
早期人类演化过程中的关键变量及相互关系
2017-10-11 07:25陈宥成
黄河·黄土·黄种人(华夏文明) 2017年9期
□陈宥成
早期人类演化过程中的关键变量及相互关系
□陈宥成
一、引言
现有分子生物学证据表明,人类和黑猩猩在基因上分离的时间是距今800万年—500万年。这是一个正常的自然选择过程,新生代以来不断有新的哺乳动物出现和老的哺乳动物灭亡。古人类学证据表明,最早人类的显著特征是调整结构后的腿和脚,能够在陆地上两足直立行走。早期人类比现代人类的身材小,和现生大猿一样表现出高度的性二型性,就像两足直立行走的黑猩猩一样[1]。但是,考古学和古人类学证据表明,在随后的人类演化历程中,尤其是在最近300万年左右的时段里,人类自身发生了更多、更显著、更独特的变化。人类制造的石器工具大量出现,脑量成倍增长,性二型性特征明显减弱,并且人类的社会关系向复杂化方向发展。本文试图通过对早期人类的大脑、性二型性等古人类学证据的梳理,结合旧石器考古学的证据,分析人类演化过程中大脑、性二型性与石器技术、语言、社会行为等若干关键变量及其之间的关系。不当之处,敬请方家指正。
二、早期人类大脑的演化
人类与其他动物相比,至少在脑部进化方面具有速度上的绝对优势。在过去的400万年里,我们从一种平凡的、脑量较小的两足直立行走的动物,进化成为不凡的、脑量较大的高级动物。
1.人类大脑演化与语言、打制石器的发展。
人类的大脑是自然界最完美的物质,其精髓在于覆盖在大脑表面的灰质,也就是大脑皮质。鱼类、爬行类和鸟类,大脑皮质由旧皮质和古皮质构成。而哺乳动物大脑皮质以新皮质为主。新皮质的出现是动物神经系统进化过程中具有里程碑意义的事件,因为新皮质具有更高的分析、综合能力。人类具有极为发达的新皮质,其新皮质与其他哺乳动物相比有以下几个优势:一是人类脑容积(相对于体重来说)最大;二是人类大脑皮质沟回最为复杂,大大增加了大脑皮质的表面积;三是人类大脑皮质分区发达和复杂,随着语言的出现,大脑皮质出现了人类特有的语言中枢。
现代人的大脑表面可谓沟壑纵横,其皮质被中央沟、顶枕沟和外侧沟分为额叶、顶叶、枕叶和颞叶不同的部分,不同的分叶上仍发育有复杂的沟回。正电子发射体层摄影(PET)技术,可以通过检测在完成任务过程中局部脑血流量的变化来研究与任务相关的大脑活动。通过此类技术,我们得知大脑皮层不同位置具有不同的功能。其中与人类制造石器有密切关联的部位包括与语言有关的左额下回[2]、与运动有关的中央前回和与空间认知有关的中央后回[3]。长期以来多位学者都提出人类语言、姿势和工具使用的关系。达尔文指出,把一块燧石打成一件古朴的工具,需要双手有灵活性。从这个角度理解,双手的结构可以和发声器官相比[4]。恩格斯说:双手的发展促进了语言的演化[5]。这些假说需要得到神经系统科学的证据。近年来有声语言和手工联系的假说关系得到了一些认知神经科学的证据支持。虽然语言处理长期被认为是和大脑中功能特化的、位置离散的模块有关,现在清楚地表明这些所谓的“语言区域”是和一系列包括工具的使用的非语言行为有关的[6]。大脑中一对一的复杂功能的行为区域图已经被更为抽象的计算性能和情境下的相互作用所代替。
在这种理论框架下需要用灵活的方式把复杂的行为和大脑神经基板联系起来,大脑中每个单一的区域会参与不同的功能[7]。其中比较著名的证据是布洛卡区(Broca’s area),该区域存在语言和行为组织的交叉。布洛卡区过去被认为是左侧第三额下回(the left third inferior frontal convolution)中一个相对独立的负责语言的区域[8]。但是,现在认为语言相关皮质 (frontal languagerelevant cortex)穿过了整个额下回(inferior frontal gyrus,IFG),并且包括理解、语法、语义、语音结构等不同的语言功能。同时,额下回还参与了对象操纵(objectmanipulation)、序列预测(sequence prediction)、视觉搜索、计算、音乐等多种非语言行为。这些表面的多样性行为都来自于额下回的计算功能。并且推测这些功能起源自最初的手工练习(manual praxis)[9]。 语言和使用工具都是有目的的动作。和其他的动作一样,它们的执行和理解都依赖于集成了感觉和动作控制的神经环路(neuaral circuits)。语言和使用工具的不同之处在于,语言以听觉和发声的形式存在,而使用工具显著地和视觉空间、躯体感觉和动手能力有关。无论如何,语言和工具使用有很大的相似性,包括额下回的功能重叠等。
2.人类大脑演化与食肉、狩猎行为的发展。
在过去的400万年里,人类的脑量大概翻了3倍,从最早南方古猿的400毫升,到现代人类的1300~1400毫升。人类化石的证据表明人脑量的大幅度提升发生在距今200万—170万年的早期人属阶段。距今400万—150万年的南方古猿的脑量为430~530毫升;而距今190万—160万年的能人的脑量大于600毫升;距今180万—150万年的直立人的脑量为800~900毫升[10]。虽然直立人的身高也有所增高,但是脑量的增幅明显大于身高增幅。同时,直立人的脑形成等级比今天所有的人类之外的现生灵长类都要高[11]。在南方古猿的进化过程中,南方古猿阿法种的牙齿嚼面的研磨面积为460mm2,后快速增长到南方古猿鲍氏种的756 mm2。相比之下,自距今200万年以来,随着人属的出现,臼齿的嚼面面积明显减少,能人有 478 mm2,而直立人有 377 mm2。直立人与南方古猿相比个头更大,其身高、体重和身体比例更像现代人类[12]。来自化石的证据表明,类人猿的布洛卡区不明显,没有扩大。南方古猿和能人的额叶都具有扩大的布洛卡区。布洛卡区的扩大为语言的形成提供了神经学基础。布洛卡区并不意味着南方古猿和能人能够说话了,只是提示用手制造工具后,脑扩大了,脑的功能复杂化,为语言交流提供了可能性[13]。
人类的大脑与其他哺乳动物的不同之处就是人类的大脑特别大。对这个现象进行解释的一个假说是“昂贵组织假说”(expensivetissue hypothesis),其认为人类在进化过程中在人脑尺寸和消化道尺寸之间有一个权衡,因为人类的消化道比其他灵长类的消化道尺寸要小。人类与其他灵长类的显著区别(比如直立行走、大的脑量)与人类独特的营养需求有关[14]。大脑和其他神经组织对能量的需求(kcal/g/min)是很高的,大约是骨骼肌组织的16倍[15]。但是,人类身体的其他组织对能量的需求与同样尺寸的其他哺乳动物相比是低的。人体大脑的RMR(restingmetabolic rate)占人体的20%~25%,其他灵长类只占到8%~10%[16]。人体大脑新陈代谢独特的能量预算可以暗示出人类独特的饮食需求。
古人类学证据表明人类大脑在180万年前直立人阶段开始快速增长,并且伴随着饮食、身高和行为的变化。大的脑量消耗更多的能量,人类大脑的新陈代谢比其他灵长类要消耗更多的能量,同时也需要高能量、高营养的饮食来维持运转。直立人的骨骼结构表明其食物中植物类食物有所减少,而高热量食物有所增加。直立人体质变化与环境的变化有明显关联。随着气候变干,森林面积减少,草原面积增大,在非洲,严重影响了人类食物资源的分布,比如动物性食物的丰富程度增加,体质也会随之发生变化。考古证据表明,直立人具有比较初步的合作狩猎行为,其通过狩猎和对抗性的活动获得肉食[17]。同时,直立人会把宰杀后的猎物运回中心营地,在群体内部分享[18]。人类食肉量的增加,不但提供了更多的能量,而且提高了人体内非常关键的脂肪酸的含量,其为人脑的进化提供了重要基础。
三、早期人类的性二型性特征的演化
自从达尔文的《人类的由来》(The desent of Man)和《性别的选择》(Selection in relation to sex)发表以来,学术界对于动物界尤其是灵长类和人类的性二型性有着广泛的讨论,无论是现生的还是灭绝的物种。其中讨论的重点是如何去解释早期人类及其祖先的性二型性,及关于对人类的交配系统和社会结构的推断。很多研究都通过犬齿大小和体重来估算南方古猿的性二型性,并推测其交配系统。但是,推测结果包括所有不同的交配方式,从一夫一妻到一夫多妻[19]。
对于大多数类人猿来说,雄性都比雌性的质量要大。当今人类,男性比女性的质量大大约15%;而现生大猩猩、猩猩等则大于50%。要得到现生物种的单体质量是比较容易的,但是对于化石而言,尤其是残缺的化石,其单体质量是比较难得到的。McHenry曾通过肱骨、股骨、胫骨的长度计算身高,进而计算距今400万—140万年的95个人类化石(包括我们熟知的Hadar地区的大量化石标本、Sterkfontein的大量化石标本、Omo遗址的化石标本、Olduvai的化石标本、Swartkrans的化石标本、Kromdraai的化石标本等)样本的身高、体重,结果显示:南方古猿阿法种男性和女性的体重是45公斤和29公斤;南方古猿非洲种男性和女性的体重是41公斤和30公斤;南方古猿粗壮种男性和女性的体重是40公斤和32公斤;南方古猿鲍氏种男性和女性的体重是49公斤和34公斤;能人男性和女性的体重是52公斤和32公斤[20]。McHenry随后指出南方古猿阿法种的男性比女性大50%,南方古猿非洲种男性比女性大40%,直立人的男性比女性大20%。表明人类两性之间的能量消耗在逐渐平衡。
后来发现学者们对于南方古猿的性二型性认识也有误解,原因包括:(1)选取估算质量的标本太小;(2)性二型性估算是建立在性别鉴定准确的基础上的,但事实上,第二性征通常表现在盆骨上,而化石中盆骨经常缺失;研究者只能认为雄性骨骼尺寸大于雌性骨骼,其准确性大大降低;(3)性二型性的确定需要建立在种属鉴定正确的基础上,但种属鉴定往往有争论,所以拿不同种的化石来比较性二型性是有问题的;(4)性二型性的变化是非常缓慢的,其尺度是几十万年或几万年;(5)如果拿不同地区的个体做比较,会放大性二型性的尺度。最近,有科学家通过化石股骨头的大小来估算人科物种的质量。Reno等通过统计建模绕开了残存化石的上述问题,采用了新的方法去计算南方古猿阿法种的性二型性,其选用的数据来源是埃塞俄比亚Hadar地区的A.L.333的大概320万年前因灾难性事件同时死亡的南方古猿阿法种化石标本。选用A.L.288的保存有40%的Lucy作为形态测定的模板,因为其保存有较为完整的股骨头和其他长骨 (Lucy是南方古猿阿法种最完整的化石,是形态测定的模板,标本现存于埃塞俄比亚自然博物馆);然后统计了其他阿法种标本的股骨头直径,分析的结果表明南方古猿阿法种有中等程度的性二型性,比大猩猩等更像人类和黑猩猩。综合不同学者的意见整体来看,南方古猿阿法种的犬齿性二型性低于黑猩猩[21],直立人的性二型性明显要低于南方古猿阿法种,与现代人类差别不大。
四、早期人类合作行为的演化
人类的合作起源于灵长类,包括分配、外婆喂养、食物分享等。灵长类的群居生活使每个个体都有收益,并且付出的代价也非常明显。直接的代价来自群体内部对食物、配偶和其他资源的竞争。间接地付出包括群体内部成员对于团结的协调与配合。为了平衡竞争,需要很高程度的合作。在许多灵长类种群中,个体在种群中的等级位置和繁殖的成功依赖于与群体内其他成员的合作和协调能力[22]。灵长类中除了人类之外,黑猩猩具有最广泛的合作行为。雄性黑猩猩形成了长期的稳定联盟,一起出行,为彼此打扮,还有合作狩猎。占统治地位的雄性依靠联盟去保持自己的等级,并通过允许接触雌性奖励它们的支持者。在黑猩猩和倭黑猩猩的群体内,每个成员都清楚地知道自己联盟的其他成员,群外和一个群体中的某个个体的冲突可能会导致和整个群体的冲突。
但是,灵长类研究表明,性二型性程度高的黑猩猩、大猩猩等物种,雄性将大量的能量消耗在寻找伴侣的竞争之中,但孩子出生之后付出甚少,导致孩子不知其父。关于性二型性与竞争的关联分析是:当性二型性高的时候,雄性之间的竞争比较普遍;当性二型性低的时候,雄性之间的竞争较为不普遍[23]。在繁殖后代的过程中,雌性灵长类承担的责任包括怀孕、分娩和抚养孩子等,需要消耗大量的能量;对于雄性灵长类来说,包括寻找、吸引和守护伴侣。无论是雄性还是雌性,能量的消耗都是决定行为的重要因素。具体来讲,繁殖过程中能量的消耗会影响什么时候合作、与谁合作等的决定[24]。
人类身体的大小与大脑的大小、喂食行为、栖居行为、社会行为等都有联系[25]。通过对现生灵长类和人类的研究,发现雄性和雌性之间的协作发生在雌性在繁殖过程中的能量消耗高于雄性时,这时父亲的抚养就会出现。人类在演化过程中做出了新的选择,人类中的男性在繁育过程中,将寻找伴侣、竞争的能量减少,同时将更多的能量弥补于孩子出生后男女之间的长期合作。人类在演化过程中性二型性逐渐降低,尤其是到直立人阶段。这表明人类在进化的过程中女性的能量消耗逐渐增大,这时需要两性之间更多的合作,比如父亲的抚养,男性耗费更多的能量用于为女性和孩子提供食物,用于教育孩子,以保证孩子的成活,使自己的基因得以延续。
人类的合作不仅存在于母亲和孩子、丈夫和妻子之间,同时,人类的协作包括更广阔的个体网络,有更多样的行为。人类的社会网络包括非家族的、家族的,以及不同年龄和性别的[26]。考古学证据表明,50万—40万年前,合作狩猎大型动物的行为出现在德国Schoeningen和英国Boxgrove等遗址中。在这一阶段,男性之间的合作明显增强,尤其是在为女性和孩子提供食物方面。无论是古人类学的性二型性证据,还是考古学的证据,都暗示人类在直立人阶段已经形成了复杂的社会合作关系。
五、讨论与结语
在距今大约300万年的时候,地球上发生了很多特殊的事件,包括一些特殊的灵长类开始大规模制造和使用工具,这些特殊的灵长类的脑量急速增大,两性之间的差异明显缩小,并且这些特殊的灵长类的规模日益增大,形成了复杂的社会网络,并凭借其高智力而非身躯成为地球的统治者。这些事件在地球上是史无前例的。毫无疑问,这些特殊的灵长类就是人类。人类从一种默默无闻的动物逐渐成为地球上的不凡的强大的动物。
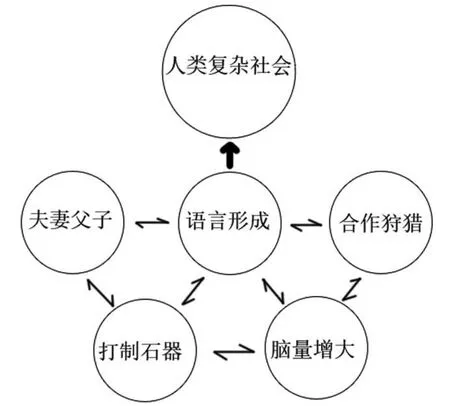
那么,强大的人类是怎样变成的?这其中涉及前文提到的人类演化过程中的一系列关键变量,包括脑量、语言、夫妻关系、父子关系打制石器及合作狩猎(见下图)。其中语言的出现是非常重要的。南方古猿和能人扩大的布洛卡区暗示人类最初的简单语言可能出现,而直立人阶段翻倍的脑量、复杂的石器技术、广泛的合作狩猎行为,显示人类已经拥有了早期语言。如果没有语言,人类在合作狩猎的过程中难以快速分享情报和快速做出判断,进而成功狩猎,获取肉食,为人类大脑的演化提供能量基础。如果没有语言,人类一生积累的经验、知识和技能难以代代传递,石器技术的稳定延续就难以保障,不能步步进化和复杂化。
同时,这些变量之间相互构成了一个“超循环”体系,相互促进,相互影响。如脑量的增大可以进一步为语言的形成提供神经基础,而大脑中负责语言和与打制石器相关的计算属于同一区域,它们之间可以相互促进,人类社会不断复杂化需要建立更广泛的合作关系以维持其成功运转,对语言的需求也不断增加。这些变量之间相互作用,就像滚雪球似的,越滚越大,推动了人类复杂社会的形成,进而奠定了人类的文化性格与社会性格。〔本文为“国家哲学社会科学基金青年项目 (项目编号:16CKG004)”和“北京市教委科技创新平台项目——首都手工业遗产的文化传承与利用创新平台”的阶段性研究成果〕。
人类演化过程中的关键变量及其关系
[1]R ICHARD G.KLEIN:Archaeology and the evolution of human behavior[J].Evolutionary Anthropology,2000,(9).
[2]BROCA P.:Comments regarding the seat of the faculty ofspoken language,followed by an observation of aphem ia (loss of speech),In Broca’s region(eds Y.GRODZINSKY&K.AMUNTS)[M].Oxford,UK:Oxford University Press,2006.
[3]DIETRICH STOUT,N ICHOLAS TOTH &KATHY SCHICK:Stone tool-making and brain activation:position em ission tomography (PET)studies[J].Journalofarchaeologicalscience,2000,(12).
[4]DARW IN C.:The descent of man,and selection in relation to sex[M].London,UK:Penguin Books,2004.
[5]EEGELS F.:The part played by labor in the transition from ape to man,In Philosophy of technology[M].London,UK:Blackwell,2003.
[6]BOOKHEIMER S.:Functional MRI of language:new approaches to understanding the cortical organization of semantic processing[J].Annual Review of Neuroscience,2002,(25).
[7]DEACON T.W.:The symbolic species:the coevolution of language and the brain[M].New York:W.W.Norton,1997.
[8]BROCA P.:Comments regarding the seat of the faculty of spoken language, followed by an observation of aphemia (loss of speech),In Broca’s region(eds Y.GRODZINSKY&K.AMUNTS)[M].Oxford,UK:Oxford University Press,2006.
[9]PULVERMUELLER F.& FADIGA L.:Active perception:sensorimotor circuitsasa corticalbasis for language[J].Nat.rev.Neurosci.2010,(11).
[10]W illiam R.Leonard,J.Josh Snodgrass,and Marcia L.Robertson:Effects of brain evolution on human nutrition and metabolism[J].The Annual review of Nutrition,2007,(27).
[11]LEONARDW.R.&ROBERTSONM.L.:Evolutionary perspectives on human nutrition:the influence of brain and body size on diet and metabolism[J].American JournalofHuman Biology,1994,(6).
[12]MCHENRY H.M.:Body size and proportions in early hom inids [J].American Journal of physical Anthropology,1992,(87).
根据图纸说明并结合相关标准规范要求,在建筑模型中创建构造柱、圈梁、门过梁、反坎和门槛等二次结构构件,并生成材料明细表精确提取单层材料运输量,对构件进行平面标识,输出二维和三维深化图,指导现场施工(见图 5)。
[13]吴秀杰:《化石人类脑演化研究概况》,《人类学学报》2003年第22卷第3期。
[14]AIELLO L.C.&WHEELER P.:The expensivetissue hypothesis:the brain and the digestive system in human and primate evolution [J].Current Anthropology,1995,(36).
[15]HOLLIDAYM.A.:Body composition and energy needs during grow th,In Human Grow th:A comprehensive Treatise [M].Volume 2,ed.F.FALKNAR,J.M.TANNER.New York:Plenum,1986.
[16]LEONARDW.R.&ROBERTSONM.L.:Evolutionary perspectives on human nutrition:the influence of brain and body size on diet and metabolism[J].American Journalof Human Biology,1994,(6).
[17]PLUUMER T.:Flaked stones and old bones:biological and cultural evolution at the dawn of technology[J].Physical Anthropology,2004,(47).
[18]POTTS R.:Early Hom inids activities at O lduvai[M].New York:Aldine,1988.
[19]PLAVCAN J.M.&VAN SCHAIK C.P.:Interpreting hom inid behavior on the basis of sexual dimorphism[J].Journalof Human Evolution,1997,(32).
[20]HENRY M.MCHENRY.:Body size and proportions in early Hom inids[J].American Journal of Physical Anthropology,1992,(87).
[21]CLARK S.LARSEN.:Equality for the sexes in human evolution?Early hom inid sexual dimorphism and implications for mating systems and social behavior[J].PNAS,2003,(100).
[22][25]ROBIN D.U.ChirsKnightand Cam illaPower:The evolutionofculture[M].RutgersUniversityPress,2003.
[23]PLAVCAN J.M.&CARREL P.VAN SCHAIK.:Interpreting hom inid behavior on the basis of sexual dimorphism[J].JournalofHuman Evolution,1997,(32).
[24]HENRY M.MCHENRY.:How big were early Hom inids[J].Evolutionary Anthropology,1992,(1).
[26]WHALLON R.:Elements of cultural change in the later Paleolithic,(edsMELLARSP.&STINGER C.),The Human Revolution[M].Edinburgh:Edinburgh University Press,1989.
(作者单位 首都师范大学历史学院)
[责任编辑 赵建春 尚元昕]
猜你喜欢
海外星云(2021年9期)2021-10-14
考试与评价·高二版(2020年6期)2020-09-10
小猕猴智力画刊(2020年2期)2020-03-08
儿童故事画报·发现号趣味百科(2019年12期)2019-02-02
森林与人类(2016年7期)2016-08-11
小天使·三年级语数英综合(2016年9期)2016-05-14
小学科学(2015年2期)2015-03-11
小学科学(2015年1期)2015-03-11
科学大众·小诺贝尔(2009年9期)2009-10-23
小学生导刊(中年级)(2007年6期)2007-06-08
