
鼠疫耶尔森氏菌环二鸟苷酸代谢及生物被膜形成调控研究进展
2017-11-03 05:58:39赵菊平孙义成
生物工程学报 2017年9期
赵菊平,孙义成
中国医学科学院/北京协和医学院 病原生物学研究所,北京 100730
鼠疫耶尔森氏菌环二鸟苷酸代谢及生物被膜形成调控研究进展
赵菊平,孙义成
中国医学科学院/北京协和医学院 病原生物学研究所,北京 100730
鼠疫耶尔森氏菌(Yersinia pestis,以下简称“鼠疫菌”)是烈性传染病鼠疫的病原菌,以鼠蚤作为传播媒介。鼠疫菌在其传播媒介鼠蚤的前胃中形成生物被膜从而促进其在宿主间传播。鼠疫菌生物被膜的形成受第二信使分子环二鸟苷酸(c-di-GMP)的正向调控。鼠疫菌中c-di-GMP由二鸟苷酸环化酶(DGC)HmsT和HmsD合成,由磷酸二酯酶(PDE)HmsP降解。文中主要介绍影响鼠疫菌环二鸟苷酸代谢及生物被膜形成的调控因子,并对其作用机制进行讨论和总结。
鼠疫耶尔森氏菌,生物被膜,环二鸟苷酸,二鸟苷酸环化酶,磷酸二酯酶
鼠疫耶尔森氏菌Yersinia pestis由相对温和的肠道致病菌假结核耶尔森氏菌(Yersinia pseudotuberculosis,以下简称“假结核菌”)进化而来。鼠疫菌主要以跳蚤为传播媒介在哺乳动物间传播。跳蚤叮咬感染鼠疫菌的宿主后,鼠疫菌感染并在跳蚤前胃中形成生物被膜(Biofilm)进而形成“栓塞”,部分或完全阻塞血液进入中肠吸收利用,饥饿的跳蚤反复叮咬宿主并使带菌的血液回流通过叮咬的伤口感染宿主[1]。因此鼠疫菌生物被膜形成能力是其定殖于跳蚤前胃并传播感染的关键因素。
与其他细菌一样,鼠疫菌生物被膜形成受细菌第二信使分子环二鸟苷酸(3ʹ-5ʹ-cyclic diguanylic acid,c-di-GMP)浓度水平的正向调控,c-di-GMP同时促进负责合成并转运生物被膜胞外多糖(Exopolysaccharide,EPS)的hmsHFRS操纵子表达[2]。c-di-GMP分别由二鸟苷酸环化酶(Diguanylate cyclases,DGC)合成和磷酸二酯酶(Phosphodiesterases,PDE)降解。鼠疫菌编码2个DGC(HmsT和HmsD)和1个PDE(HmsP)。3个代谢酶的表达及活性受到环境信号和细胞内调节因子多水平的调控,进而影响菌体内c-di-GMP浓度水平,精细调控生物被膜的形成以适应不同的环境条件(图1)。本文重点就c-di-GMP信号通路中环境条件及细胞内调节因子对c-di-GMP代谢酶、hmsHFRS的表达和活性调控进而调控细胞内c-di-GMP浓度及生物被膜形成这一领域进行综述。
1 细菌生物被膜
1978年Costerton等[3]首次提出了生物被膜的相关理论。细菌粘附于物体表面时,形成一个含有微菌落的基质层,使细菌得以与物体表面相结合,随着粘附的微菌落的增大和数目增多,这些包着微菌落的基质相互融合而形成了生物被膜。生物被膜主要包含多糖、蛋白质、脂质和胞外DNA(eDNA),以及淀粉、纤维素、菌毛、鞭毛、细胞裂解物等,其中最主要的成分是水,比例高达97%,eDNA在生物被膜基质结构组织和稳固中具有重要作用[4-5]。
生物被膜是细菌为了适应自然环境而普遍存在的一种保护性的生存方式,由单个或多个细菌物种组成,似一个空间异质的生态系统,是一种高度异质性、高度组织化,促进其自身功能及特性的结构。生物被膜细胞间可以相互通讯并共享生物被膜基质中所有营养资源及信号分子[6]。以生物被膜的方式生存的细菌与单个浮游的细菌相比,具有很多新的特性:生长在生物被膜中的近距离细胞间相互作用,使基因交换频率增加;生物被膜作为一种多细胞组织结构,其水合作用及捕获和降解消化生长环境中营养资源的能力均有利于菌体生长代谢[7];由于生物被膜基质的阻滞作用阻止药物扩散,基质中所含的水解酶对药物的降解作用,及生物被膜深处的菌体因为营养缺乏,菌体生长缓慢等因素,均使菌体对抗菌药物的耐受性增强[8-10];另外,生物被膜中大量的水分还使细菌对干燥环境的耐受性大大增强。细菌在高等生物体内以生物被膜的形式定殖容易造成持续感染,污染医疗器械和移植物等,给人类带来了很多难题。
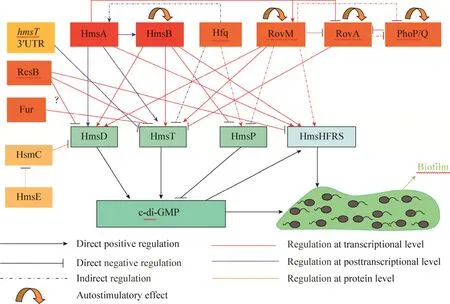
图1 鼠疫菌中不同调控因子对c-di-GMP代谢及生物被膜形成的调控作用(c-di-GMP代谢及生物被膜形成在细胞内受到多种类型调控因子在转录水平、转录后水平以及蛋白-蛋白相互作用的调控.红色框内为sRNA,橙色框内为调控蛋白,黄色框内为mRNA上的顺式作用序列.HmsC、HmsE与HmsD由同一个操纵子编码,在细胞内调控HmsD的活性,HmsHFRS负责生物被膜胞外多糖合成)Fig.1 The regulators involved in modulation ofc-di-GMP metabolism and biofilm formation inY.pestis.Thec-di-GMP metabolism and biofilm formation are controlled by different cellular regulators at transcriptional, posttranscriptional and protein level.The red boxes contain sRNA, orange boxes contain protein and in the yellow box is the cis-acting sequence on mRNA.HmsC and HmsE are encoded by one operon with HmsD and control the activity of HmsD.HmsHFRS is responsible for biosynthesis of extracellular polysaccharide(EPS)matrix of biofilm.
2 鼠疫菌生物被膜
鼠疫菌是鼠疫这一种古老的自然疫源性疾病的病原菌,在5021−7022年前由假结核菌进化而来[10]。假结核菌在环境中广泛存在,可以通过食物或水经口传播而引发多种哺乳动物相对温和的肠道疾病。鼠疫菌依赖于吸血昆虫跳蚤作为传播媒介,毒性和入侵性都很强,可引发全身系统性感染的致死性疾病。人类历史上记载的鼠疫致死人数高达2亿,并曾发生过3次世界鼠疫大流行,给人类造成巨大的灾难,被冠以“最具毁灭性传染病”的名头[11]。
大部分假结核菌株能在跳蚤消化道内生长,但是都不能形成生物被膜并阻塞前胃。鼠疫菌由假结核菌进化而来,获得了在跳蚤中肠和前肠定殖并在前胃形成生物被膜的能力,从而实现以跳蚤作为媒介的疾病传播[12-13],使得鼠疫菌在全球范围内迅速传播。感染了鼠疫菌的哺乳动物死亡前常出现严重的败血症,而这种高浓度的带菌血再次被跳蚤吮吸后就足以感染跳蚤并维持鼠疫菌的传播循环[14]。鼠疫菌借助跳蚤传播最重要的方式依赖于其在跳蚤消化道定殖并在跳蚤前胃形成生物被膜的能力,前胃是连接跳蚤食管和中肠的前肠瓣膜结构,跳蚤前胃形成的生物被膜部分或完全阻塞血液摄取,饥饿的跳蚤反复叮咬宿主,同时使染菌的血液回流通过叮咬伤口直接感染宿主,极大促进了鼠疫菌的传播和感染能力[15-17]。鼠疫菌经血液传播方式的获得是其进化中的关键步骤。
鼠疫菌传播依赖于其在跳蚤前胃形成生物被膜,hmsHFRS操纵子是鼠疫菌在前胃定殖并形成生物被膜必需的,组成生物被膜的胞外多糖(EPS)由hmsHFRS操纵子负责合成并转运到胞外,HmsH和HmsF是外膜蛋白,HmsR和HmsS是内膜蛋白,HmsR具有糖基转移酶活性,HmsF具多糖去乙酰化酶及糖基水解酶活性[18-20]。EPS表达和转运受到细胞内c-di-GMP浓度在转录后水平的正向调控[2,21]。c-di-GMP是细菌中普遍存在的第二信使分子,是调控细菌生物被膜形成的核心元件,整合细胞内外多种调控因素参与生物被膜形成多个过程的调控作用。并对病原菌致病因子表达、运动性和细胞周期等起到调控作用[22-24]。
3 c-di-GMP调控系统
1987年 Ross等[25]首次在木葡糖酸醋杆菌Acetobacter xylinum中鉴定出c-di-GMP为纤维素合酶的别构激活剂(Allosteric activator)。c-di-GMP是细菌细胞内复杂功能的关键调节因子,高c-di-GMP水平促进胞外多糖产生和表面粘附,促进生物被膜形成。而低c-di-GMP水平促进鞭毛基因表达和各种形式的细胞运动,控制细菌菌体在浮游状态(Planktonic state)和生物被膜模式间转换[24],并与细菌致病性相关。
c-di-GMP代谢受合成酶二鸟苷酸环化酶(DGC)和降解酶磷酸二酯酶(PDE)的表达与活性调控。DGC将两分子的GTP合成c-di-GMP,PDE使c-di-GMP线性化为pGpG,在细胞中被进一步降解为 GMP(图2)。DGC活性与GGDEF结构域相关。PDE活性依赖于EAL或HD-GYP结构域。GGDEF结构域含 GG[D/E]EF保守序列,大多数DGC还含有c-di-GMP结合的别构调控位点RxxD,位于催化位点上游5个氨基酸处,起到产物抑制的反馈调节作用,避免细胞中过量 GTP消耗并控制c-di-GMP的量[26-27]。EAL结构域由7个分离的保守氨基酸残基构成并螯合Mg2+[28]。HD-GYP结构域蛋白首先在植物病原菌野油菜黄单胞菌Xanthomonas campestris中被鉴定,具有降解c-di-GMP的功能,由7个螺旋紧密折叠形成[29-30]。
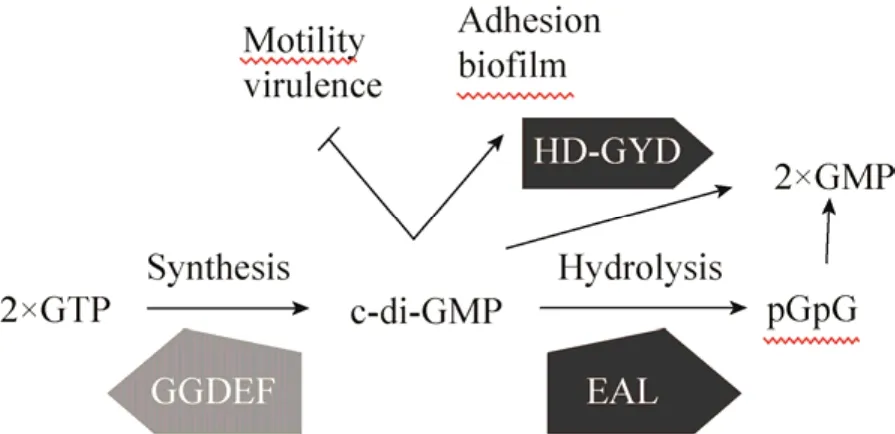
图2 c-di-GMP代谢及生理功能(c-di-GMP由GGDEF结构域蛋白合成,由EAL或HD-GYD结构域蛋白降解,在细胞内具有多种调节功能)Fig.2 Schematic representation metabolism and physiological functions ofc-di-GMP.c-di-GMP inY.pestisis synthesized by two GGDEF domain proteins and degraded by EAL or HD-GYD domain proteins.Various cellular functions are regulated byc-di-GMP.
GGDEF、EAL、HD-GYP结构域蛋白广泛分布于细菌中,而且许多细菌编码蛋白数量众多,尤其是γ-变形菌门(Gamma Proteobacteria),例如铜绿假单胞菌Pseudomonas aeruginosa不同菌株编码40个左右这类蛋白,大肠杆菌Escherichia coli编码28个,霍乱弧菌Vibrio cholera编码高达56个。并且这些蛋白大多数含有信号感应结构域及空间定位信号(如PAS、REC、HAMP、GAF结构域),或是信号二次输出结构域(如HDOD),及跨膜螺旋等多种定位信号[28,31],印证了c-di-GMP信号系统多样性和多水平复杂的功能。而鼠疫菌的c-di-GMP代谢酶网络相对简化,仅含2个合成酶HmsT、HmsD和1个降解酶HmsP。这些蛋白的表达及活性受到环境信号的调控而影响菌体内c-di-GMP的代谢,从而精细调控细菌生物被膜的形成使其适应不同的环境。
4 鼠疫菌c-di-GMP代谢调控因子及其对生物被膜的影响
4.1 c-di-GMP代谢酶
鼠疫菌含10个编码 GGDEF及 EAL或HD-GYP结构域的基因,只有hmsT、hmsP及位于hmsCDE操纵子中的hmsD是有功能的,其他的是假基因或是编码没有催化功能的蛋白或是不表达[32-33]。HmsT是内膜蛋白,含GGDEF结构域,具有DGC活性,催化鼠疫菌中c-di-GMP合成,正调节生物被膜形成[32,34-35],细胞内多胺在翻译水平激活hmsT表达[36]。HmsP含 EAL结构域和GGDEF结构域,但是GGDEF结构域是退化失活的。HmsP EAL结构域具有PDE活性,能够特异性降解c-di-GMP,负调节生物被膜形成,其中E506和L508氨基酸残基起到关键作用[35,37]。HmsD在跳蚤体内生物被膜形成中起主要作用,HmsT在体外生物被膜形成中起主要作用[33],说明两种DGC在不同的条件下受到不同的调控。而两种条件下,hmsT、hmsD转录水平是一样的,所以两种环境条件的调控作用可能主要发生在转录后水平[33]。
HmsD由hmsCDE操纵子编码,是一个含有GGDEF结构域的DGC,受到HmsC及HmsE蛋白作用的调控。hmsCDE三基因共转录,HmsD为内膜蛋白,含2个跨膜结构域,分隔开1个周质空间结构域(Periplasmic domain,PD)、细胞质侧的1个HAMP信号转换结构域和1个信号输出结构域GGDEF。HmsC是周质空间蛋白,其N末端跨膜结构域介导C末端转运到周质空间,N末端跨膜结构在 HmsC成熟时被剪切下来。HmsC通过与HmsD的PD结合而影响HmsD蛋白稳定性,抑制 HmsD活性,从而使细胞中c-di-GMP水平降低,生物被膜形成水平减弱。保守的氨基酸残基E156对于HmsC的抑制功能是必需的。HmsE为外膜脂蛋白,作用于HmsC抵消其抑制作用而间接激活 HmsD活性。跳蚤体内的生物被膜形成实验同样印证了 HmsE的激活作用。并且HmsC、HmsE的调节功能在各鼠疫菌菌株间是保守的[38-40]。
4.2 Rcs磷酸中继系统的调控作用
Rcs磷酸中继系统(Rcs phosphorelay system)包含蛋白RcsB、RcsC、RcsD及辅助蛋白RcsA。RcsB存在于细胞质中,是信号应答调节因子,通过与DNA结合发生作用。磷酸化激活的RcsB单一作用或是与辅助蛋白RcsA组成RcsAB复合物,作为转录调控因子识别特定的反向重复序列TAAGAAT-ATTCTTA,与被调节基因的启动子近端序列(Promoter-proximal sequence)结合,调控基因表达[41-42]。被调节的基因主要负责胞外多糖合成、鞭毛运动性及Rcs系统自调节。
鼠疫菌由假结核菌进化而来时,rcsA被阴性选择为无活性的假基因,在鼠疫菌内依然能够表达,但是没有生物被膜相关表型[43]。转化有功能的rcsA到鼠疫菌中表达时,鼠疫菌不再能够阻塞跳蚤前胃[44]。鼠疫菌中RcsD可以解除RcsB对hmsT转录及生物被膜形成的抑制作用,可能是通过使RcsB去磷酸化的方式[43,45]。
单一的RcsB能够抑制生物被膜形成及c-di-GMP产生,与RcsA组成复合物RcsAB时,抑制效应更强,单一的RcsA则无此效应[40,45]。田鼠型201鼠疫菌菌株(Y.pestis Microtusstrain201)的研究表明,RcsAB可以抑制hmsCDE、hmsT、hmsHFRS转录,并以间接的方式促进hmsP表达。RcsAB与hmsT、hmsHFRS启动子–10区及转录起始位点的特异性结合位点紧密结合,阻碍RNA聚合酶进入从而抑制转录。然而hmsCDE的RcsAB结合位点在启动子–35区上游[40,45]。鼠疫菌KIM6+菌株中,RcsB同样抑制hmsT转录,而促进hmsCDE转录。不同菌株中,HmsD有不同的调控机制,可能受到其他因素的影响,而详细的机制还有待进一步的研究[40,46]。
4.3 hmsT3ʹUTR的调控作用
Zhu等[44]的研究发现hmsT3ʹUTR使mRNA的稳定性降低,负调控hmsT表达,从而抑制c-di-GMP合成,负调节鼠疫菌生物被膜形成。研究表明,3ʹUTR并未通过顺式编码的sRNA介导调控 mRNA衰减。之前有报道Hfq抑制hmsT表达[47-48],而双删除hmsT3ʹUTR和hfq,hmsTmRNA水平比单一删除hfq更高,说明Hfq依赖的反式编码sRNA的调控作用在3ʹUTR介导的调控中并未占据主要作用。进一步的研究揭示了3ʹUTR包含多个顺式作用的调节序列共同起到调节作用,并且调节效应是累加性的。多核苷酸磷酸化酶 PNPase在3ʹUTR介导的hmsTmRNA降解中发挥作用[44]。
hmsT转录不受温度影响,但是在环境温度(21℃)时HmsT水平高,在哺乳动物体温(37℃)时 HmsT水平低,鼠疫菌在从跳蚤寄生到哺乳动物宿主时就经历这样的生长温度变化。hmsT3ʹUTR在37℃时对基因表达的抑制效应比在环境温度时更强。故而,鼠疫菌在从跳蚤寄生到哺乳动物宿主时可能经历了3ʹUTR介导的快速mRNA降解,EPS合成水平降低,同时细菌毒性增强[44]。
4.4 PhoP/PhoQ调节系统的调控作用
phoPQ存在于2个操纵子中,PhoP起调节作用,而PhoQ是感应蛋白。在Mg2+浓度低时,PhoQ被磷酸化,进而磷酸基团被转移到PhoP,磷酸化的PhoP调节下游基因表达。phoPQ操纵子由PhoP自诱导上调表达[49]。在鼠疫菌和假结核菌中PhoP调节生物被膜形成。并且假结核菌中PhoP抑制hmsT表达,phoP野生型菌株不表达或很少表达HmsT[21]。phoP突变株在跳蚤前胃及体外形成的生物被膜粘附性没有野生型强,更易在收缩蠕动或水流作用力下脱离附着表面,降低鼠疫菌阻塞跳蚤前胃并传播感染的能力。鼠疫菌感染跳蚤时,由于低pH及其他应激条件存在,PhoP/PhoQ被诱导表达,是鼠疫菌形成生物被膜阻塞跳蚤前胃,促进鼠疫菌传播所必需的[50-51]。
4.5 RovM/RovA的调控作用
RovM是LysR型转录因子,对鼠疫菌致病性及生物被膜形成等方面具有广泛的调控作用[52]。RovA是鼠疫菌致病性必需的因子[53]。PhoP可直接作用于rovA抑制转录[54]。RovM 促进c-di-GMP产生及生物被膜形成,而RovA起相反的作用,下调c-di-GMP产生及生物被膜形成。RovM 以剂量依赖性的方式与hmsT、hmsCDE启动子近端序列结合直接调控转录,促进基因表达。RovM 通过间接的方式调控hmsHFRS、hmsP转录,促进hmsHFRS表达,抑制hmsP表达。RovA与hmsT启动子近端序列结合,直接抑制转录[55-56],但是对hmsCDE、hmsHFRS和hmsP转录无调控作用。
RovM/RovA还可以调控调节基因表达而间接调控c-di-GMP代谢及生物被膜形成。RovM可以直接与rovA的启动子近端序列结合抑制转录,并能够间接抑制phoPQ转录,以及与自身基因启动子近端序列结合,自我激活转录表达。RovA可通过调节自身启动子活性,促进自身基因表达,并能够间接抑制phoPQ转录。
温度从26℃变化到37℃时,rovM表达上调,rovA表达下调[53,56]。在跳蚤体内生长时,rovM表达上调,rovA表达下调,促进生物被膜形成[57]。
4.6 铁吸收调节因子Fur的调控作用
铁吸收调节因子(Fur)在细菌中具有广泛的功能,并在鼠疫菌中控制复杂的调节级联反应,Fur特异性的结合序列为AATGATAATNATT ATCATT,具有9-1-9的反向重复结构[58-59]。Fur抑制鼠疫菌 EPS合成并抑制生物被膜形成,hmsT有2个启动子近端区域含Fur结合序列,分别位于起始密码子上游102–71及283–244处,Fur与hmsT启动子近端区域特异性结合,抑制hmsT转录。目前尚未发现 Fur对hmsHFRS及hmsCDE的转录调节作用。Fur通过抑制hmsT表达而抑制c-di-GMP产生,为鼠疫菌生物被膜形成的负调节因子[60]。
4.7 sRNA及伴侣蛋白Hfq的调控作用
cDNA克隆及RNA测序的方法已经被用来鉴定鼠疫菌全基因组水平的sRNA(Small regulatory RNAs),其中的一些sRNA也已经被鉴定出具有特定的基因表达调控功能[61-64]。HmsB是第一个被报道具有生物被膜形成调控作用的sRNA[65]。hmsB位于hmsC与其上游基因的间隔序列,长262 bp,与hmsC向相反方向转录。HmsB促进c-di-GMP浓度水平上升及生物被膜胞外多糖合成。HmsB促进生物被膜正调控基因hmsT、hmsCDE、hmsHFRS及自身的启动子活性,促进转录。而hmsB突变菌株,hmsPmRNA及蛋白水平均上调,但是并不影响hmsP的启动子活性,可见HmsB可能在转录后水平负调控hmsP基因表达[65]。然而,是否 HmsB也与hmsT、hmsCDE、hmsHFRS及hmsBmRNA翻译起始区域作用而调控翻译过程,或者是基因的转录因子或是转录抑制因子受到调控而调控基因转录过程还有待进一步的研究阐明。
鼠疫菌进化中获得的质粒 pPCP1编码的sRNA HmsA在转录水平促进hmsHFRS表达,在转录后水平促进hmsT及hmsCDE表达。另外HmsA在转录后水平通过促进hmsBmRNA稳定性促进hmsB表达。HmsA还在转录水平抑制rovA、促进rovM表达[66]。HmsA的调控作用依赖于伴侣蛋白Hfq的作用[63]。
Hfq协助sRNA的基因表达调控作用,促进并稳定 sRNA与靶向的mRNA相互作用,Hfq还能调控适当水平的sRNA表达,并保护sRNA不被核酸酶降解[67-68]。在体外实验中,Hfq作用于hmsP启动子区域,在转录水平促进hmsP表达,同时作用于hmsTmRNA在转录后水平抑制hmsT表达,从而抑制c-di-GMP产生及生物被膜形成。在 BHI培养基中,Hfq负调控生物被膜形成,并且调控作用不受温度影响[48]。然而在含0.2%半乳糖的TMH培养基中,Hfq正调控生物被膜形成[51]。可见环境和营养条件可能对 Hfq对生物被膜的调控作用有影响。Hfq对于鼠疫菌在跳蚤前胃形成生物被膜是必需的,hfq突变菌株不能在跳蚤前胃形成生物被膜并阻塞前胃[47]。
4.8 环境条件的调控作用
除了细胞内调控因子,环境条件也对c-di-GMP代谢具有显著的调控作用。hmsT在26℃时表达水平高,温度过渡到37℃时HmsT蛋白被大量降解。与26℃相比,37℃时HmsP的酶活性也显著提高。二者可能均参与了鼠疫菌生物被膜形成的温度调控作用,HmsT可能在其中起到主要作用[37,69]。
Ren等[70]之前的工作选择了12个体外条件模拟跳蚤消化道内的条件,以评价不同的条件对c-di-GMP代谢和生物被膜形成的调控作用,即温度、二硫苏糖醇(Dithiothreitol,DTT)、蔗糖、CaCl2、MgCl2、FeCl2、CuSO4、NaCl、十二烷基硫酸钠(Sodium dodecyl sulfate,SDS)、2,2’-联吡啶(2,2’-dipyridyl DIP)、氧化环境(H2O2)、酸性条件(pH5)。
在4mmol/L CaCl2、4mmol/L MgCl2、2mmol/L FeCl2、100mmol/L DIP、1mmol/L CuSO4、0.01%SDS、6%蔗糖、4mmol/L DTT、10mmol/L H2O2、4% NaCl、pH5、21℃、26℃、37℃条件时,细菌生长没有受到显著影响,在此浓度条件下,蔗糖、CuSO4、CaCl2、SDS和 DTT存在时,c-di-GMP浓度上升,生物被膜形成增加;在温度上升,及 FeCl2、NaCl条件下时,c-di-GMP浓度下降,生物被膜形成减少;H2O2、DIP作用时,生物被膜及c-di-GMP浓度均不受影响。而温度下降及 MgCl2作用时,生物被膜形成增加,但c-di-GMP水平没有显著改变,也许是通过影响其他调节因子而调控生物被膜形成。
同样浓度条件下,CaCl2作用时,hmsT转录水平上升,蛋白水平也提高。37℃时,hmsT转录水平上升,但是蛋白水平降低到了0.52倍。21℃时hmsT转录水平降低,但是蛋白水平上升到1.41倍。hmsT表达受到温度及渗透压的转录后水平调控。而hmsP在NaCl、37℃、pH5、DTT条件下时,转录水平上升,但是蛋白水平没有受到显著影响,说明这些条件是转录后水平的调控作用,SDS作用时转录水平下降,蛋白水平也下降。hmsCDE在FeCl2和NaCl作用时,转录水平降低,HmsD蛋白水平降低。DTT作用时转录水平上升,HmsD蛋白水平也上升,hmsD表达受到转录水平的调控。但是在CaCl2、MgCl2、CuSO4、21℃条件下时,hmsC表达水平上升,FeCl2、NaCl、蔗糖、pH5、37℃作用时,表达水平下降。CaCl2、NaCl、蔗糖、pH5、37℃作用时,hmsE表达降低。相比较HmsD,HmsC和HmsE在转录后水平受到更多因素的调控。不同的环境条件调节三种代谢酶的表达水平,调控c-di-GMP代谢,进而精细调控调控生物被膜形成[70]。
5 展望
鼠疫菌可在跳蚤消化道定殖并在前胃形成生物被膜,这一特性对于其以跳蚤作为媒介的高效传播具有关键的作用。越来越多的研究证明了c-di-GMP在鼠疫菌生物被膜形成的调控网络中的核心作用。鼠疫菌生物被膜形成及c-di-GMP代谢调控通路的研究已经取得了大量进展,相关的研究有利于揭示鼠疫预防和治疗新的靶点。然而c-di-GMP调节网络包含了4个构件:接受环境信号及细胞内信号的DGC和 PDE、c-di-GMP直接结合并别构激活的效应分子以及效应分子作用的靶点,最终产生调节作用[24]。文章中描述了多种调节因子对c-di-GMP代谢及鼠疫菌生物被膜形成相关调节基因的表达调控作用,但是c-di-GMP受体却还未有任何报道,还有待更多的研究去发现,而其他细菌的c-di-GMP受体已经有了一些发现,如多种细菌所含的PilZ结构域,铜绿假单胞菌的转录因子FleQ[71]。另一方面,作用于c-di-GMP调节网络的环境信号及细胞内信号以及hmsHFRS、hmsT、hmsD和hmsP受到的更具体的表达调控机制还需要更多的研究去填补,以丰富鼠疫菌传播及致病的知识信息网络。病原体的传播方式与传染病大暴发的条件息息相关,故而相关的机制阐明均能在一定程度上有益于鼠疫的预防和控制研究。
[1]Bacot AW.LXXXI.Further notes on the mechanism of the transmission of plague by fleas.J Hyg,1915,14(S):774–776.3.
[2]Römling U, Amikam D.Cyclic di-GMP as a second messenger.Curr Opin Microbiol,2006,9(2):218–228.
[3]Costerton JW, Geesey GG, Cheng KJ.How bacteria stick.Sci Am,1978,238(1):86–95.
[4]Böckelmann U, Janke A, Kuhn R, et al.Bacterial extracellular DNA forming a defined network-like structure.FEMS Microbiol Lett,2006,262(1):31–38.
[5]Jennings LK, Storek KM, Ledvina HE, et al.Pel is a cationic exopolysaccharide that cross-links extracellular DNA in thePseudomonas aeruginosabiofilm matrix.Proc Natl Acad Sci USA,2015,112(36):11353–11358.
[6]West SA, Griffin AS, Gardner A, et al.Social evolution theory for microorganisms.Nat Rev Microbiol,2006,4(8):597–607.
[7]Flemming HC, Wingender J, Szewzyk U, et al.Biofilms: an emergent form of bacterial life.Nat Rev Microbiol,2016,14(9):563–575.
[8]Walters III MC, Roe F, Bugnicourt A, et al.Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance ofPseudomonasaeruginosabiofilms to ciprofloxacin and tobramycin.Antimicrob Agents Chemother,2003,47(1):317–323.
[9]Choi CQ.Bug vs.superbug.Hungry microbes can hunt down drug-resistant superbugs.Sci Am,2013,309(2):24.
[10]Rasmussen S, Allentoft ME, Nielsen K, et al.Early divergent strains ofYersiniapestisin Eurasia5000 years ago.Cell,2015,163(3):571–582.
[11]Perry RD, Fetherston JD.Yersiniapestis——etiologic agent of plague.Clin Microbiol Rev,1997,10(1):35–66.
[12]Erickson DL, Jarrett CO, Wren BW, et al.Serotype differences and lack of biofilm formation characterizeYersiniapseudotuberculosisinfection of theXenopsyllacheopisfleavector ofYersinia pestis.J Bacteriol,2006,188(3):1113–1119.
[13]Sun YC, Jarrett CO, Bosio CF, et al.Retracing the evolutionary path that led to flea-borne transmission ofYersiniapestis.Cell Host Microbe,2014,15(5):578–586.
[14]Lorange EA, Race BL, Sebbane F, et al.Poor vector competence of fleas and the evolution of hypervirulence inYersiniapestis.J Infect Dis,2005,191(11):1907–1912.
[15]Hinnebusch BJ, Perry RD, Schwan TG.Role of theYersiniapestishemin storage(hms)locus in the transmission of plague by fleas.Science,1996,273(5273):367–370.
[16]Hinnebusch BJ, Erickson DL.Yersiniapestisbiofilm in the flea vector and its role in the transmission of plague.In: Romeo T, Ed.BacterialBiofilms.Berlin Heidelberg: Springer,2008:229–248.
[17]Jarrett CO, Deak E, Isherwood KE, et al.Transmission ofYersiniapestisfrom an infectious biofilm in the flea vector.J Infect Dis,2004,190(4):782–792.
[18]Abu Khweek A, Fetherston JD, Perry RD.Analysis of HmsH and its role in plague biofilm formation.Microbiology,2010,156(5):1424–1438.
[19]Bobrov AG, Kirillina O, Forman S, et al.Insights intoYersiniapestisbiofilm development: topology and co-interaction of Hms inner membrane proteins involved in exopolysaccharide production.Environ Microbiol,2008,10(6):1419–1432.
[20]Darby C.Uniquely insidious:Yersiniapestisbiofilms.Trends Microbiol,2008,16(4):158–164.
[21]Sun YC, Koumoutsi A, Darby C.The response regulator PhoP negatively regulatesYersinia pseudotuberculosisandYersiniapestisbiofilms.FEMS Microbiol Lett,2009,290(1):85–90.
[22]Duerig A, Abel S, Folcher M, et al.Second messenger-mediated spatiotemporal control of protein degradation regulates bacterial cell cycle progression.Genes Dev,2009,23(1):93–104.
[23]McDougald D, Rice SA, Barraud N, et al.Should we stay or should we go: mechanisms and ecological consequences for biofilm dispersal.Nat Rev Microbiol,2011,10(1):39–50.
[24]Hengge R.Principles ofc-di-GMP signalling in bacteria.Nat Rev Microbiol,2009,7(4):263–273.
[25]Ross P, Weinhouse H, Aloni Y, et al.Regulation of cellulose synthesis inAcetobacterxylinumby cyclic diguanylic acid.Nature,1987,325(6101):279–281.
[26]Malone JG, Williams R, Christen M, et al.The structure-function relationship of WspR, aPseudomonasfluorescensresponse regulator with a GGDEF output domain.Microbiology,2007,153(4):980–994.
[27]Christen B, Christen M, Paul R, et al.Allosteric control of cyclic di-GMP signaling.J Biol Chem,2006,281(42):32015–32024.
[28]Seshasayee ASN, Fraser GM, Luscombe NM.Comparative genomics of cyclic-di-GMP signalling in bacteria: post-translational regulation and catalytic activity.Nucleic Acids Res,2010,38(18):5970–5981.
[29]Ryan RP, Fouhy Y, Lucey JF, et al.Cell-cell signaling inXanthomonascampestrisinvolves an HD-GYP domain protein that functions in cyclic di-GMP turnover.Proc Natl Acad Sci USA,2006,103(17):6712–6717.
[30]Krasteva PV, Giglio KM, Sondermann H.Sensing the messenger: the diverse ways that bacteria signal throughc-di-GMP.Protein Sci,2012,21(7):929–948.
[31]Wolfe AJ, Visick KL.Get the message out:cyclic-Di-GMP regulates multiple levels of flagellum-based motility.J Bacteriol,2008,190(2):463–475.
[32]Bobrov AG, Kirillina O, Ryjenkov DA, et al.Systematic analysis of cyclic di-GMP signalling enzymes and their role in biofilm formation and virulence inYersiniapestis.Mol Microbiol,2011,79(2):533–551.
[33]Sun YC, Koumoutsi A, Jarrett C, et al.Differential control ofYersiniapestisbiofilm formationin vitroand in the flea vector by twoc-di-GMP diguanylate cyclases.PLoS ONE,2011,6(4):e19267.
[34]Jones HA, Lillard JW Jr, Perry RD.HmsT, a protein essential for expression of the haemin storage(Hms+)phenotype ofYersiniapestis.Microbiology,1999,145(8):2117–2128.
[35]Kirillina O, Fetherston JD, Bobrov AG, et al.HmsP, a putative phosphodiesterase, and HmsT, a putative diguanylate cyclase, control Hms-dependent biofilm formation inYersiniapestis.Mol Microbiol,2004,54(1):75–88.
[36]Wortham BW, Oliveira MA, Fetherston JD, et al.Polyamines are required for the expression of key Hms proteins important forYersiniapestisbiofilm formation.Environ Microbiol,2010,12(7):2034–2047.
[37]Bobrov AG, Kirillina O, Perry RD.The phosphodiesterase activity of the HmsP EAL domain is required for negative regulation of biofilm formation inYersiniapestis.FEMS Microbiol Lett,2005,247(2):123–130.
[38]Ren GX, Yan HQ, Zhu H, et al.HmsC, a periplasmic protein, controls biofilm formation via repression of HmsD, a diguanylate cyclase inYersiniapestis.Environ Microbiol,2014,16(4):1202–1216.
[39]Bobrov AG, Kirillina O, Vadyvaloo V, et al.TheYersiniapestisHmsCDE regulatory system is essential for blockage of the oriental rat flea(Xenopsyllacheopis), a classic plague vector.Environ Microbiol,2015,17(4):947–959.
[40]Fang N, Yang HY, Fang HH, et al.RcsAB is a major repressor ofYersiniabiofilm development through directly acting onhmsCDE,hmsT, andhmsHFRS.Sci Rep,2015,5:9566.
[41]Majdalani N, Gottesman S.The Rcs phosphorelay:a complex signal transduction system.Annu Rev Microbiol,2005,59(1):379–405.
[42]Huang YH, Ferrieres L, Clarke DJ.The role of the Rcs phosphorelay in Enterobacteriaceae.Res Microbiol,2006,157(3):206–212.
[43]Sun YC, Hinnebusch BJ, Darby C.Experimental evidence for negative selection in the evolution of aYersiniapestispseudogene.Proc Natl Acad Sci USA,2008,105(23):8097–8101.
[44]Zhu H, Mao XJ, Guo XP, et al.ThehmsT3ʹ untranslated region mediates c-di-GMP metabolism and biofilm formation inYersinia pestis.Mol Microbiol,2016,99(6):1167–1178.
[45]Sun YC, Guo XP, Hinnebusch BJ, et al.TheYersiniapestisRcs phosphorelay inhibits biofilm formation by repressing transcription of the diguanylate cyclase genehmsT.J Bacteriol,2012,194(8):2020–2026.
[46]Guo XP, Ren GX, Zhu H, et al.Differential regulation of the hmsCDE operon inYersinia pestisandYersiniapseudotuberculosisby the Rcs phosphorelay system.Sci Rep,2015,5:8412.
[47]Bellows LE, Koestler BJ, Karaba SM, et al.Hfq-dependent, co-ordinate control of cyclic diguanylate synthesis and catabolism in the plague pathogenYersiniapestis.Mol Microbiol,2012,86(3):661–674.
[48]Rempe KA, Hinz AK, Vadyvaloo V.Hfq regulates biofilm gut blockage that facilitates flea-borne transmission ofYersiniapestis.J Bacteriol,2012,194(8):2036–2040.
[49]Groisman EA.The pleiotropic two-component regulatory system PhoP-PhoQ.J Bacteriol,2001,183(6):1835–1842.
[50]Rebeil R, Jarrett CO, Driver JD, et al.Induction of theYersiniapestisPhoP-PhoQ regulatory system in the flea and its role in producing a transmissible infection.J Bacteriol,2013,195(9):1920–1930.
[51]Vadyvaloo V, Viall AK, Jarrett CO, et al.Role of the PhoP-PhoQ gene regulatory system in adaptation ofYersiniapestisto environmental stress in the flea digestive tract.Microbiology,2015,161(6):1198–1210.
[52]Vadyvaloo V, Hinz AK. A LysR-type transcriptional regulator, RovM, senses nutritional cues suggesting that it is involved in metabolic adaptation ofYersiniapestisto the flea gut.PLoS ONE,2015,10(9): e0137508.
[53]Cathelyn JS, Crosby SD, Lathem WW, et al.RovA,a global regulator ofYersiniapestis, specifically required for bubonic plague.Proc Natl Acad Sci USA,2006,103(36):13514–13519.
[54]Zhang YQ, Gao H, Wang L, et al.Molecular characterization of transcriptional regulation ofrovAby PhoP and RovA inYersiniapestis.PLoS ONE,2011,6(9): e25484.
[55]Tan L, Darby C.Yersiniapestisis viable with endotoxin composed of only lipid A.J Bacteriol,2005,187(18):6599–6600.
[56]Liu L, Fang HH, Yang HY, et al.Reciprocal regulation ofYersiniapestisbiofilm formation and virulence by RovM and RovA.Open Biol,2016,6(3):150198.
[57]Vadyvaloo V, Jarrett C, Sturdevant DE, et al.Transit through the flea vector induces a pretransmission innate immunity resistance phenotype inYersiniapestis.PLoS Pathog,2010,6(2): e1000783.
[58]Zhou DS, Qin L, Han YP, et al.Global analysis of iron assimilation and fur regulation inYersinia pestis.FEMS Microbiol Lett,2006,258(1):9–17.
[59]Gao H, Zhou DS, Li YL, et al.The iron-responsive Fur regulon inYersiniapestis.J Bacteriol,2008,190(8):3063–3075.
[60]Sun FJ, Gao H, Zhang YQ, et al.Fur is a repressor of biofilm formation inYersiniapestis.PLoS ONE,2012,7(12): e52392.
[61]Qu Y, Bi LJ, Ji XL, et al.Identification by cDNA cloning of abundant sRNAs in a human-avirulentYersiniapestisstrain grown under five different growth conditions.Future Microbiol,2012,7(4):535–547.
[62]Beauregard A, Smith E, Petrone B, et al.Identification and characterization of small RNAs inYersiniapestis.RNA Biol,2013,10(3):397–405.
[63]Yan YF, Su SC, Meng XR, et al.Determination of sRNA expressions by RNA-seq inYersiniapestisgrowninvitroand during infection.PLoS ONE,2013,8(9): e74495.
[64]Schiano CA, Koo JT, Schipma MJ, et al.Genome-wide analysis of small RNAs expressed byYersiniapestisidentifies a regulator of the Yop-Ysc type III secretion system.J Bacteriol,2014,196(9):1659–1670.
[65]Fang N, Qu S, Yang HY, et al.HmsB enhances biofilm formation inYersiniapestis.Front Microbiol,2014,5:685.
[66]Liu ZZ, Gao XF, Wang HD, et al.Plasmid pPCP1-derived sRNA HmsA promotes biofilm formation ofYersiniapestis.BMC Microbiol,2016,16:176.
[67]Vogel J, Luisi BF.Hfq and its constellation of RNA.Nat Rev Microbiol,2011,9(8):578–589.
[68]Brennan RG, Link TM.Hfq structure, function and ligand binding.Curr Opin Microbiol,2007,10(2):125–133.
[69]Perry RD, Bobrov AG, Kirillina O, et al.Temperature regulation of the hemin storage(Hms+)phenotype ofYersiniapestisis posttranscriptional.J Bacteriol,2004,186(6):1638–1647.
[70]Ren GX, Fan S, Guo XP, et al.Differential regulation ofc-di-GMP metabolic enzymes by environmental signals modulates biofilm formation inYersiniapestis.Front Microbiol,2016,7:821.
[71]Hickman JW, Harwood CS.Identification of FleQ fromPseudomonasaeruginosaas ac-di-GMP-responsive transcription factor.Mol Microbiol,2008,69(2):376–389.
(本文责编 陈宏宇)
Regulation ofc-di-GMP metabolism and biofilm formation inYersinia pestis
Juping Zhao, and Yicheng Sun
Institute of Pathogen Biology,Chinese Academy of Medical Sciences and Peking Union Medical College,Beijing100730,China
Yersinia pestis,the cause of plague, is transmitted by flea bite.Y.pestisforms a biofilm in the proventriculus of its flea vector to enhance transmission.Biofilm formation inY.pestisis positively regulated by the intracellular levels of the second messenger cyclic diguanylate(c-di-GMP).Thec-di-GMP inY.pestisis synthesized by two diguanylate cyclases(DGC), HmsT and HmsD, and degraded by phosphodiesterase(PDE), HmsP.Here we summarized the regulators that modulatec-di-GMP metabolism and biofilm formation inY.pestisand discussed their regulatory mechanism.
Yersinia pestis, biofilm,c-di-GMP, diguanylate cyclases, phosphodiesterase
February25,2017;Accepted:June5,2017
Yicheng Sun.Tel: +86-10-67837366; E-mail: sunyc@ipbcams.ac.cn

赵菊平, 孙义成.鼠疫耶尔森氏菌环二鸟苷酸代谢及生物被膜形成调控研究进展.生物工程学报,2017,33(9):1513–1524.
Zhao JP, Sun YC.Regulation ofc-di-GMP metabolism and biofilm formation inYersinia pestis.Chin J Biotech,2017,33(9):1513–1524.
Supported by:National Natural Science Foundation of China(No.31670139).
国家自然科学基金(No.31670139)资助。
时间:2017-06-19
http://kns.cnki.net/kcms/detail/11.1998.Q.20170619.1533.001.html
孙义成 博士,副研究员,硕士生导师。先后毕业于北京理工大学和北京大学,美国国家卫生研究院(NIH)过敏及传染病研究所(NIAID)博士后。现任中国医学科学院/北京协和医学院病原生物学研究所课题组组长,主要从事病原菌生物被膜的形成及调控机制研究。作为第一作者或通讯作者在Cell Host & Microbe、Proc Natl Acad Sci USA、Environ Micobiol和Mol Micobiol等杂志发表论文多篇。主持或参与国家自然科学基金面上项目、国家重点研发计划及973计划等多项国家级科研项目。兼任Scientific Reports和Fronti Microbiol的学术编辑。
猜你喜欢
趣味(作文与阅读)(2022年3期)2022-06-10 02:12:48
新民周刊(2019年47期)2019-12-26 21:28:00
少儿美术(快乐历史地理)(2019年8期)2019-12-21 01:50:10
小太阳画报(2019年5期)2019-06-25 10:56:04
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
小主人报(2018年20期)2018-10-25 05:47:34
特别健康(2018年4期)2018-07-03 00:38:20
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
人间(2015年21期)2015-03-11 15:23:07
