的单位面积上对应粒级总干重生物量除以该粒级上个体干重的变化幅度为纵坐标[28]。正态化生物量粒径谱包含几个重要参数信息:斜率(a)、截距(b)和回归系数(R2)。
正态化生物量粒径谱参数能够描述鱼类群落的结构特征、比较不同鱼类群落间的差异和预测鱼类群落未来的发展状况[13]。a表示鱼类群落生物量与粒径之间的关系:a=-1,鱼类群落生物量随着粒径的增大而保持为一个不变的常数;a<-1,鱼类群落生物量随着粒径的增大而减小;a>-1,鱼类群落生物量随着粒径的增大而增大。b表示鱼类群落的丰度和生产力水平,b值不同则代表不同鱼类群落生产力水平之间的差异[26];一般而言,生产力水平的大小与b值的大小呈正相关[29]。R2表示鱼类群落偏离稳定状态的程度。
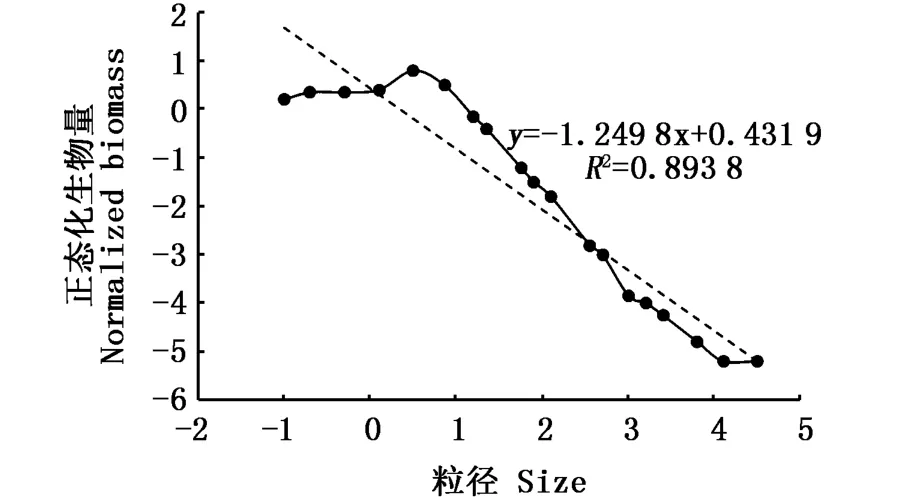
图1 安大略湖鱼类生物量谱,虚线为正态化生物量谱线(线性项)[13]Fig.1 Fish biomass size spectrum for Lake Ontario with fitted parabolas and the linear“backbone”[13]
2.3 鱼类粒径谱主要研究方法
2.3.1 库尔特计数法
1967年库尔特发明计数器以后,库尔特计数法开始应用于海洋生物粒径谱的研究[14],根据小孔电阻原理来测量颗粒大小。通过测量每个颗粒大小,统计出粒径分布,是最早研究海洋生态系统粒径的一种方法。
2.3.2 拖网捕捞法
对于鱼类群落的研究一般是海上现场调查,根据所需鱼种类型选择网型和网目直径,通过拖网捕捞获取鱼类数据开展研究。该方法较为直观,但也具有一定的局限性。如网目过大会导致小型鱼类遗漏,网目过小会导致过度捕捞,都不利于鱼类群落的研究和鱼类资源合理开发。MACPHERSON等[30]用拖网调查对地中海西北部鱼类群落研究表明,受保护海域,鱼类群落粒径结构较平稳,生物粒径谱斜率较平缓;未受保护海域,鱼类群落粒径结构急剧变化,生物粒径谱斜率较陡峭。
2.3.3 粒径分选法
1983年,SHIRAYAMA[31]最早将此方法用于海洋底栖生物粒径结构的研究,之后逐步应用于鱼类的研究。该方法主要是根据鱼类大小来选取网目直径,获得不同粒级范围内鱼类物种的生物量与丰度状况,是研究鱼类粒径谱最常用的方法。
2.3.4 异速生长 P∶B比率法
异速生长P∶B比率法即鱼类生产量与生物量之间比率(production:biomass,P∶B),最早由BANSE等[32]提出被广泛应用于评估鱼类群落的生产力。JUNG等[33]于1995~2000年对切萨皮克海湾鱼类群落研究,用P∶B比率来评估鱼类群落的生产力状况,并提出公式:P(w)=β(w)×100.44-0.26lg(w)。其中,P(w)代表粒级 w的年际生产量;β(w)代表粒级w的年均现存生物量;w单位是kcal,1 g湿重=1 269 kcal;年际总鱼类生产量 P=ΣP(w)。
2.3.5 PPMR和 TE
平均捕食者规格与被捕食者规格比值(the ratio of the mean predator size to mean prey size,PPMR)和营养转化效率(trophic transfer efficiency,TE)对于鱼类群落丰度-质量关系粒径谱的斜率有显著的约束性,且二者在不同的生态系统中具有显著的一致性[34]。CYR[35]和BROWN等[36]指出,在一个完整的粒径谱中,PPMR和TE可以预测不同粒径鱼类物种的能量变化和斜率变化。JENNINGS等[37]为评估捕捞对北海鱼类群落的影响,计算PPMR和TE来预测未开发海域鱼类生物粒径谱的斜率,与现有数据估算的丰度-质量关系进行比较,得出未开发海域到开发海域的鱼类生物量随生物体质量的缩放范围为 M-0.1~M-1.0(M:body mass)。
2.3.6 生化分析法
用生物、化学原理和方法来研究鱼类粒径谱。主要包括化合物分析、元素分析(常量元素、微量元素[38]、同位素)、C/N比值[39](主要是脂类与蛋白质的组成成分)和能量分析等方法。CHANDLER等[40]于1998年利用稳定同位素方法(δ15N)对美国安大略湖东北部湖泊鱼类群落研究表明:鱼类群落营养结构呈现“金字塔”模式[41]。
2.4 影响鱼类粒径谱的主要因素
2.4.1 生产力
鱼类正态化生物粒径谱的斜率与海域生产力相关[42]。生产力高的海域(上升流中心),最小粒级的丰度非常高,鱼类群落生物量随着粒径的增大而明显降低。生产力低的海域,鱼类群落生物量随着粒径的增加而增加或保持为一个不变的常数,且鱼类粒径谱的斜率往往较低[42]。EMMRICH等[43]于2011年对德国北部湖泊鱼类群落研究表明,生产力低的湖泊,大型食肉鱼类的丰度较高,鱼类生物粒径谱的斜率在水体深处比较平缓。
2.4.2 栖息地环境
栖息地环境影响鱼类的生长,导致鱼类生物粒径谱上各粒级范围内鱼类物种结构的不同。MACPHERSON等[30]于1997~1998年对地中海西北部沿海区域鱼类群落研究表明,岩石基底海域,鱼类群落生物量随着粒径的增大而保持为一个不变的常数。海草床基底海域,保护区鱼类群落生物量随着粒径的增大而增大;未保护区鱼类群落生物量随着粒径的增大而保持为一个不变的常数。
2.4.3 捕捞强度
捕捞强度影响鱼类的粒径结构与丰度[44],导致鱼类群落平均粒径下降和大型鱼类比例减少,且鱼类生物粒径谱的斜率随着捕捞强度的增大而更加陡峭。MACPHERSON等[30]对地中海西北部鱼类群落研究表明,未受捕捞海域,鱼类群落粒径结构较平稳且粒径谱斜率较平缓;受捕捞海域,鱼类群落粒径结构急剧变化且粒径谱斜率较陡峭。ZWANENBURG[45]也表明,与捕捞强度低的鱼类群落相比,捕捞强度高的鱼类群落粒径谱的斜率往往比较陡峭。
2.4.4 竞争
在生态系统食物网中,生物之间存在捕食、竞争、捕食和竞争三种关系。竞争影响鱼类群落的粒径结构。ROSEN[21]研究表明,正是由于生态系统中存在竞争关系,群落中大型物种生物量减少促使小型物种生物量的增殖,从而导致鱼类生物粒径谱中任意粒级范围内不同粒径物种比例的差异。
2.4.5 营养级水平
强大的粒径结构在海洋生态系统中是普遍存在的[7],且营养水平与生物体粒径大小有关[46]。在大多数水生生态群落中,营养级水平随着生物体粒径的增大而不断地上升[47]。PAHL[48]于1993年对生态系统食物网时空格局研究表明,食物网中营养级水平不仅可以比较不同生态系统的结构特征,而且可以作为预测食物网内部动态结构的指标。JENNINGS等[47]于2002年对英国北海鱼类群落研究表明,在不考虑物种差异前提下,鱼类生物体粒径大小与其对应的营养级水平呈显著性相关;在相似的鱼类群落中,营养级水平与生物体质量呈线性关系,且这种线性关系可用来预测鱼类群落的营养级结构特征。
2.4.6 渔具选择
渔业具有选择性,往往体型大的、营养价值高的鱼类物种是消费者所热衷的,因此需要根据所需鱼类物种来选择渔具,常见的主要渔具有底拖网、流刺网和张网[49]等,同时渔具的物理影响会导致鱼类选择性死亡。JENNINGS等[47]于2002年对英国北海鱼类群落研究表明,当拖网拖过海床时,脆弱的大型鱼类物种可能被捕获,而小型鱼类物种可能被渔具前面的压力波推到一边,从而导致拖网海域底层的群落结构由小型鱼类物种组成,且粒径谱的斜率在拖网严重的海域较大。
2.4.7 季节变化
季节变化是影响鱼类群落粒径结构的主要因子,不同的季节,鱼类群落的粒径结构组成不同。当鱼类物种由小型向大型转变时、鱼类群落动态变化跨越大的湖泊和海洋时,可以通过季节变化来研究给定鱼类群落粒径结构的时空差异。SPRULES等[9]于1991年对美国密歇根湖鱼类群落研究表明,鱼类群落粒径结构在季节上存在显著的差异性。
2.4.8 补偿比率
种群间鱼类物种的补偿比率影响鱼类群落的粒径结构,且年际补偿中大的波动影响鱼类的密度,导致鱼类群落生物量与粒径结构的变化[50]。补偿比率大,鱼类群落的粒径结构就相对完整;补偿比率小,鱼类群落的粒径结构就相对混乱。JENNINGS等[50]于2007年对海洋生物粒径开发和保护的研究中指出,鱼类群落密度依赖于调控,产卵-补充关系中现存的补偿比率对鱼类生物粒径谱的斜率起着关键的机制作用。
2.5 鱼类粒径谱的应用
2.5.1 环境扰动监测
通过鱼类正态化粒径谱斜率可以捕获到人为和自然干扰破坏生态的过程。DUPLISEA等[51]用鱼类正态化粒径谱参数来说明自然扰动对加拿大司考田沙洲深层鱼类的影响。BLANCHARD等[52]以北海4个鱼类种群为例建立粒径谱模型来评估种群和群落对捕捞的响应,进行环境监测。GRAHAM等[53]用鱼类粒径谱来研究捕捞对珊瑚礁鱼类的影响,得出结论,鱼类粒径谱的斜率随着捕捞强度的增加而逐步减小,且在高捕捞强度海域斜率变得显著增加;鱼类粒径谱的中点高度(粒径间隔中点值所对应的y轴生物量值)随着捕捞强度的增加而显著地下降。在粒级范围内,鱼类群落的丰度与捕捞强度呈负相关性。在捕捞海域,大型鱼类丰度的下降速度比小型鱼类丰度的下降速度更快(图2、图3)。同时鱼类粒径谱可以用来监测水体的营养状态。如:SPRULES等[28]研究表明,在富营养化生态系统中,鱼类生物粒径谱的斜率较小、截距较高或曲率较大、峰值较高。EMMRICH等[43]对德国北部湖泊鱼类群落研究表明:在生产力较低的湖泊,大型食肉鱼类丰度较高,鱼类生物粒径谱的斜率在水体深处较小。
2.5.2 反映鱼类群落特征
鱼类粒径谱曲线上穹顶与波谷所形成的“峰型”能够反映鱼类群落的结构特征。JUNG等[33]对切萨皮克海湾鱼类群落研究得出,鱼类生物粒径谱呈现“双峰”型,且第一峰穹顶对应粒径物种结构由小型食浮游动物的鱼类组成,主要是鳀鱼(Engraulis japonicus);第二峰穹顶对应粒径物种结构由大型杂食性的鱼类组成。季节上,第一峰穹顶对应的鱼类生物量水平和生物个体平均粒径从4月到10月呈逐步增加趋势;第二峰穹顶对应的鱼类生物量水平相对稳定。地域上,上、下海湾的第二峰穹顶的生物量水平高于第一峰穹顶,但当季节性和间接性缺氧同时发生时,中海湾区的第一、二峰穹顶的生物量水平最小(图3)。同时,在水生生态系统中,鱼类粒径谱能够反映鱼类群落的生态过程、生物量分布和能量流动[54]。重要的生态作用和生理过程都依赖于生物体粒径的大小,如:死亡率、转换效率、新陈代谢和 生 长 等[24]。PETERSON等[55]在 SILVERT等[18]基础上以粒径为基础建立粒径模型来估算鱼类群落的死亡率。
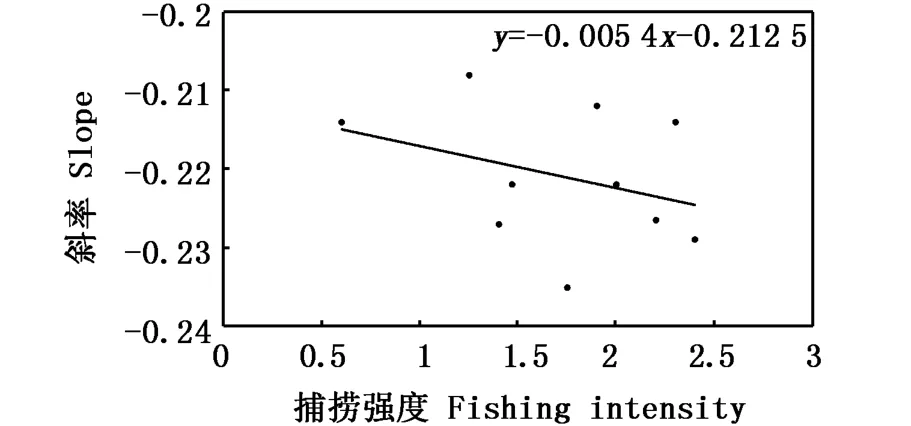
图2 捕捞强度与粒径谱斜率关系[53]Fig.2 Relationship between slope and fishing intensity[53]

图3 捕捞强度与中点高度关系[53]Fig.3 Relationship between mid-point height and fishing intensity[53]
2.5.3 预测鱼类生物量和生产量
BORGMANN[3]和 THIEBAUX等[16-17]用捕食者-被捕食者异速生长关系建立模型来评估不同营养组的生物量和生产量。BORGMANN认为,如果一个粒径组的生产量或生物量、粒径转换效率ε、以粒径为依赖的指数流动率n是已知的,那么任意粒径组生物体的粒径范围、生物量和生产量都可以被预测。SPRULES等[9]于1991年用已公布的ε和n值来预测美国密歇根湖浮游植物、浮游动物和鱼类的生物量,且与实际观测密切符合。THYGESEN等[56]用水生生物群落丰度粒径谱模型来计算给定粒径的鱼类的生物量大小。

图4 1995~2000年切萨皮克海湾中层拖网收集的鱼类平均正态化生物量粒径谱,由第一穹顶的优势种和第二穹顶物种杂食性鱼类组成(虚线为线性项)[33]Fig.4 Mean normalized biomass size spectrum for fish species,the dominant species in the first dome and the piscivorous fishes component of the second dome collected by mid-water trawl in Chesapeake Bay from 1995 to 2000(dotted line is linear term)[33]
2.5.4 反映生态系统的结构和功能
鱼类正态化粒径谱参数能够反映生态系统的结构和功能[18]。MACPHERSON等[30]对地中海西北部鱼类群落研究表明,受保护海域,斜率均大于-1,则鱼类生物量随着粒径的增大而增大;大的破碎岩石基底海域的截距大于海藻床基底海域的截距,则其鱼类群落生产力水平高于海藻床基底海域的生产力水平。未保护海域,轻微倾斜岩石基底海域的斜率小于-1,则鱼类生物量随着粒径的增大而减小;海藻床基底海域的斜率大于-1,则鱼类生物量随着粒径的增大而增大;轻微倾斜岩石基底海域的截距大于海藻床基底海域的截距,则其鱼类群落的生产力水平高于海藻床基底海域的生产力水平。R2值不同,则偏离群落稳定状态的程度有所差异,如表1。

表1 地中海西北部沿海保护海域与未保护海域鱼类群落NBSS参数值[30]Tab.1 Statistics for normalized biomass size spectra by area(protected and unprotected areas)of littoral fishes in the north western Mediterranean[30]
3 研究展望
自从粒径谱理论被引入水生生态系统研究以来,我国对于浮游生物、海洋底栖生物粒径谱的研究较为广泛,而对于鱼类粒径谱的研究甚少,几乎为空白。粒径谱理论引入为研究鱼类乃至海洋生态学提供了新的方法。与经典的依靠分类阶元进行鱼类生物群落特征的描述相比,粒径谱方法可以不依赖于分类学知识,避免因分类鉴定不准确而引起的计算误差,可以更好地增强对海洋生态系统群落特征以及鱼类物种生态和生理过程的认识,同时预测、评估鱼类群落的生产量、生产力、分布以及对环境进行监测。
当前大多数鱼类粒径谱理论的研究均是基于海上调查的生态过程,且许多理论预测的线性正态化生物量粒径谱的斜率为-1,同时也产生了一些新的问题需要我们今后去解决[57]:第一,与淡水生物量粒径谱相比,为什么海洋生物量粒径谱更多呈现线性关系;第二,普遍观察到的正态化生物量粒径谱的穹顶是否是稳定的;第三,在受到外界干扰恢复之后,鱼类群落能否有一个平衡状态的粒径结构[58]。
鱼类粒径谱的研究极大依赖于鱼类群落生物个体的粒径大小和所抽取的样本大小。研究样品的获得以及测量是目前学术界对鱼类粒径谱的研究存在差异性的主要原因。首先,不同的研究者所采取的研究方法、条件有所不同,必然造成结果出现差异;其次,多数研究者均是人工采样及测量数据,避免不了出现误差,不能真实反映鱼类群落特征。比如:测量与换算误差、划分粒径依据、样本大小、仪器设备新旧、研究区域差异、鱼类生活习性(产卵期、捕食期、竞争)以及季节差异等因素,都会影响研究结果;最后,因为研究对象和地域的时空差异,先前多数学者的研究成果不能很好地进行比较研究,给后人研究带来一定的困难。因此,在抽样调查时应抽取尽可能大且粒径范围广泛的生物体样本,确保所有粒径间隔上的生物体丰度都能被严格的估算到,并加大对新采样技术的研发与应用,如水声学和光学等,以取代费时费力的拖网采样,以便更好地反映整个鱼类群落的粒径结构和生态特征。同时,研发通用的基于粒径谱的鱼类生物量评估理论模型,并规范衡量标准,应该是未来鱼类粒径谱研究的趋势之一。
[1] 沈国英.海洋生态学(第3版)[M].北京:科学出版社,2010.
SHEN G Y.Marine ecology(The third edition)[M].Beijing:Science press,2010.
[2] BENOIT E,ROCHET M J.A continuous model of biomass size spectra governed by predation and the effects of fishing on them[J].Journal of Theoretical Biology,2004,226(1):9-21.
[3] BORGMANN U.Particle-size-conversion efficiency and total animal production in pelagic ecosystems[J].Canadian Journal of Fisheries and Aquatic Sciences,2011,39(5):668-674.
[4] BOUDREAU PR,DICKIE L M.Biomass spectra of aquatic ecosystems in relation to fisheries yield[J].Canadian Journal of Fisheries and Aquatic Sciences,2011,49(8):1528-1538.
[5] CYR H,PETERS R H.Biomass-size spectra and the prediction of fish biomass in lakes[J].Canadian Journal of Fisheries and Aquatic Sciences,2011,53(5):994-1006.
[6] LEACH J H,DICKIE L M,SHUTER B J,et al.A review of methods for prediction of potential fish production with application to the Great Lakes and Lake Winnipeg[J].Canadian Journal of Fisheries and Aquatic Sciences,2011,44(S2):471-485.
[7] SHELDON R W,PRAKASH A,SUTCLIFFE W H.The size distribution of particles in the ocean[J].Limnology and Oceanography,1972,17(3):327-340.
[8] SHELDON R W,SUTCLIFFE W H,PARANJAPE M A.Structure of pelagic food chain and relationship between plankton and fish production[J].Journal of the Fisheries Research Board of Canada,1977,34(12):2344-2353.
[9] SPRULESW G,BRANDT SB,STEWART D J,et al.Biomass size spectrum of the Lake Michigan Pelagic food web[J].Canadian Journal of Fisheries and Aquatic Sciences,1991,48(1):105-115.
[10] BORGMANN U,WHITTLE D M.Particle-sizeconversion efficiency and contaminant concentrations in Lake Ontario Biota[J].Canadian Journal of Fisheries and Aquatic Sciences,2011,40(3):328-336.
[11] COTTINGHAM K L.Nutrients and zooplankton as multiple stressors of phytoplankton communities:Evidence from size structure[J].Limnology and Oceanography,1999,44(3):810-827.
[12] SHELDON R W,PARSONST R.A continuous size spectrum for particulate matter in the sea[J].Journal of the Fisheries Research Board of Canada,2011,24(5):909-915.
[13] 周林滨,谭烨辉,黄良民,等.水生生物粒径谱/生物量谱研究进展[J].生态学报,2010,30(12):3319-3333.
ZHOU L B,TAN Y H,HUANG L M,et al.The advances in the aquatic particle/biomass size spectra study[J].Acta Ecologica Sinica,2010,30(12):3319-3333.
[14] 王睿照,张志南.海洋底栖生物粒径谱的研究[J].海洋湖沼通报,2003(4):61-68.WANG R Z,ZHANG Z N.A study on the size spectra of benthos[J].Transactions of Oceanology and Limnology,2003(4):61-68.
[15] KERR SR.Theory of size distribution in ecological communities[J].Journal of the Fisheries Research Board of Canada,2011,31(12):1859-1862.
[16] THIEBAUX M L,DICKIE L M.Models of aquatic biomass size spectra and the common structure of their solutions[J].Journal of Theoretical Biology,1992,159(2):147-161.
[17] THIEBAUX M L,DICKIE L M.Structure of the body-size spectrum of the biomass in aquatic ecosystems:A consequence of allometry in predatorprey interactions[J].Canadian Journal of Fisheries and Aquatic Sciences,1993,50(6):1308-1317.
[18] SILVERT W,PLATT T.Energy flux in the pelagic ecosystem: A time-dependent equation[J].Limnology and Oceanography,1978,23(4):813-816.
[19] BOUDREAU PR,DICKIE L M,KERR SR.Bodysize spectra of production and biomass as systemlevel indicators of ecological dynamics[J].Journal of Theoretical Biology,1991,152(3):329-339.
[20] ZHOU M,HUNTLEY M E.Population dynamics theory of plankton based on biomass spectra[J].Marine Ecology Progress Series,1997,159(159):61-73.
[21] ROSEN C J.Modelling an exploited marine fish community with 15 parameters-results from a simple size-based model[J]. Ices Journal of Marine Science,2006(63):1029-1044.
[22] LAWR,PLANK M J,JAMESA,et al.Size-spectra dynamics from stochastic predation and growth of individuals[J].Ecology,2009,90(3):802-811.
[23] ROSSBERG A G.6-a complete analytic theory for structure and dynamics of populations and communities spanning wide ranges in body size[J].Advances in Ecological Research,2012(46):427-521.
[24] KERR SR,DICKIE L M.The biomass spectrum:A predator-prey theory of aqautic production[M].Japan:Columbia University Press,2001.
[25] PLATT T,DENMAN K.The structure of pelagic marine ecosystems[J].Rapports Et Proces Verbaux Des Reunions,2010(173):60-65.
[26] 邓 可,张志南,黄 勇,等.南黄海典型站位底栖动物粒径谱及其应用[J].中国海洋大学学报(自然科学版),2005,35(6):1005-1010.DENG K, ZHANG Z N, HUANG Y, et al.
Application of benthic biomass size spectra of macroand microfuana at typical stations in the Southern Yellow Sea, China[J]. Periodical of Ocean University of China(Natural Science Edition),2005,35(6):1005-1010.
[27] WITEK Z,KRAJEWSKASOLTYSA.Some examples of the epipelagic plankton size structure in high latitude oceans[J].Journal of Plankton Research,1989,11(6):1143-1155.
[28] SPRULESW,MUNAWAR M.Plankton size spectra in relation to ecosystem productivity, size, and perturbation[J].Canadian Journal of Fisheries and Aquatic Sciences,1986,43(9):1789-1794.
[29] PLATT T,DENMAN K.Organisation in the pelagic ecosystem [J]. Helgoländer Wissenschaftliche Meeresunter suchungen,1977,30(1-4):575-581.
[30] MACPHERSON E,GORDOA A,GARCIA-RUBIES A.Biomass size spectra in littoral fishes in protected and unprotected areas in the NWMediterranean[J].Estuarine Coastal and Shelf Science,2002,55(5):777-788.
[31] SHIRAYAMA Y.Ecology of deep-sea meiobenthos in the western Pacific[J]. Journal of the Oceanographical Society of Japan,1989,45(45):83-93.
[32] BANSE K,MOSHER S.Adult body mass and annual production/biomass relationships of field populations[J].Ecological Monographs,1980,50(3):355-379.
[33] JUNG S,HOUDE E D.Fish biomass size spectra in Chesapeake Bay[J].Estuaries and Coasts,2005,28(2):226-240.
[34] JENNINGS S.Size-based analysis of aquatic food webs[J].Oxford University Press,2005:86-97.
[35] CYR H.Individual energy use and the allometry of population density:Scaling in biology[M].New York:Oxford University Press,2000.
[36] BROWN JH,GILLOOLY JF.Ecological food webs:High-quality data facilitate theoretical unification[J]. Proceedings of the National Academy of Sciences,2003,100(4):1467-1468.
[37] JENNINGS S,BLANCHARD J L.Fish abundance with no fishing:predictions based on macroecological theory[J].Journal of Animal Ecology,2004,73(4):632-642.
[38] 刘 云,王际英,李宝山,等.海水鱼类微量元素需求研究进展[J].海洋渔业,2015,37(4):378-385.
LIU Y,WANG J Y,LI B S,et al.Research progress on requirements of marine fishes for trace elements[J].Marine Fisheries,2015,37(4):378-385.
[39] 赵书燕,林黑着,黄 忠,等.不同蛋白质水平下添加小肽对石斑鱼生长、消化酶、血清生化和抗氧化能力的影响[J].南方水产科学,2016,12(3):15-23.
ZHAO SY,LIN H Z,HUANG Z,et al.Effect of small peptide supplementation at different protein levels on growth performance,digestive enzymes activities,serum biochemical indices and antioxidant abilities of grouper(Epinephelus akaara)[J].South China Fisheries Science,2016,12(3):15-23.
[40] CHANDLER M,PETERS R.Mapping trophic continua of benthic foodwebs: Body size-δ15N relationships[J].Marine Ecology Progress Series,1998(174):301-306.
[41] 纪炜炜,李圣法,陈雪忠.鱼类营养级在海洋生态系统研究中的应用[J].中国水产科学,2010,17(4):878-887.
JI W W,LI S F,CHEN X Z.Application of fish trophic level in marine ecosystem[J].Journal of Fishery Sciences of China,2010,17(4):878-887.
[42] MACPHERSON E,GORDOA A.Biomass spectra in benthic fish assemblages in the Benguela System[J].Marine Ecology Progress Series,1996,138(1-3):27-32.
[43] EMMRICH M,BRUCET S,RITTERBUSCH D,et al.Size spectra of lake fish assemblages:Responses along gradients of general environmental factors and intensity of lake-use[J].Freshwater Biology,2011,56(11):2316-2333.
[44] 王跃中,孙典荣,贾晓平,等.捕捞压力和气候变化对东海马面鲀渔获量的影响[J].南方水产科学,2013,9(1):8-15.
WANG Y Z,SUN D R,JIA X P,et al.Influence of fishing pressure and climate change on filefish catches in East China Sea[J].South China Fisheries Science,2013,9(1):8-15.
[45] ZWANENBURG K C T.The effects of fishing on demersal fish communities of the Scotian Shelf[J].Ices Journal of Marine Science,2000,57(3):503-509.
[46] FRY B,QUINONESR.Biomass spectra and stable isotope indicators of trophic level in zooplankton of the northwest Atlantic[J].Marine Ecology Progress Series,1994,112(1-2):201-204.
[47] JENNINGS S,PINNEGAR J K,POLUNIN N V C,et al.Linking size-based and trophic analyses of benthic community structure[J].Marine Ecology Progress Series,2002,226(1):77-85.
[48] PAHL WC.Food webs and ecological networks across temporal and spatial scales[J].Oikos,1993,67(3):415-432.
[49] 晏 磊,谭永光,杨炳忠,等.基于张网渔业休渔前后的黄茅海河口渔业资源群落比较[J].南方水产科学,2016,12(6):1-8.
YAN L,TAN Y G,YANG B Z,et al.Comparison on resources community of stow-net fishery before and after fishing off season in Huangmaohai Estuary[J].South China Fisheries Science,2016,12(6):1-8.
[50] JENNINGS S,REYNOLDS J D.Body size,exploitation and conservation of marine organisms[M]//Body size:The Structure and Function of Aquatic Ecosystems.England:Cambridge University Press,2007.
[51] DUPLISEA D E,KERR S R.Application of a biomass size spectrum model to demersal fish data from the Scotian Shelf[J].Journal of Theoretical Biology,1995,177(3):263-269.
[52] BLANCHARD J L,ANDERSEN K H,SCOTT F,et al.Evaluating targets and trade-offs among fisheries and conservation objectives using a multispecies size spectrum model[J].Journal of Applied Ecology,2014,51(3):612-622.
[53] GRAHAM N,DULVY N K,JENNINGS S,et al.Size-spectra as indicators of the effects of fishing on coral reef fish assemblages[J].Coral Reefs,2005,24(1):118-124.
[54] TREBILCO R,BAUM J K,SALOMON A K,et al.Ecosystem ecology:Size-based constraints on the pyramids of life[J]. Trends in Ecology and Evolution,2013,28(7):423-431.
[55] PETERSON I,WROBLEWSKI J S.Mortality rate of fishes in the pelagic ecosystem[J]. Canadian Journal of Fisheries and Aquatic Sciences,2011,41(7):1117-1120.
[56] THYGESEN U H,FARNSWORTH K D,ANDERSEN K H,et al.How optimal life history changes with the community size-spectrum[J].Proceedings Biological Sciences,2005,272(1570):1323-1331.
[57] SPRULES W G,BARTH L E.Surfing the biomass size spectrum:Some remarks on history,theory,and application[J].Canadian Journal of Fisheries and Aquatic Sciences,2015,12(9):3459-3466.
[58] ANDERSEN K H,JACOBSEN NS,FARNSWORTH K D.The theoretical foundations for size spectrum models of fish communities[J].Canadian Journal of Fisheries and Aquatic Sciences,2016,73(4):575-588.
Advances in fish particle size spectra study
GUO Jian-zhong1,2,CHEN Zuo-zhi1,XU Shan-nan1
(1.Key Laboratory of South China Sea Fishery Resources Exploitation&Utilization of Ministry of Agriculture;Key Laboratory of Fishery Ecology and Environment of Guangdong Province,South China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Guangzhou 510300,China;2.College of Fisheries and Life Science,Shanghai Ocean University,Shanghai 201306,China)
Particle size spectrum is a kind of curve graph that represents the relationship between biomass and particle size.It can reflect the structure and function of the ecosystem and its dynamic connection inside the system,which is currently becoming a hotspot issue of the aquatic ecology.At present,the research on fish particle size spectrum in our country is quite little,but there are more researches in this field and more advanced theory and methods in foreign country,which can provide worthy reference for domestic scholars.In this paper,a large number of fish particle size spectrum research literatures and the development of recent years were summarized.The paper introduced theoretical principles,research methods,influence factors and application of particle size spectrum,which referred to fish production and biomass assessment,ecological monitoring,reflection on the characteristics of fish community and ecosystem structure and function,and the prediction of human activities and natural disturbance impact on marine ecosystem.Fishing intensity,marine productivity level,the level of nutrition,fish community habitat environment,fish community species competition and compensation efficiency,seasonal changes and the types of fishing gears and meshes are main factors affecting the fish particle size spectrum.Most of the current fish particle size spectrum theory studies were based on the investigation of marine ecological processes,and many theoretical predictions of the linear slope of normalized biomass particle size spectra as-1.The slope,intercept and regression coefficient respectively affected the characteristics of fish community.The slope represented the relationship between the biomass and the size.The intercept represented the magnitude of productivity level.The regression coefficient indicated the degree of community deviation from the steady state.It also brought some new problems needed to solve in the future:firstly,compared with fresh biomass particle size spectra,why did the marine biomass particle size spectra show more linear relationships?Secondly,was the generally observed normalized biomass particle size spectrum of dome is stable?Thirdly,after the resumption from the interference by environment,did the fish community have a balanced state of the particle size structure?Based on the deficiencies and problems in current researches,this paper proposes the application of new observation and sampling technology,and the generally theoretical model research based on fish particle size spectrum.
particle size spectrum;fish particle size spectrum;ecosystem
S 931
A
1004-2490(2017)05-0582-10
2016-10-08
国家重点基础研究发展计划(2015CB452904);广东省自然科学基金项目(2015A030313785);国家自然科学基金项目(31100362);广州市科技计划项目珠江科技新星专项(2014J2200020)
郭建忠(1990-),男,河南省南召县人,硕士研究生,主要从事渔业生态研究。
E-mail:1115982121@qq.com
徐姗楠,副研究员。Tel:020-89108303,E-mail:xushannan@scsfri.ac.cn
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05 07:38:24
物理之友(2020年12期)2020-07-16 05:39:16
探索科学(2017年5期)2017-06-21 21:16:16
统计与决策(2017年2期)2017-03-20 15:25:23
福建中学数学(2016年7期)2016-12-03 07:10:28
光学精密工程(2016年1期)2016-11-07 09:01:53
金色少年(奇趣科普)(2016年8期)2016-09-21 02:04:58
电测与仪表(2016年15期)2016-04-12 00:30:58
电测与仪表(2016年6期)2016-04-11 12:05:54
湖北师范大学学报(自然科学版)(2015年1期)2016-01-10 08:41:14
