
新疆褐牛CD46基因的原核表达及其多克隆抗体的制备
2017-10-11 08:23木哈买提江铁格斯张慧敏杜润慈加尔肯苏战强冉多良刘建华
动物医学进展 2017年8期
木哈买提江·铁格斯,张慧敏,杜润慈,加尔肯,苏战强,冉多良,刘建华
(新疆农业大学动物医学学院,新疆乌鲁木齐 830052)
新疆褐牛CD46基因的原核表达及其多克隆抗体的制备
木哈买提江·铁格斯1,张慧敏,杜润慈,加尔肯,苏战强,冉多良,刘建华*
(新疆农业大学动物医学学院,新疆乌鲁木齐 830052)
旨在克隆并表达新疆褐牛CD46基因,制备其多克隆抗体,为进一步研究牛CD46分子生物学功能奠定基础。采用RT-PCR方法从新疆褐牛脾脏中扩增CD46基因全长,进行测序,并对测序结果进行生物信息学分析,将CD46部分序列亚克隆于pET-28a(+)和pVAX1载体中。将阳性重组质粒pET-28a-△CD46转化于E.coliRosetta-gamiB(DE3)感受态细胞,诱导表达截短CD46蛋白。用切胶纯化的His-△CD46融合蛋白免疫新西兰大白兔,制备多克隆抗体;采用ELISA测定多克隆抗体的效价;取无内毒素重组质粒pVAX1-△CD46转入BHK-21(仓鼠肾细胞)细胞,表达产物以制备的多克隆抗体为一抗,采用Western blot法检测该多克隆抗体的特异性。CD46基因的测序结果表明,部分基因蛋白表达、纯化条带大小与预期一致;制备的抗牛CD46多克隆抗体效价高于1∶128 000,并具有良好的特异性。
新疆褐牛;CD46;表达;融合蛋白;多克隆抗体
CD46又称为膜辅助蛋白(membrane cofactor protein,MCP),是一类表达于所有有核细胞的Ⅰ型跨膜糖蛋白。CD46主要由4个串联的能够结合C3b和C4b的补体控制蛋白模式(CCP1-CCP4),1个富含丝氨酸、苏氨酸和脯氨酸残基的O糖基化结构域(STP),1个跨膜结构域和1个C端细胞浆尾构成[1]。CD46的主要功能是作为一种辅助因子,促进蛋白酶因子Ⅰ介导的C3b和C4b降解,并增强C3转化酶的活性(尤其是替代途径的C3转化酶),被认为是补体旁路激活途径最有效的抑制因子。因此,CD46通过抑制C3分子激活的联级反应,调节补体的功能,保护宿主自身的正常细胞免遭补体介导的溶解破环[2]。此外,CD46还是许多病毒、细菌等病原体的受体,可以通过胞外结构域与这些病原体特异性结合激发某些生物学或免疫学效应。有研究显示,麻疹病毒、第6型人疱疹病毒与CD46的相互作用能够抑制巨噬细胞IL-12的分泌(IL-12是一种Th分化及NK、T细胞活化必须的细胞因子),并且下调维持细胞内环境稳定的信使分子NO的合成[3]。Price J D等[4]研究表明,链球菌与CD46的相互作用能够诱导人CD4+T细胞分泌大量免疫抑制性细胞因子IL-10并促进调节性T细胞的分化。Li K等[5]研究显示,大肠埃希菌能够通过C3b与CD46的结合感染人的肾小管上皮细胞引发肾盂肾炎。Maurer K等[6]和Krey T等[7]研究发现,抗CD46单抗和CD46分子封闭均能有效抑制BVDV感染细胞,说明CD46是BVDV的细胞膜受体。随后的研究进一步显示,BVDV特异性地吸附CD46能够通过网格蛋白介导的内吞作用以pH依赖方式启动病毒粒子的细胞穿入,引起病毒感染[8]。CD46无论是对正常细胞功能的维持还是在病原的入侵过程中都起到了至关重要的作用。
表达CD46并制备其多克隆抗体对于基因的功能研究将起重要作用,免疫组化、蛋白质相互作用的确定及阻断病原体感染细胞等许多工作的开展也都需要有特异的抗体作为辅助[9]。本试验首次从新疆褐牛脾脏中成功扩增CD46基因全长,进行序列测序,并对测序结果进行生物信息学分析,通过原核表达系统表达截短的CD46蛋白,利用纯化后的重组蛋白免疫新西兰大白兔制备多克隆抗体,为进一步研究牛CD46分子的生物学功能奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒、动物组织和试验用动物E.coliDH5α、E.coliRosetta-gamiB(DE3)、原核表达载体pET-28a (+)、真核表达载体pVAX1、pGM-T、BHK-21细胞,新疆农业大学动物医学学院传染病实验室保存;新疆褐牛脾脏组织采自乌鲁木齐华凌畜牧屠宰场;2只体重均为1.8 kg~2.5 kg的新西兰大白兔由新疆维尔自治区疾病防控中心实验动物中心提供。
1.1.2 主要试剂BamHⅠ和XhoⅠ限制性内切酶、Taq酶、dNTP、T4 DNA连接酶、DNA 标准 DL 2 000、DNA 标准 DL 5 000、DNA 标准 DL 15 000,宝生物工程(大连)有限公司产品;cDNA第1链合成试剂盒,北京全式金生物技术有限公司产品;Trizol、LipofectamineTM 2000,Invitrogen公司产品;弗氏完全佐剂和弗氏不完全佐剂均,Sigma公司产品;OPTI-MEM、DMEM培养基,Hyclone公司产品;胎牛血清,Gibco公司产品;胶回收试剂盒、无内毒素小提质粒试剂盒,OMEGA公司产品;细胞培养瓶、细胞培养用6孔板均,NUNC公司产品;Goat Anti-Rabbit IgG(H+L)-HRP antibody,武汉三鹰生物技术有限公司产品。
1.2 方法
1.2.1 新疆褐牛CD46基因的扩增和TA克隆 根据GenBank数据库中牛CD46基因序列(登录号NM-001242561.1),使用Primer Premier 5.0设计1对特异性引物,序列见表1,引物由上海生工生物工程技术服务有限公司合成。
取脾脏组织于液氮中磨碎,采用Trizol试剂抽提细胞总RNA,经cDNA第1条链合成试剂盒将其反转录成cDNA;以cDNA为模板,P1/P2为特异性引物,PCR扩增CD46基因。反应参数为:95 ℃ 5 min;95 ℃ 45 s,56 ℃ 45 s,72 ℃ 1 min,共35个循环;72 ℃ 10 min。采用DNA凝胶回收试剂盒回收目的基因,连接于pGM-T,并转化E.coliDH5α感受态细胞,涂布于含氨苄青霉素的LB固体培养基,于37 ℃过夜培养,挑取单菌落摇菌并鉴定,将鉴定为阳性的重组质粒命名为pGM-T-CD46。

表1 引物和序列
1.2.2 原核表达质粒pET-28a-△CD46与真核表达质粒pVAX1-△CD46的构建 分别采用DNA Man、Blast、SignalP、TMHMM和Protean软件对CD46基因的测序结果及其编码蛋白的信号肽、跨膜区、抗原性进行分析。选择CD46基因编码蛋白抗原性较高的胞质区为扩增区域,采用Primer Premier 5.0软件设计两对特异性引物P3/P4、P5/P6,序列见表2,预期大小为756 bp,引物由上海生工生物工程技术服务有限公司合成。
分别以质粒pGM-T-CD46为模板,P3/P4、P5/P6为特异性引物,扩增目的基因部分片段△CD46,用DNA胶回收试剂盒回收目的片段。双酶切PCR产物及空载体pET-28a(+)和pVAX1,回收纯化。分别取酶切载体、目的片段于16℃过夜连接,转化E.coliDH5α感受态细胞,涂布于含卡那霉素或氨苄青霉素的LB固体培养基,37℃过夜培养,挑取单菌落并鉴定,鉴定为阳性的重组质粒送上海生工生物工程技术服务有限公司测序。测序正确的质粒分别命名为pET-28a-△CD46和pVAX1-△CD46。

表2 引物和序列
1.2.3 His-△CD46融合蛋白的诱导表达 取阳性重组质粒pET-28a-△CD46,转化E.coliRosetta-gamiB(DE3)感受态细胞,经IPTG 诱导融合蛋白His-△CD46表达;采用SDS-PAGE切胶纯化方法纯化融合蛋白,经Brand Ford 蛋白含量测定试剂盒测定蛋白浓度。
1.2.4 兔抗His-△CD46融合蛋白多克隆抗体的制备 取融合蛋白与弗氏完全佐剂等体积混合,乳化完全后背部皮下多点注射免疫新西兰大白兔,剂量为1 mg/只。以后每间隔2周以弗氏不完全佐剂与融合蛋白完全乳化后进行再次免疫。四免后7 d心脏采血,分离血清保存待检。
1.2.5 抗体效价测定 以纯化的His-△CD46融合蛋白为包被抗原,4 ℃过夜包被ELISA板(0.2 μg/孔),PBST洗涤3次(5 min/次,下同)后用50 g/L的脱脂奶粉37 ℃封闭2 h,PBST洗涤3次,以兔抗His-△CD46融合蛋白多克隆抗体为一抗(1∶1 000~1∶128 000),37 ℃孵育1 h ,PBST洗涤3次,加入Goat Anti-Rabbit IgG(H+L)-HRP antibody(1∶8 000),37 ℃孵育1 h ,PBST洗涤3次后加入TMB显色底物,显色10 min后加入终止液,酶标仪检测OD值。当试验组OD值大于或等于2 倍阴性对照组(免疫前血清)时判为阳性,以阳性血清最大稀释倍数的倒数为该抗体的效价。
1.2.6 抗体特异性检测 取无内毒素重组质粒pVAX1-△CD46和空载体pVAX1分别转入BHK-21细胞,48 h后制备细胞总蛋白,以制备的兔抗His-△CD46融合蛋白多克隆抗体为一抗,Goat Anti-Rabbit IgG(H+L)-HRP antibody为二抗,采用Western blot法检测该多克隆抗体的特异性。
2 结果
2.1 CD46基因的扩增和TA克隆
以新疆褐牛脾细胞总RNA为模板,用特异性引物P1/P2进行CD46基因的RT- PCR扩增,并进行TA克隆,构建重组质粒pGM-T-CD46。RT-PCR扩增产物经10 g/L琼脂糖凝胶电泳可观察到1条约1 257 bp大小的清晰条带,与预期大小相符;pGM-T-CD46的BamHⅠ/XhoⅠ双酶产物经10 g/L琼脂糖凝胶电泳后得到两条分别为1 257 bp、3 000 bp的清晰条带,与预期结果符合(图1);测序结果显示,阳性质粒中目的基因序列与参考序列同源性为100%。

M.DNA 标准 DL 5 000;1.pGM-T-CD46的BamHⅠ/XhoⅠ双酶切产物;2.pGM-T空载体;3.CD46基因RT-PCR扩增产物
M.DNA Marker DL 5 000;1.Enzyme digestion products of recombinant plasmid pGM-T-CD46 byBamHⅠ/XhoⅠ;2.Empty plasmid pGM-T;3.RT-PCR product of CD46
图1重组质粒pGM-T-CD46鉴定结果
Fig.1 Identification of recombinant plasmid pGM-T-CD46
2.2 原核表达质粒pET-28a-△CD46和真核表达质粒pVAX1-△CD46的构建
2.2.1 CD46基因生物信息学分析 采用ORF Finder对CD46基因全序列分析发现,1 bp~1 257 bp是个大的开放阅读框。用Protean软件对CD46基因全序列(1257 bp)分析显示,155-233 aa亲水性较强,易体外表达。1-25、89-118、389-400、406-419 aa抗原性较强,易形成潜在的抗原表位;CD46分子表面可及性较好,该蛋白能被抗体结合的可能性高。使用TMHMM软件预测其跨膜区,该蛋白的1-365 aa位于细胞膜外,366-388 aa之间形成一个典型的跨膜螺旋区,389-418 aa位于细胞膜内。用Signalp软件预测发现该基因编码的蛋白具有信号肽序列,最可能的剪切点位于42-43 aa之间。2.2.2 原核表达载体pET-28a-△CD46与真核表达载体pVAX1-△CD46的鉴定结果 重组质粒pVAX1-△CD46(图2A)和pET-28a-△CD46(图2B)分别进行PCR扩增和BamHⅠ/XhoⅠ双酶切鉴定,产物经10 g/L琼脂糖凝胶电泳后得到与预期结果符合的清晰条带,分别为756、3 000、5 200 bp。测序结果显示,阳性重组质粒中目的基因序列与原始序列同源性为100%,说明成功构建了CD46基因原核及真核表达载体。
2.3 His-△CD46融合蛋白的表达检测及纯化
将重组质粒pET-28a-△CD46转化至受体菌E.coliRosetta-gamiB(DE3)中,经IPTG诱导后SDS- PAGE分析,在37℃,IPTG 浓度为1 mmol/L,诱导3 h,目的基因得到高效表达,目的蛋白主要以包涵体形式存在,其相对分子质量约为35 ku(图3A),与预期大小吻合。利用SDS-PAGE电泳切胶纯化方法获得了高纯度目的蛋白(图3B),其浓度为10.6 mg/mL,可用作免疫抗原。
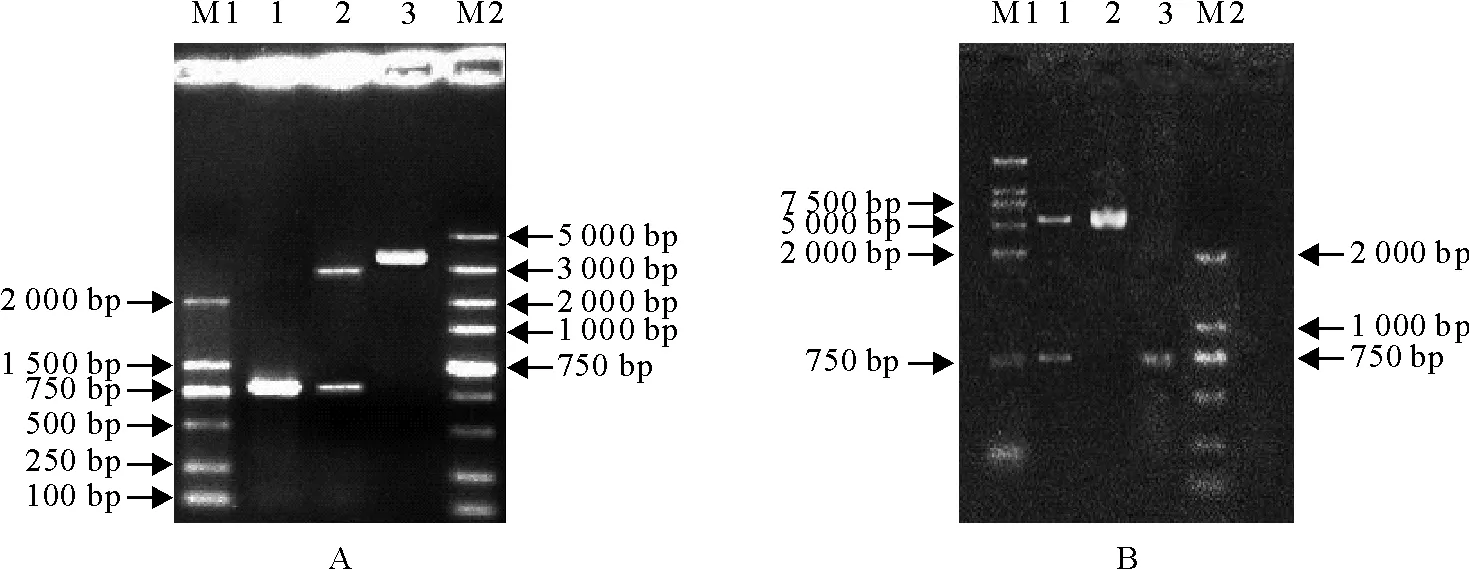
A.M1.DNA 标准 DL 2 000;M2.DNA 标准 DL 5 000;1.pVAX1-△CD46 PCR产物;2.pVAX1-△CD46双酶切产物(BamHⅠ/XhoⅠ);3.pVAX1-△CD46
B.M1.DNA 标准 DL 15 000;M2.DNA 标准 DL 2 000;1.pET-28a-△CD46双酶切产物(BamHⅠ/XhoⅠ);2.pET-28a空载体;3.pET-28a-△CD46 PCR产物
A.M1.DNA Marker DL 2 000;M2.DNA Marker DL 5 000;1.PCR product of pVAX1-△CD46;2.Enzyme digestion products of pVAX1-△CD46 byBamHⅠ/XhoⅠ;3.pVAX1-△CD46
B.M1.DNA Marker DL 15 000;M2.DNA Marker DL 2 000;1.Enzyme digestion products of pET-28a-△CD46 byBamHⅠ/XhoⅠ;2.Empty plasmid pET-28a;3.PCR product of pET-28a-△CD46
图2重组质粒pET-28a-△CD46和pVAX1-△CD46的PCR和双酶切鉴定结果
Fig.2 PCR and enzyme digestion identification of pET-28a-△CD46 and pVAX1-△CD46 recombinant plasmids

A. M.蛋白分子质量标准;1.菌体裂解后包涵体;2.菌体裂解后上清
B. M.蛋白分子质量标准;1.诱导重组菌总蛋白;2.融合蛋白纯化产物
A. M.Protein molecular weight Marker;1. Inclusions after cracking bacteria;2.Supernatant after cracking bacteria
B. M.Protein molecular weight Marker;1.The induced proteins of recombinant bacteria;2.Purified products of fusion proteins
图3 His-△CD46融合蛋白纯化结果
Fig.3 Purification results of fusion protein His-△CD46
2.4 抗体效价检测
以纯化的His-△CD46融合蛋白为包被抗原,采用4免后7 d的兔血清为一抗,倍比稀释,间接ELISA 法测定多克隆抗体的效价,结果制备的CD46抗体效价高于1∶128 000。
2.5 多克隆抗体特异性分析
取无内毒素重组质粒pVAX1-△CD46和空载体pVAX1分别转入BHK-21细胞,诱导表达后,对产物进行Western blot分析,结果显示,在35 ku左右处出现特异性条带,与预期结果一致(图4),空白对照组未出现任何条带,表明制备的抗CD46多克隆抗体能识别真核表达的CD46蛋白。

M.蛋白分子质量标准;1.pVAX1-△CD46转染组;2.pVAX1空载体转染组
M.Protein molecular weight Marker;1.Transfection group with pVAX1-△CD46;2.Transfection group with pVAX1
图4多克隆抗体的Western blot鉴定
Fig.4 Identification of polyclonal antibodies by Western blot
3 讨论
抗血清为含有某一类具有特异免疫功能的抗体分子的血清,一般为动物被人工注射某类抗原后制备的动物血清,高效价的抗血清用于各种研究工作[10]。免疫血清的质量直接影响试验的特异性和敏感性。免疫产生抗体的特异性很大方面取决于所使用的免疫原的抗原性和纯度[11]。因此,本研究在选择表达区域时,分别采用DNA Man、BLAST、SignalP、TMHMM和Protean软件对CD46基因的测序结果及其编码蛋白的信号肽、跨膜区、抗原性进行分析。选择CD46基因编码蛋白抗原性较高的胞质区为扩增区域,成功构建原核表达载体pET-28a-△CD46和真核表达载体pVAX1-△CD46。
蛋白的表达受到许多因素的制约与影响,本研究采用pET载体系列的pET-28a作为表达载体,该载体利用大肠埃希菌的T7噬菌体为启动子[12],可对各种目的基因进行高效率表达,同时载体含有6×组氨酸标签,简化了表达蛋白的纯化步骤。本研究采用pET-28a载体进行重组蛋白表达,最终获得了成功。
为了获得重组蛋白的高特异性抗体,必须先予抗原纯化。不同的重组蛋白需要采用不同的纯化方式,通常要用到凝胶过滤,离子交换和亲和层析等纯化方法[13]。本研究表达的His-△CD46融合蛋白在试验设计之初拟采用His亲和层析纯化,但是在试验过程中发现纯化的融合蛋白并未达到预期的纯度且纯化步骤烦琐费时,往往一次所能纯化的蛋白量很有限,为获得所需蛋白用量需多次反复进行。切胶免疫的应用则提供了便利,分离的蛋白不仅纯度高而且凝胶在研磨后可以起到类似于免疫佐剂样的作用。陈鸿军等[14]研究证实用,切胶纯化重组蛋白免疫接种动物能满足制备单克隆抗体的要求。本研究用该法获得高纯度融合蛋白后免疫家兔制备抗血清,间接ELISA 测定多克隆抗体效价,结果显示抗体效价高于1∶12 8000,试验结果再次证明切胶免疫可以制备出高效价的抗血清。
特异性检测在制备多克隆抗体中是必不可少的[15],本研究采取无内毒素重组质粒pVAX1-△CD46转染BHK-21细胞进行目的蛋白真核表达,对表达产物以制备的兔抗His-△CD46融合蛋白多克隆抗体为一抗,采用Western blot检测该多克隆抗体的特异性,结果表明制备的抗牛CD46阳性血清能识别真核表达的目的蛋白,具有较好的特异性。
综上所述,本研究首次扩增到牛CD46基因,在对其进行生物学分析的基础上,成功构建真核和原核表达质粒,并用诱导表达的重组蛋白免疫接种新西兰大白兔制备了抗牛CD46的多克隆抗体,为今后牛CD46生物学功能的研究积累了数据与材料。
[1] Liszewski M K,Atkinson J P.Complement regulator CD46:genetic variants and disease associations[J].Human Genomics,2015,9(1):7.
[2] Cardone J,Le F G,Kemper C,et al.CD46 in innate and adaptive immunity:an update[J].Clin Exp Immunol,2011,164(3):301-311.
[3] Yamamoto H,Fara A F,Dasgupta P,et al.CD46:the ‘multitasker’ of complement proteins[J].Int J Biochem Cell B,2013,45(12):2808-2820.
[4] Price J D,Schaumburg J,Sandin C,et al.Induction of a regulatory phenotype in human CD4+T cells by streptococcal M protein[J].J Immunol,2005,175(2):677-684.
[5] Li K,Zhou W,Hong Y,et al.Synergy between type 1 fimbriae expression and C3 opsonisation increases internalisat ion ofE.coliby human tubular epithelial cells[J].BMC Microbiol,2009(9):64.
[6] Maurer K,Krey T,Moennig V,et al.CD46 is a cellular receptor for bovine viral diarrhea virus[J].J Virol,2004,78(4):1792-1799.
[7] Krey T,Himmelreich A,Heimann M,et al.Function of bovine CD46 as a cellular receptor for bovine viral diarrhea virus is determined by complement control protein 1[J].J Virol,2006,80(8):3912-3922.
[8] Lecot S,Belouzard S,Dubuisson J,et al.Bovine viral diarrhea virus entry is dependent on clathrin-medi- ated endocytosis [J].J Virol,2005,79(16):10826-10829.
[9] Kotani O,Iwatayoshikawa N,Suzuki T,et al.Establishment of a panel of in-house polyclonal antibodies for the diagnosis of enterovirus infections[J].Neuropathology,2014,35(2):107-121.
[10] 张淑莉,李 妮,张 琪.兔抗伤寒杆菌免疫血清在免疫学检验实验教学中的应用[J].检验医学与临床,2015(18):2661-2663.
[11] 钟运华.SmD1蛋白表达与免疫原性研究及其抗体检测初步应用[D].广东广州:广州中医药大学,2014.
[12] Studier F W.Use of bacteriophage T7 lysozyme to improve an inducible T7 expression system [J].J Mol Biol,1991,219(1):37-44.
[13] 宗 超.重组人胰岛素样生长因子-1在大肠杆菌中的高效表达和分离纯化[D].浙江杭州:浙江大学,2013.
[14] 陈鸿军,宋翠萍,秦爱建,等.MDV-1 VP22羧基端在大肠杆菌中高效可溶性表达[J].中国病毒学,2006,21(2):169-172.
[15] 张 迪,张 瑜,陈丹阳,等.OGT多克隆抗体的制备及特异性分析[J].现代生物医学进展,2016(3):414-418.
Abstract:This study aimed to clone and express the CD46 gene from Xinjiang brown cattle and prepare its polyclonal antibody.In this experiment,CD46 gene was amplified from spleen of Xinjiang brown cattle by RT-PCR technique,and after conducting bioinformatics analysis some of this gene fragments were cloned into pVAX1 and pET-28a expression vectors.In order to express target protein,pET28a-△CD46 was transformed intoE.coliRosetta.His-△CD46 expression was induced by IPTG,and then the yield of expressed protein was used to prepare polyclonal antibody in New Zealand white rabbit.Then ELISA method was applied to determine the produced polyclonal antibody titer against bovine CD46.Meanwhile,the recombinant expression plasmid pVAX1-△CD46 was transfected into BHK-21 cells,and then the expressed products were detected via the polyclonal antibody.Results from DNA sequencing of the CD46 full-length gene and size of proteins from CD46 gene fragments was consistent with our expectations.Data from ELISA indicated that titer of the polyclonal antibody was higher than 1∶128 0000,and the antibody also had a good specificity,which was confirmed by Western blot.By this experiment,the His-△CD46 protein was successfully expressed,and a specific polyclonal antibody against bovine CD46 prepared,which provided scientific and material basis for further study on biological function of bovine CD46 gene.
Keywords:Xinjiang brown cattle; CD46; expression; fusion protein; polyclonal antibody
ProkaryoticExpressionofXinjiangBrownCattleCD46GeneandPolyclonalAntibodyPreparationMuhametjan T,
ZHANG Hui-min,DU Run-ci,Jarhin,SU Zhan-qiang,RAN Duo-liang,LIU Jian-hua
(CollegeofVeterinaryMedicine,XinjiangAgriculturalUniversity,Urumqi,Xinjiang,830052,China)
S823.89
A
1007-5038(2017)08-0033-06
2016-12-01
新疆维吾尔自治区自然科学基金项目(2013211A030)
木哈买提江·铁格斯(1988-),男,新疆阿勒泰人,硕士,主要从事分子病毒学研究。*
猜你喜欢
环球时报(2022-09-20)2022-09-20
江西农业学报(2021年4期)2021-04-20
中国生殖健康(2020年2期)2021-01-18
今日农业(2020年24期)2020-12-15
中国医疗保险(2017年5期)2017-05-17
中国康复理论与实践(2015年10期)2015-12-24
小资CHIC!ELEGANCE(2015年15期)2015-09-01
西南医科大学学报(2015年1期)2015-08-22
现代电生理学杂志(2015年1期)2015-07-18
中国当代医药(2015年9期)2015-03-01
