
高盐胁迫对凡纳滨对虾生长指标、血浆渗透压及Na+-K+-ATP酶活力的影响
2017-09-19 15:52王仁杰赵玉超苏文辉李玉全
浙江海洋大学学报(自然科学版) 2017年3期
李 娜,王仁杰,赵玉超,沈 敏,苏文辉,赵 超,李玉全
(1.青岛农业大学海洋科学与工程学院,山东青岛 266109;2.东营市海洋与渔业局,山东东营 257091;3.莱西市水产局,山东青岛 266600)
高盐胁迫对凡纳滨对虾生长指标、血浆渗透压及Na+-K+-ATP酶活力的影响
李 娜1,王仁杰1,赵玉超1,沈 敏1,苏文辉2,赵 超3,李玉全1
(1.青岛农业大学海洋科学与工程学院,山东青岛 266109;2.东营市海洋与渔业局,山东东营 257091;3.莱西市水产局,山东青岛 266600)
为探讨高盐对凡纳滨对虾的生长指标、血浆渗透压及Na+-K+-ATP酶活力的影响,设置30、40、50、60共4个盐度梯度,检测存活率、相对增重率、特定生长率、摄食率、饵料转化率、血浆渗透压、Na+-K+-ATP酶活力等指标。结果表明,随着盐度增加,凡纳滨对虾的存活率、相对增重率、特定生长率、饵料转化率等指标逐渐降低,摄食率、血浆渗透压及Na+-K+-ATP酶活力逐渐升高。说明高盐显著影响凡纳滨对虾的渗透压及Na+-K+-ATP酶活力,且高盐不利于凡纳滨对虾的生长及饵料利用。
凡纳滨对虾;高盐;生长指标;血浆渗透压;Na+-K+-ATP酶
凡纳滨对虾Litopenaeus vannamei,俗称南美白对虾,原产于秘鲁北部至墨西哥桑诺拉一带的太平洋沿岸。具广盐性,生长快,环境适应能力强等特点,于1988年首次引进我国,近几年随着对虾养殖的发展,其养殖规模和面积不断扩大,是全球养殖产量最高的对虾品种[1]。据报道,凡纳滨对虾可耐受的盐度变化为2~78[2]。因其具有耐广盐的特性,国内在淡水、半咸水、高盐水中都进行了大规模养殖。对虾在较低盐度环境条件下生长较快,高盐度下生长减慢[3],但高盐虾在口感、品质等方面明显优于低盐虾。我国沿海或西北地区还存在大量的高盐水体,如山东盐场盐度30~70的水域面积约13×104hm2。这些水域可以发展凡纳滨对虾养殖,并且也有不少高盐水体已经开展了相关的养殖生产,但单位面积产量一般不高于400 kg/hm2,远低于其他盐度水体。关于盐度对虾类的影响前人已做过大量报道,涉及生长、存活、饵料利用、酶活、分子等多个方面,但研究主要集中在低于正常海水盐度的水体[4-7],然而目前,关于高盐度对对虾生长、代谢、理化等的影响国内相关文献报道甚少。本实验以凡纳滨对虾为实验材料,通过分析高盐对对虾存活率、相对增重率、特定生长率、饵料转化率、血浆渗透压、Na+-K+-ATP酶活力的影响,以期丰富对虾逆境生物学理论,并为高盐对虾养殖提供理论借鉴。
1 材料与方法
1.1 实验对象的来源、暂养与驯化
实验在青岛农业大学海洋科学与工程学院开放实验室进行。实验所用凡纳滨对虾购自昌邑市海丰水产养殖有限责任公司。实验用虾运至青岛农业大学大学实验室后,暂养于水族箱中。暂养期间所用海水为天然海水,盐度30±2,pH值7.9±0.5,用加热棒将温度控制在27±0.5℃,24 h充气泵充氧,每日换水2次,每次换水30%。暂养7 d后,筛选相近规格的凡纳滨对虾,以每天5个盐度的速度由30逐渐驯化至实验设计梯度。高盐度海水由天然海水(盐度30左右)与粗盐调配而成。
1.2 试验设置与数据测定
实验于35 cm×25 cm×20 cm PVC箱中进行,设置30、40、50、60共4个盐度梯度,每个处理设3个平行组,每个处理放入调整到设计盐度的凡纳滨对虾20尾。试验前停食1 d,以排空肠胃中的粪便。各试验组连续充气。每天投喂2次(7:30、19:30),日投喂量为对虾湿重的8%,每天换水2次,每次换水30%;每天于9:30和21:30收集残饵2次,70℃烘干称重后保存;每隔10 d测定1次体长和体重,计算特定生长率、存活率、相对增重率、摄食率等指标。
分别称取10 g饵料和随机挑选20尾凡纳滨对虾,称重后在70℃条件下烘干至恒重,估算试验开始时饵料和对虾的干重(g)、水分含量(g),重复3次。
在相同的水环境下,分别称取10 g饵料,记作m0,放入不同盐度处理中,浸泡2.0 h,收集残饵于70℃条件下烘干至恒重,记作m1,计算饵料在不同盐度海水中的溶失率。
实验结束后每个处理随机取6尾凡纳滨对虾,用l.0 mL一次性注射器从对虾心脏抽取血淋巴,收集于1.5 mL的Eppendorf管中,4℃过夜后4 000 r/min离心15 min,收集血清测量其渗透压。取其鳃组织放入离心管中,迅速投入液氮冷冻,再转移到-80℃冰箱保存,以上操作均在冰盘中进行。组织样品先用剪刀剪碎,准确称量后置于玻璃匀浆管中,加入9倍体积的生理盐水,匀浆机匀浆,后冷冻离心机3 000 r/min离心10 min,分别稀释至所需浓度后进行Na+-K+-ATP酶活力的测定。Na+-K+-ATP酶活力及蛋白含量(蛋白的测定采用考马斯亮蓝法)的测定均采用南京建成生物研究所研制的检测试剂盒,按照操作步骤进行操作。
1.3 数据计算
饵料溶失率=(m0-m1)/m0× 100%
存活率=(存活数/开始时的总数)×100%
相对增重率 WG=100(Wt-W0)/W0
特定生长率SGR=100(LnW2-LnW1)/T
摄食率FId=100F/[T*(W2+W1)/2]
食物转化率FCR=100(W2-W1)/F
式中:W0(W1)和Wt(W2)分别为实验起始和结束时对虾湿(干)重(g),T为实验天数(d),F为实验过程中对虾所摄取的饵料量(换算为干重,g)。
1.4 数据处理
所得数据采用SPSS13.0软件进行方差分析(ANOVA)及Duncan多重比较,以P<0.05作为差异显著。
2 结果与分析
2.1 高盐对凡纳滨对虾生长指标的影响
2.1.1 饵料溶失率
从表1可以看出,随着盐度的升高,饵料溶失率逐渐降低,盐度30~50处理间差异不显著(P>0.05),盐度50和60处理间差异显著(P<0.05)。在盐度30时,饵料溶失率最高,达到25.86%;盐度60时,溶失率最低,为21.85%。

表1 高盐对饵料溶失率的影响Tab.1 Effect of high salinity on dissolve-loss ratio of feed
2.1.2 存活率
从表2可以看出,随着盐度的升高,凡纳滨对虾的存活率逐渐降低,且各处理组间差异显著(P<0.05)。盐度30时存活率最高,为88.33%,盐度60时存活率最低,为38.33%。

表2 高盐对凡纳滨对虾存活率的影响Tab.2 Effect of high salinity on survival rate of L.vannamei
2.1.3 相对增重率
从表3可以看出,随着盐度的升高,凡纳滨对虾的相对增重率逐渐降低,且各处理组间差异显著(P<0.05)。盐度30时相对增重率最高,为549.57%,盐度60时相对增重率最低,为203.07%。
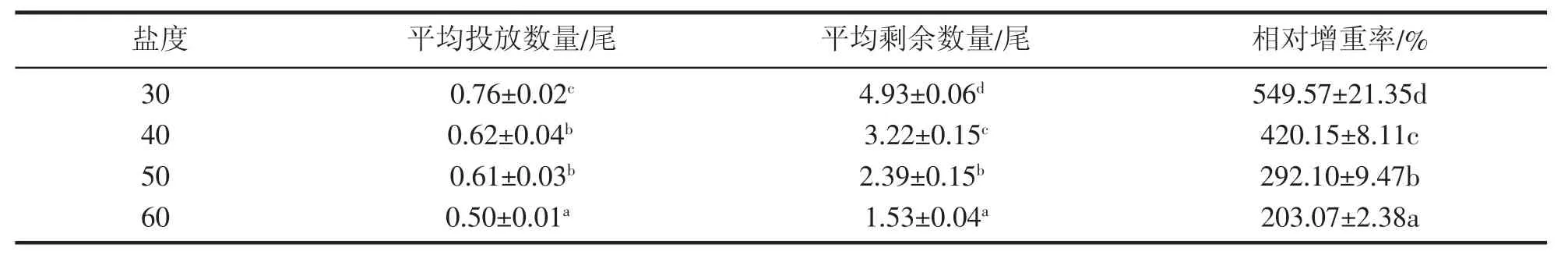
表3 高盐对凡纳滨对虾相对增重率的影响Tab.3 Effect of high salinity on the relative weight gain rate of L.vannamei
2.1.4 特定生长率
从表4可以看出,随着盐度的升高,凡纳滨对虾的特定生长率逐渐降低,且各处理组间差异显著(P<0.05)。盐度30时特定生长率最高,为56.24%,盐度60时特定生长率最低,为3.70%。
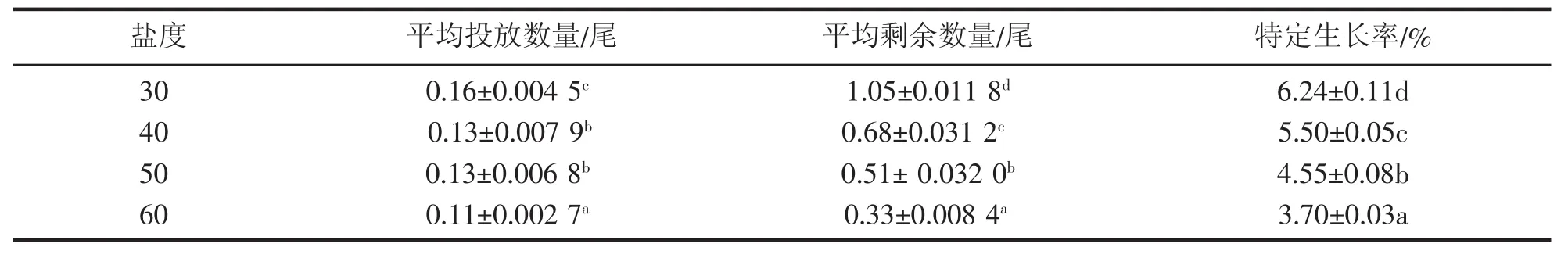
表4 高盐对凡纳滨对虾特定生长率的影响Tab.4 Effect of high salinity on specific growth rate of L.vannamei
2.1.5 摄食率
从表5可以看出,随着盐度的升高,凡纳滨对虾的摄食率逐渐升高,且各处理组间差异显著(P<0.05)。盐度30时摄食率最低,为6.07%,盐度60时摄食率最高,为9.04%。
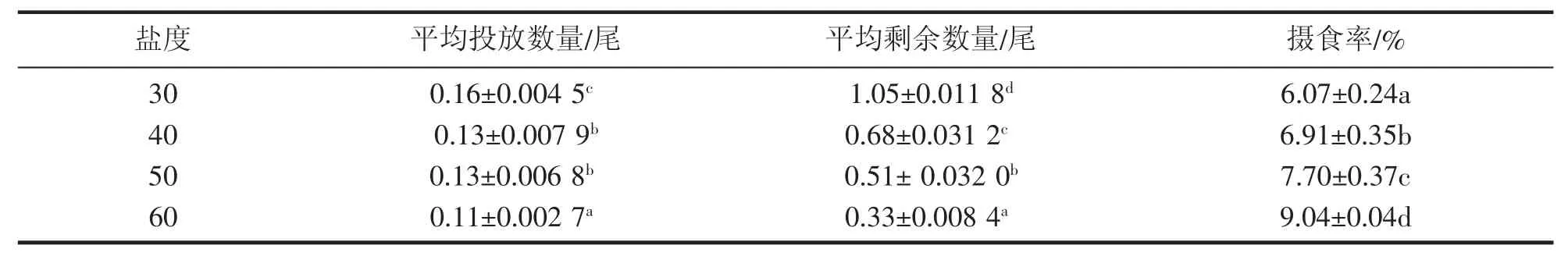
表5 高盐对凡纳滨对虾摄食率的影响Tab.5 Effect of high salinity on feeding rate of L.vannamei
2.1.6 饵料转化率
从表6可以看出,随着盐度的升高,凡纳滨对虾的饵料转化率逐渐降低,且各处理组间差异显著(P<0.05)。盐度30时饵料转化率最高,为80.61%,盐度60时饵料转化率最低,为37.16%。

表6 高盐对凡纳滨对虾饵料转化率的影响Tab.6 Effect of high salinity on FCR of L.vannamei
2.2 高盐对凡纳滨对虾血浆渗透压的影响
从图1可以看出,随着盐度的升高,血浆渗透压呈显著上升的趋势。血浆渗透压表现为盐度为50时最高(P<0.05),盐度 30时最低(P<0.05),盐度 50和 60处理组间差异不显著(P>0.05),盐度 30、40和 50处理组间差异显著(P<0.05)。
2.3 高盐对凡纳滨对虾Na+-K+-ATP酶活力的影响
从图2可以看出,随着盐度的升高,Na+-K+-ATP酶活力呈显著上升的趋势。Na+-K+-ATP酶活力表现为盐度为60时最高(P<0.05),盐度30时最低(P<0.05),盐度50和60处理组间、盐度40和50处理组间、盐度50和60处理组间差异不显著(P>0.05),盐度30和50、60处理组间、盐度40和60处理组间差异显著(P<0.05)。

图1 高盐对凡纳滨对虾血浆渗透压的影响Fig.1 Effect of high salinity on plasma osmotic pressure of L.vannamei

图2 高盐对凡纳滨对虾Na+-K+-ATP酶活力的影响Fig.2 Effect of high salinity on Na+-K+-ATPase activities in L.vannamei
3 讨论
3.1 高盐对凡纳滨对虾生长指标的影响
盐度是一种渗透调节因子,在不适的盐度环境中水生生物会产生应激反应来维持体内外的渗透平衡,因此就要消耗额外的能量,从而影响其生长、存活等[4]。申玉春等[6]认为,凡纳滨对虾最适生长盐度在22左右,盐度为30时,饵料转化率最高。黄凯等[8]研究发现,盐度能显著影响凡纳滨对虾成活率、体质量增长率和饲料系数(P<0.05),在盐度20时生长最好。王兴强等[9]研究发现,凡纳滨对虾特定生长率在盐度20时最高,存活率在盐度35时最高。朱春华等[10]研究发现,凡纳滨对虾的等渗点在14~22盐度附近,此盐度范围内用于渗透压调节的耗能较少,而盐度低于14或高于22时,身体内外由于存在较大的渗透压梯度差,为维持自身渗透平衡消耗了更多的能量,在一定程度上影响了其生长。PANIKKAR[11]研究发现,广盐性虾类可通过离子调节和血淋巴的渗透调节来适应外界环境盐度的变化,具有二重性。高盐度条件下,虾类需将体内多余的盐分排出体外,保持体内的正常水分;较低盐度条件下,又需要摄取足够的盐分,排掉多余的水分。在渗透压主动调节过程中,虾类要消耗体内储存的能量,因而代谢率升高,降低了生长效能。对虾生活的盐度在等渗点附近时新陈代谢能耗降低,能够使生长加快[12]。林黑着等[13]对鱼类的研究发现,盐度升高,代谢消耗增大,消化速度也随之加快,消化率显著提高。本研究发现,凡纳滨对虾在不同盐度下的存活率、相对增重率、特定生长率、饵料转化率均表现为 30>40>50>60,摄食率表现为 60>50>40>30。说明,高盐下凡纳滨对虾虽然摄取了较多的饵料,但由于用于调节渗透的能量增加,生长受到抑制。因此,从本实验结果来看,凡纳滨对虾在盐度30-40范围内生长性能最好,这与前人在凡纳滨对虾上的报道一致[4-12]。
3.2 高盐对凡纳滨对虾渗透压和Na+-K+-ATP酶活力的影响
有研究指出盐度是甲壳类赖以生长与存活的重要环境因子,它能通过调节渗透压从而影响甲壳类的生理活动,而Na+-K+-ATPase对外界环境因子的变化也十分敏感,该酶的活性是反映机体生长状况、渗透调节和生理代谢的重要指标[14-17]。前人为了研究盐度变化对甲壳动物体内生理生化指标的影响,对甲壳动物在不同盐度环境下血浆渗透压和鳃组织中的Na+-K+-ATPase活性进行了大量研究[18-21]。LIU等[22]研究发现,随着盐度的升高,中华绒螯蟹血浆渗透压也逐渐升高。BROOKS等[23]研究发现,广盐性海洋甲壳动物盐度变化后,鳃上皮细胞Na+-K+-ATPase的活性增大,其活性与体内外的渗透压差呈正相关。FURRIEL[24]研究发现,罗氏沼虾在提高盐度后鳃中的酶比活力增加。本实验中,随着盐度的升高,凡纳滨对虾血浆渗透压和Na+-K+-ATP活力也逐渐升高,这与前人的报道一致[22-24]。分析认为,外界环境盐度增加导致体内渗透压增高和细胞的内外极性增加,由于顶部质膜的V-ATPase为Cl-的吸收提供了动力,导致鳃上皮细胞中Cl-浓度增加,为了最终实现渗透平衡,进而导致Na+-K+-ATPase活力增高,从而促进Na+通过基底侧质膜进入血淋巴。有研究认为,基因表达量由于渗透压的长期调节刺激而发生了变化,进而对Na+-K+-ATPase位点数量进行了调节,从而影响了酶的活力,使酶活力升高[25]。高盐的分子调控机理还需进一步研究。
[1]华建权,郑忠明,陈辉煌,等.凡纳滨对虾池塘不同混养模式技术的研究 [J].浙江海洋学院学报:自然科学版,2014,33(5):455-482.
[2]张伟权.世界重要养殖品种—南美白对虾生物学简介[J].海洋科学,1990(3):69-72.
[3]BRAY W A,LAWRENCE A L,LEUNG-TURJILLO J R.The effect of salinity on growth and survival of Penaeus vannamei,with observations on the interaction of HHN virus and salinity[J].Aquaculture,1994,122(2):133-146.
[4]李玉全.盐度对脊尾白虾生长、摄食及饵料转化率的影响[J].江苏农业科学,2014,42(9):191-193.
[5]戴习林,张立田,臧维玲,等.Ca2+、Mg2+、盐度对凡纳滨对虾存活、生长及风味的影响[J].水产学报,2012,36(6):914-921.
[6]申玉春,陈作洲,刘 丽,等.盐度和营养对凡纳滨对虾蜕壳和生长的影响[J].水产学报,2012,36(2):290-299.
[7]侯文杰,臧维玲,刘永士,等.盐度及Ca2+与Mg2+含量对凡纳滨对虾生长及虾体钙镁含量的影响[J].江苏农业科学,2012,40(3):193-196.
[8]黄 凯,王 武,卢 洁,等.盐度对南美白对虾的生长及生化成分的影响[J].海洋科学,2004,28(9):20-25.
[9]王兴强,曹 梅,马 甡,等.盐度对南美白对虾的生长及生化成分的影响[J].海洋水产研究,2006,27(1):8-13.
[10]朱春华.盐度对南美白对虾生长性能的影响[J].水产养殖,2002(3):25-27.
[11]PANIKKAR N K.Osmotic behavior of shrimps and prawn sin relation to their biology and culture[R].FAO Fish Rep,BCSP/67/E/25,1967:527-538.
[12]RUSCOE I M,SHELLEY C C,WILLIAMS G R.The combined effects of temperature and salinity on growth and survival of juvenile mud crabs(Scylla serrata Forskal)[J].Aquaculture,2004,238(1/4):239-247.
[13]林黑着,江 琦,石 红,等.盐度对鲻鱼表观消化率的影响[J].浙江海洋学院学报:自然科学版,2001,20:80-82.
[14]王庚申,谢建军,施 慧,等.不同盐度对脊尾白虾非特异性免疫及抗氧化酶活性的影响[J].浙江海洋学院学报:自然科学版,2013,32(6):499-502.
[15]潘鲁青,刘泓宇.甲壳动物渗透调节生理学研究进展[J].水产学报,2005,29(1):109-115.
[16]SATYAVATHI C,RAO Y P.Inhibition of Na+-K+-ATPase in Penaeus indicus postlarvae by lead[J].Comparative Biochemistry and Physiology C:Pharmacology,Toxicology and Endocrinology,2000,127(1):11-22.
[17]SANCHO E,FERNA N V,FERRANDO M D,et al.ATPase activity as biomarker of thioben carb exposure[J].Ecotoxicology and Environmental Safety,2003,56(3):434-441.
[18]HURTADO M A,RACOTTA I S,CIVERA R,et al.Effect of hypo-and hypersaline conditions on osmolality and Na+/K+-ATPase activity in juvenile shrimp(Litopenaeus vannamei)fed low and high-HUFA diets[J].Comparative Biochemistry and Physiology A:Molecular&Integrative Physiology,2007,147(3):703-710.
[19]PAN Lu-qing,LUAN Zhi-hua,JIN Cai-xia.Effects of Na+/K+and Mg2+/Ca2+ratios in saline groundwaters on Na+-K+-ATPase activity,survival and growth of Marsupenaeus japonicus post larvae[J].Aquaculture,2006,261(4):1396-1402.
[20]LUCU C姚,DEVESCOVI M.Osmoregulation and branchial Na+,K+-ATPase in the lobster Homarus gammarus acclimated to dilute seawater[J].Journal of Experimental Marine Biology and Ecology,1999,234(2):291-304.
[21]LUCU C姚,TOWLE D W.Na++K+-ATPase in gills of aquatic crustacean[J].Comparative Biochemistry and Physiology A:Molecular&Integrative Physiology,2003,135(2):195-214.
[22]LIU Hongyu,PAN Luqing,LYU Fu.Effect of salinity on hemolymph osmotic pressure,sodium concentration and Na+-K+-ATPase activity of gill of Chinese crab,Eriocheir sinensis[J].Journal of Ocean University of China,2008,7(1):77-82.
[23]Brooks S J,Mills C L.Gill Na+,K+-ATPase in a series of hyper-regulating gammarid amphipods.Enzyme characterisation and the effects of salinity acclimation[J].Comparative Biochemistry and Physiology A:Molecular&Integrative Physiology,2006,144(1):24-32.
[24]WILDER M N,HUONG D T T,ATMOMARSONO M.Characterization of Na/K-ATPase in Macrobrachium rosenbergii and the effects of changing salinity on enzymatic activity[J].Comparative Biochemistry and Physiology A:Molecular&Integrative Physiology,2000,125(3):377-388.
[25]MENDONCA N N,MASUI D C,McNAMARA J C,et al.Long-term exposure of the freshwater shrimp Macrobrachium olfersii to elevated salinity:Effects on gill(Na+,K+)ATPase α-subunit expression and K+-phosphatase activity[J].Comparative Biochemistry and Physiology,2007,146(4):534-543.
Effects of High Salinity on Growth Index,Plasma Osmotic Pressure and Na+-K+-ATPase Activities of Litopenaeus vannamei
LI Na,WANG Ren-jie,ZHAO Yu-chao,et al
(College of Marine Science and Technology,Qingdao Agricultural University,Qingdao 266109,China)
This study was conducted to investigate the effects of high salinity on growth index,plasma osmotic pressure and Na+-K+-ATPase activities of Litopenaeus vannamei.Four salinity levels(30,40,50 and 60)were designed.The survival rate,relative weight gain rate,specific growth rate,feeding rate,feed conversion rate,plasma osmotic pressure and Na+-K+-ATPase activities were analyzed.The results showed that survival rate,relative weight gain rate,specific growth rate and food conversion rate(FCR)decreased,feeding rate,plasma osmotic pressure and Na+-K+-ATPase activities increased with salinity.Therefore,significant different were tested on plasma osmotic pressure and the activity of Na+-K+-ATPase,high salinity is not conducive to the growth and feed utilization of Litopenaeus vannamei.
Litopenaeus vannamei;high salinity;growth index;plasma osmotic pressure;Na+-K+-ATPase
S917.4
A
1008-830X(2017)03-0196-06
2017-02-19
国家自然科学基金项目(31101916);山东省现代农业产业技术体系虾蟹类创新团队(SDAIT-15-011);青岛市科技成果转化引导计划(14-2-4-87-jch)
李娜(1991-),女,山东文登人,硕士研究生,研究方向:虾蟹生理生态学.
李玉全,E-mail:jiangfangqian@163.com
猜你喜欢
林业与生态科学(2022年4期)2022-12-24
上海蔬菜(2022年2期)2022-04-29
求学·理科版(2022年4期)2022-04-03
海洋通报(2021年3期)2021-08-14
防护林科技(2020年9期)2020-11-09
浙江海洋大学学报(自然科学版)(2020年6期)2020-07-02
中西医结合心血管病杂志(电子版)(2020年13期)2020-06-23
福建基础教育研究(2020年2期)2020-05-28
当代水产(2019年2期)2019-05-16
当代水产(2019年1期)2019-05-16
