
铯胁迫对蚕豆幼苗茎、叶中非结构性碳水化合物(NSC)分配的影响
2017-11-02 01:58马尘扬
种子科技 2017年10期
马尘扬
(成都七中嘉祥外国语学校,四川 成都 610000)
铯胁迫对蚕豆幼苗茎、叶中非结构性碳水化合物(NSC)分配的影响
马尘扬
(成都七中嘉祥外国语学校,四川 成都 610000)
用浓度为0~200 mg·L-1含Cs+溶液浇灌盆栽蚕豆,模拟放射性137Cs随雨水沉降到土壤造成的污染,对蚕豆幼苗生长特性和非结构性碳水化合物(NSC)分配的影响,以探究Cs+胁迫下蚕豆幼苗的碳供应状况。结果显示,不同Cs+浇灌处理对蚕豆发芽率、叶面积无显著影响(p>0.05),也不会导致矮化;蚕豆茎的NSC含量随着Cs+浓度的升高显著降低,降低幅度达26.38%~44.00%,而叶中的NSC无显著性变化。结果表明,本试验设定的Cs+浓度浇灌不会对蚕豆萌发和地上部分生长特性产生明显的毒害作用;蚕豆叶中NSC含量没有发生显著变化,而茎中NSC含量显著降低,可能是Cs+胁迫影响了NSC从“源”到“库”(即从叶到茎)的运输或转化。
铯;非结构性碳水化合物;蚕豆;核素污染
放射性核素在工业、能源、医疗卫生等领域中广泛应用,且可通过降雨等途径沉降到土壤被植物吸收,进入生态系统,甚至危害人类生命健康[1]。土壤放射性核素污染的治理一直是个大难题。近年来,植物修复技术为土壤中放射性核素污染的防治提供了新思路,即利用植物能够忍耐或超量富集某种或某些化学元素的特性清除环境中的重金属或核素污染[2]。
铯的4种同位素中133Cs比较稳定,137Cs具有极强的放射性,137Cs是放射性尘埃和核废料的重要元素之一[3]。核工业或核事故排放的137Cs尘埃可随雨水沉降到土壤,且长期保留,可能会对植物造成毒害。研究表明,植物对137Cs的富集机制与133Cs无显著差异[4],因此,本试验含133Cs的氯化铯溶液作为处理因素模拟137Cs沉降到土壤造成的污染。
蚕豆(Vicia faba Linn)是我国栽培较广的农作物之一,其种子可作为蔬菜、粮食食用,茎、叶可作为饲料或绿肥,地上部分应用价值高,常作为植物对环境胁迫响应研究的模式植物。研究显示,蚕豆对Cs+的蓄积能力表现为根>叶>茎[5],Cs+从根部向地上部分转移较困难[6]。
碳水化合物是植物进行光合作用的主要产物,按其结构可分为结构性碳水化合物(SC)和非结构性碳水化合物(NSC)两类。NSC则主要含可溶性总糖和淀粉,主要参与植物的生命代谢活动,其含量通常反映植物碳代谢的收支平衡,对植物生长发育有重要影响[7]。NSC还能表征植株对环境胁迫的响应,许多环境因素都能影响植物体内NSC含量分配的变化[8]。近年来,关于植物NSC含量分配的研究主要集中于生态系统中NSC季节性变化,群落建成的NSC分配变化等[9,10]和NSC含量分配对营养胁迫[11]、干旱胁迫、温度和CO2胁迫[12]等环境胁迫的响应。当前,从NSC分配变化的角度研究植物对重金属、核素等胁迫的生理响应还鲜有报道。
因此,本试验用不同浓度Cs+浇灌盆栽蚕豆,模拟137Cs随雨水沉降到土壤造成的污染,着眼于蚕豆幼苗地上部分(茎、叶)的生长和NSC含量分配对土壤Cs+胁迫的响应,为用蚕豆修复放射性污染土壤的可行性和安全性提供试验依据。
1 材料与方法
1.1 材料处理与仪器
1.1.1 试验材料
蚕豆购自四川农科种业有限公司。选取饱满的种子,用3%的H2O2浸泡消毒15 min,用蒸馏水冲洗干净,待水分沥干后播到盆中,室温培养。本试验所用土壤理化性质及铯的背景值如表1所示。
1.1.2 主要试剂与仪器
氯化铯(CsCl)、蒽酮(C14H10O)、葡萄糖(C6H12O6)、淀粉[(C6H10O5)n]、浓硫酸(H2SO4)、硝酸(HNO3)、高氯酸(HClO4)等,以上试剂均为AR(分析纯)级。
HI850R型离心机,TAS-990型火焰原子吸收分光光度计,BioMate*3S分光光度计,AR223CN型电子天平(精确至 0.001 g)。

表1 供试土壤理化性质
1.2 试验设计及方法
1.2.1 试验设计
本 研 究 用 浓 度 分 别 为 25 mg·L-1、50 mg·L-1、100 mg·L-1、200 mg·L-1的 Cs+溶液浇灌盆栽蚕豆,模拟Cs+随雨水沉降到土壤造成的污染,以蒸馏水浇灌(Cs+0 mg·L-1)作为对照,每2~3 d浇灌1次,浇灌至播种后40 d,测定蚕豆生长指标,并剪取茎、叶样本测定Cs+含量和NSC含量。
1.2.2 试验方法
用火焰原子吸收光度计法测定蚕豆体内的Cs+含量[13];采用蒽酮比色法测定蚕豆茎、叶的可溶性总糖和总淀粉的含量[14]。
1.3 数据统计与分析
用Excel 2010进行数据统计,用SPSS 18.0进行单因素方差分析(ANOVA)及相关分析,用Origin 9.0做图。
2 结果与分析
2.1 对蚕豆萌发及生长特性的影响
2.1.1 发芽率
处理组(25~200 mg·L-1)与对照组(0 mg·L-1)的发芽率无显著性差异(p>0.05),且均达到90%以上(图1),这表明较高浓度的Cs+沉降到土壤也不会对蚕豆的萌发产生毒害,可能是蚕豆厚厚的种皮保护了胚的萌发。
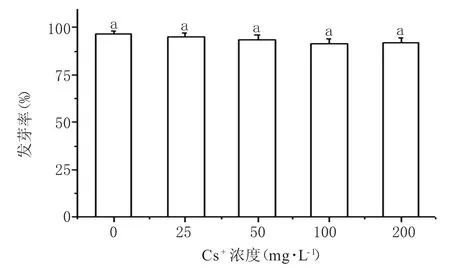
图1 Cs+胁迫对蚕豆发芽率的影响
2.1.2 株高
由图2看出,Cs+处理下蚕豆幼苗株高在一定程度上变高,在浓度为 25 mg·L-1时显著高于对照,但浓度高于50 mg·L-1后与对照差异不显著,表明铯胁迫下蚕豆幼苗没有出现矮化症状。
2.1.3 叶面积
用不同浓度的Cs+浇灌,蚕豆叶面积有所增大(图3),但处理组与对照组并无显著性差异(p>0.05),这表明试验所设定的Cs+浓度(25~200 mg·L-1)范围沉降到土壤中不会对蚕豆叶片外形生长产生显著影响。
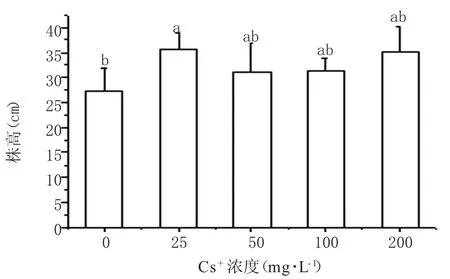
图2 Cs+胁迫对蚕豆苗株高的影响
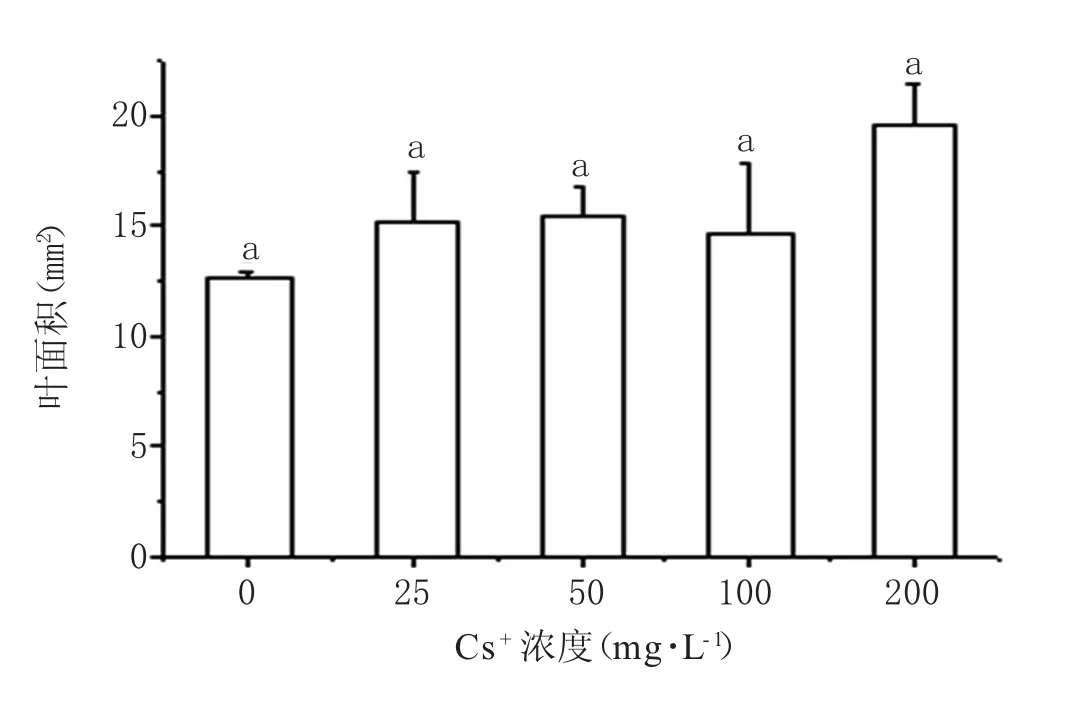
图3 Cs+胁迫对蚕豆叶面积的影响
2.2 蚕豆茎、叶铯富集量比较
随着Cs+浓度的增加,蚕豆茎、叶对Cs+的富集量显著增加(p<0.05),当浓度达到 200 mg·L-1时,蚕豆茎、叶 Cs+富集量最高,分别达到 0.114 mg·g-1DW、0.133 mg·g-1DW(图 4)。还可看出,在相同处理下,蚕豆叶中Cs+含量高于茎中的,在50 mg·L-1处理下,茎、叶中Cs+含量比茎高32.53%,差异达到最大。
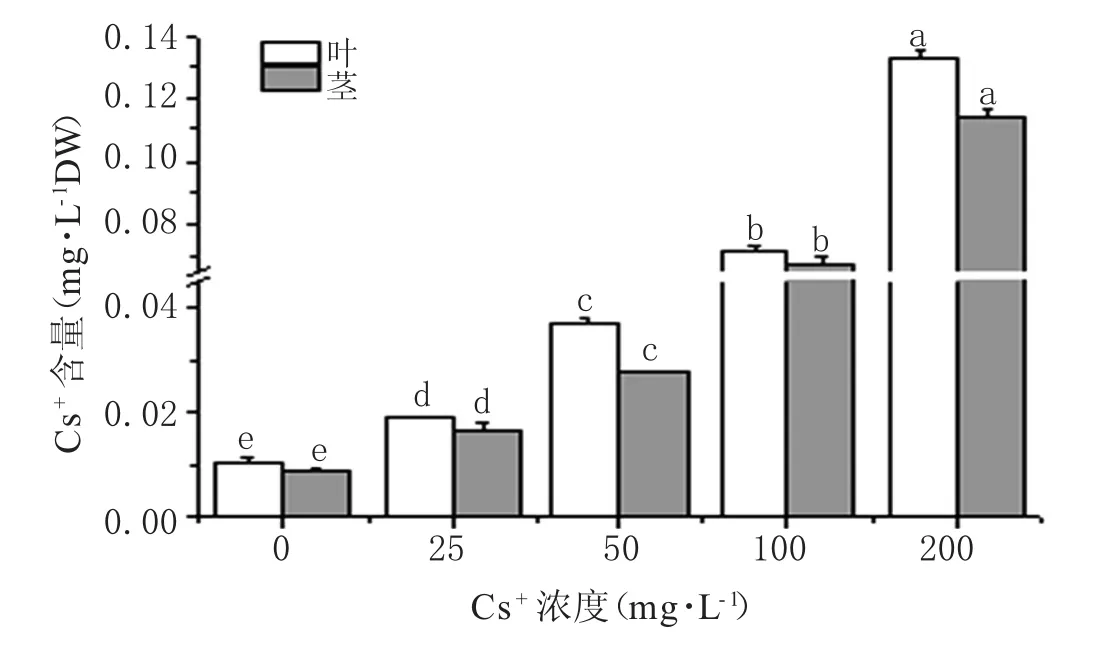
图4 蚕豆茎、叶Cs+富集量
2.3 非结构性碳水化合物在茎、叶中的分配
2.3.1 可溶性总糖
蚕豆叶和茎中可溶性总糖含量随Cs+升高均呈现下降的趋势(图 5)。浓度为 100 mg·L-1、200 mg·L-1处理下,叶中可溶性糖含量与对照组比较下降了35.05%和12.91%;在 25 mg·L-1、100 mg·L-1、200 mg·L-1处理组下,茎中可溶性糖含量分别比对照低45.15%、38.25%、42.39%。在Cs+胁迫下,茎、叶中可溶性糖含量比值更小(图 5b),在浓度为 25 mg·L-1和 200 mg·L-1时特别显著。以上结果表明,Cs+胁迫可引起蚕豆茎、叶中可溶性糖含量降低,且茎的减少幅度更大。
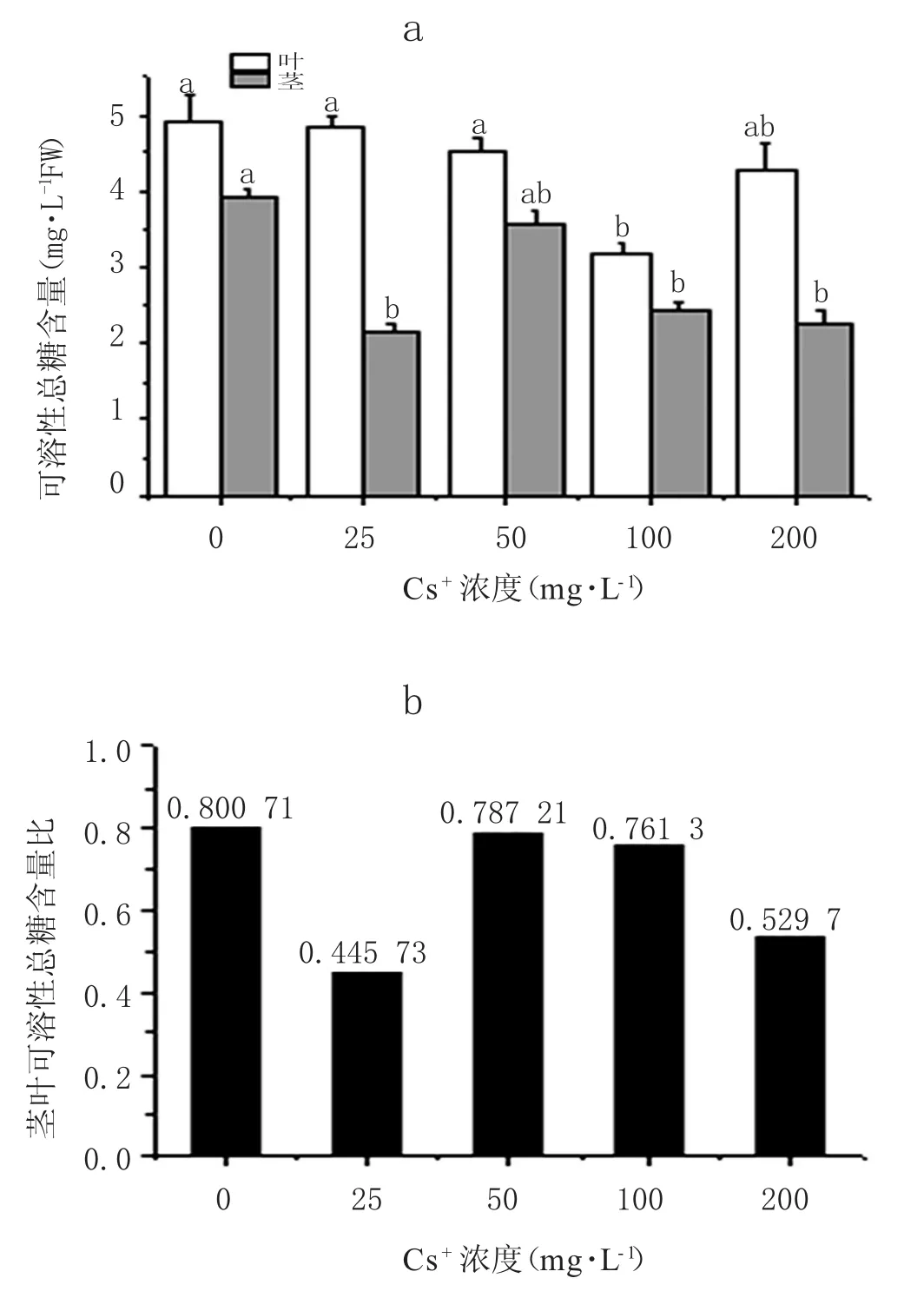
图5 不同Cs+处理下蚕豆茎、叶可溶性总糖含量
2.3.2 淀粉
如图6所示,蚕豆叶中淀粉含量随Cs+浓度的升高变化不显著(p>0.05),而茎中淀粉含量会显著下降(图6a)。在不同浓度 Cs+处理下(25~200 mg·L-1),茎中淀粉含量与对照相比下降幅度为19.95%~43.27%。这表明Cs+胁迫会降低茎中淀粉含量,且会降低茎、叶淀粉含量比值(图6b)。
2.3.3 NSC总量
如图7所示,叶中的NSC总含量随Cs+浓度的升高无显著性变化(p>0.05);在不同浓度Cs+处理下(25~200 mg·L-1),蚕豆茎中NSC含量呈现显著性降低趋势(p<0.05),分别比对照组降低 44.00%、26.38%、30.89%、28.73%。在不同浓度Cs+浇灌后,叶中NSC含量显著高于茎。在浓度为 25 mg·L-1、50 mg·L-1、200 mg·L-1处理下,叶中NSC含量比茎中含量分别高52.97%、19.14%和17.24%,这表明Cs+胁迫可能影响了NSC由叶到茎的转运。
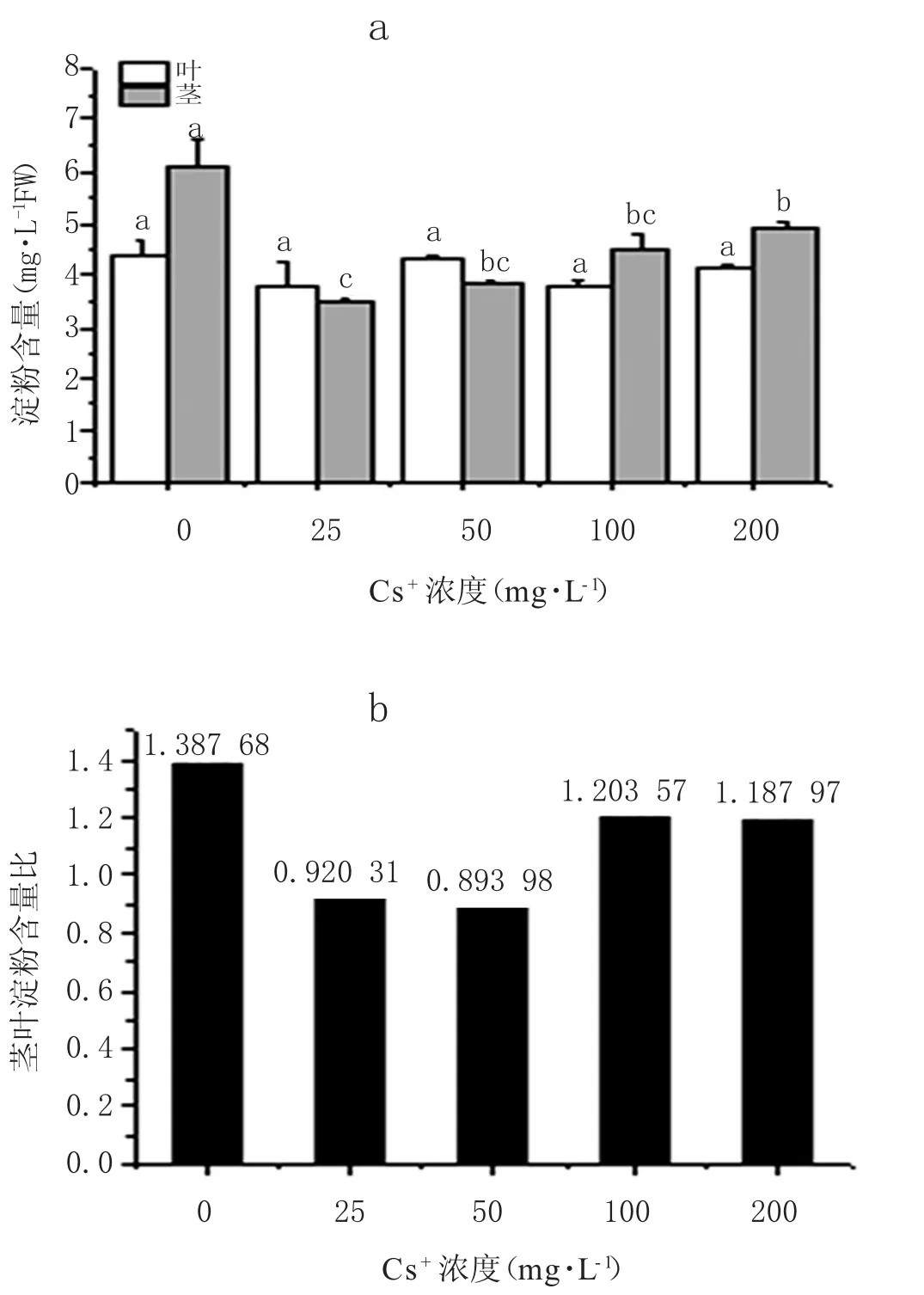
图6 不同Cs+处理下蚕豆茎、叶淀粉含量
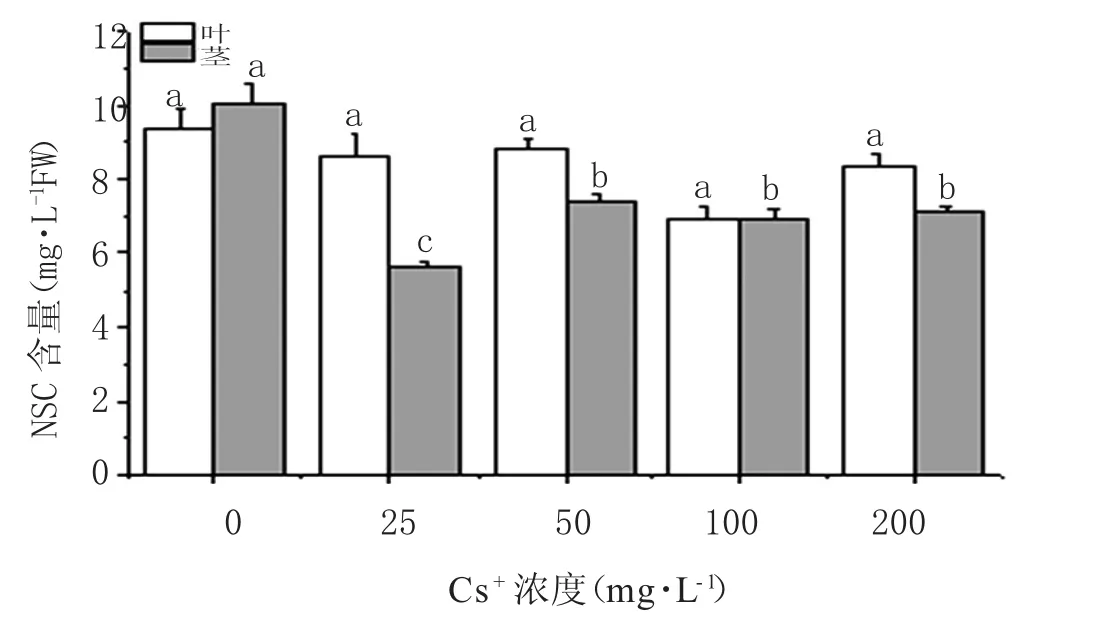
图7 铯胁迫下蚕豆茎、叶NSC含量
2.4CS+富集量与NSC含量的相关性分析
相关性分析显示(表2),在叶和茎中Cs+含量和NSC含量的相关系数分别为-0.359和-0.249,但相关性不显著(p>0.05),表明在Cs+胁迫下两者存在较弱的负相关性。

表2 蚕豆叶、茎铯富集量与NSC含量的pearson相关系数
3 讨论
植物萌发和外观形态对外界环境胁迫有敏感的响应。张红霞等[15]研究显示,用不同强度的UV-B辐射胁迫使蚕豆幼苗矮化达50%以上,叶面积和干物质量显著减少。付倩[16]等发现Sr2+胁迫会显著降低蚕豆的发芽率,根和芽伸长受到明显抑制。
本试验结果显示,用不同浓度Cs+(25~100 mg·L-1)浇灌,不会显著降低蚕豆的发芽率,对幼苗地上部分的生长特性(株高、叶面积)也无显著性影响。这可能是由于蚕豆较厚的种皮保护胚不受Cs+毒害,顺利发芽,土壤环境可改变部分Cs+的化学形态,缓解Cs+对作物的毒害作用[17]。随着Cs+浓度的增加,蚕豆幼苗茎、叶对Cs+的富集量显著增加,但富集量并不高,最高仅分别为0.114 mg·g-1DW、0.133 mg·g-1DW;在同一浓度 Cs+处理下,叶中Cs+含量高于茎。
有证据表明,蚕豆根对Cs+的富集能力远远高出茎和叶,根细胞可通过阻滞作用将Cs+固定在细胞壁上[18],在一定程度上降低Cs+对茎、叶的毒害作用,这也能很好解释本试验结果显示的蚕豆地上部分生长特性对Cs+胁迫响应不显著的结论。
植物常会打破其体内NSC含量的分配格局来对环境胁迫作出响应。可溶性糖和淀粉是植物NSC的主要成分,可溶性总糖主要由叶光合作用产生,转运到各器官;淀粉则是光合产物在植物体内重要的储存物质,两者可以相互转化,以应对外界胁迫[19]。可溶性总糖和淀粉含量因功能和生理意义不同在植物不同器官的分配不同。一般情况下,叶片作为NSC的“源”,其可溶性糖含量高于根、茎;而茎的淀粉含量高于叶片。可溶性糖含量和淀粉含量对外界环境的响应也有可能不同。
在本研究中,蚕豆茎、叶可溶性糖含量均随Cs+浓度的升高呈现下降趋势,但茎中含量下降幅度更大;叶中淀粉含量随Cs+浓度的升高变化不显著,而茎中淀粉含量大幅下降。蚕豆叶中NSC总量并无显著性变化,而茎NSC总量则显著性降低,Cs+富集量与NSC含量呈现微弱的负相关性。
综上所述,在本试验所设定的Cs+胁迫范围内没有对蚕豆萌发和地上部分生长特性产生明显的毒害作用。Cs+胁迫下,叶中NSC含量无显著变化,茎中则显著减少,这可能是Cs+胁迫改变了NSC从“源”到“库”(即叶到茎)的运输或转化,改变NSC在植物体内的分配格局,以提高对Cs+的耐受性。
[1]张琼,陈金融,张春明,等.土壤放射性核素铯污染修复研究进展[J].核农学报,2014,28(10):1 924-1 931.
[2]赵娜.螯合剂和植物联合修复Pb污染土壤的效果研究[D].保定:河北农业大学,2011.
[3]尹显和,任剑锋,华建平.放射性核素铯-137在土壤里不同质地中的分布规律[J].南华大学学报,2001,15(03):49-51.
[4]Tsukad H,Hasegawa H,Hisamatsu S,et al.Transfer of 137Cs and stable Cs from paddy soil to polished rice in Aomori,Japan.[J].Journal of Environmental Radioactivity,2002,59(03):351-363.
[5]张晓雪,王丹,张志伟,等.水培条件下十种植物对88Sr和133Cs的吸收和富集[J].北方园艺,2009(10):65-67.
[6]张晓雪,王丹,李卫锋,等.133Cs和88Sr在蚕豆苗中的蓄积及其辐射损伤效应[J].辐射研究与辐射工艺学报,2010,28(01):48-52.
[7]潘庆民,韩兴国,白永飞,等.植物非结构性贮藏碳水化合物的生理生态学研究进展[J].植物学报,2002,19(01):30-38.
[8]刘万德,苏建荣,李帅锋,等.云南普洱季风常绿阔叶林主要树种非结构性碳水化合物变异分析[J].林业科学,2017,53(06):1-9.
[9]史建伟,张育平,王孟本,等.植物体内非结构性碳水化合物变化及其影响因素[J].湖北农业科学,2008,47(01):112-115.
[10]于丽敏,王传宽,王兴昌.三种温带树种非结构性碳水化合物的分配[J].植物生态学报,2011,35(12):1 245-1 255.
[11]吕茹冰,杜莹,鲍永新,等.氮沉降对毛竹非结构性碳组成与分配的影响[J].生态学杂志,2017,36(03):584-591.
[12]肖列,刘国彬,李鹏,等.白羊草光合特性及非结构性碳水化合物含量对CO2浓度倍增和干旱胁迫的响应[J].植物营养与肥料学报,2017,23(02):389-397.
[13]赖金龙,付倩,陶宗娅,等.3种农作物可食用部分对铯的蓄积特性[J].食品科学,2016,37(09):87-91.
[14]汤绍虎,罗充.植物生理学试验教程[M].重庆:西南师范大学出版社,2012.
[15]张红霞,吴能表,胡丽涛,等.不同强度UV-B辐射胁迫对蚕豆幼苗生长及叶绿素荧光特性的影响[J].西南师范大学学报:自然科学版,2010,35(01):105-110.
[16]付倩,刘聪,赖金龙,等.锶对蚕豆根尖细胞的遗传毒性效应[J].农业环境科学学报,2015,34(09):1 646-1 652.
[17]崔妍,丁永生,公维民,等.土壤中重金属化学形态与植物吸收的关系[J].大连海事大学学报,2005(02):59-63.
[18]付倩,杨垒滟,赖金龙,等.蚕豆对铯的吸收蓄积及亚细胞分布研究[J].生态毒理学报,2015,10(06):297-304.
[19]Latt C R,Nair P K R,Kang B T.Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate[J].Forest Ecology&Management,2001,146(1-3):145-158.
1005-2690(2017)10-0151-04
X172
B
2017-09-20)
猜你喜欢
中国调味品(2022年3期)2022-03-17
中草药(2022年3期)2022-02-14
农家科技中旬版(2021年6期)2021-07-12
小读者(2021年2期)2021-03-29
少儿科技(2021年8期)2021-01-02
婚姻与家庭·性情读本(2019年11期)2019-11-18
保健与生活(2019年7期)2019-07-31
环球时报(2019-01-03)2019-01-03
分析化学(2018年4期)2018-11-02
学生天地(2018年9期)2018-09-06
