
春季长江口近海中华鲟的食物组成
2017-09-04 01:28:58王思凯
海洋渔业 2017年4期
赵 峰,王思凯,张 涛,杨 刚,王 妤,庄 平
春季长江口近海中华鲟的食物组成
赵 峰1,2,3,王思凯1,2,3,张 涛1,2,3,杨 刚1,2,3,王 妤1,2,3,庄 平1,2,3
(1.中国水产科学研究院东海水产研究所,上海 200090;2.农业部东海与长江口渔业资源环境科学观测实验站,上海 200090;3.中国水产科学研究院长江口渔业生态重点实验室,上海 200090)
2017年3月19日,在长江口近海采集到1 ind中华鲟(Acipenser sinensis),全长133.0 cm、体质量13.8 kg,对其食物组成进行了分析。食性分析结果显示,春季中华鲟在长江口近海摄食较好,摄食强度为4级,饵料生物共有6种,其中鱼类有黄鲫(Setipinna taty)、焦氏舌鳎(Cynoglossus joyneri)和龙头鱼(Harpadon nehereus)3种,甲壳类有中华管鞭虾(Solenocera crassicornis)和口虾蛄(Oratosquilla oratoria)2种,头足类仅四盘耳乌贼(Euprymna morsei)1种。结合历史资料分析认为,中下层和底层鱼类是近海中华鲟最主要的饵料生物。
中华鲟;食物组成;近海;春季
中华鲟(Acipenser sinensis)是我国一级水生野生保护动物,在鱼类进化及生物多样性研究等方面具有重要价值。由于受到过度捕捞和涉水工程建设等影响,野生中华鲟种群处于极度濒危状态,甚至于自2013年以来已连续3年在葛洲坝下产卵场未监测到产卵活动[1-2],中华鲟物种保护研究工作引起广泛的关注和讨论[3]。研究和掌握其不同生活史阶段生物学特征及其生境需求是物种保护的基础。中华鲟是我国特有的大型江海洄游性鱼类,生活史中约95%以上的时间生活在海洋中。但是,迄今对中华鲟海洋生活史阶段特性所知甚少,仅对其在我国东南沿海的洄游分布[4-5]和食性[6]有过初步报道。2017年3月19日,在长江口近海发现误捕死亡中华鲟1 ind,研究了其食物组成,并与已有研究资料进行了综合对比分析,以期为研究掌握中华鲟海洋生活习性以及制订科学保护措施提供科学资料和参考。
1 材料和方法
样本为在长江口近海(123.08°E、30.77°N;图1)收集到误捕(流刺网)死亡中华鲟1 ind(图2)。经测量、解剖后取出消化道,对消化道中饵料生物进行鉴定分类、计数和测量,并分别计算中华鲟的肥满度和胃饱满系数[7]以及饵料生物的数量百分比(N%)、质量百分比(W%)和相对重要性指数百分比(IRI%),计算公式如下:
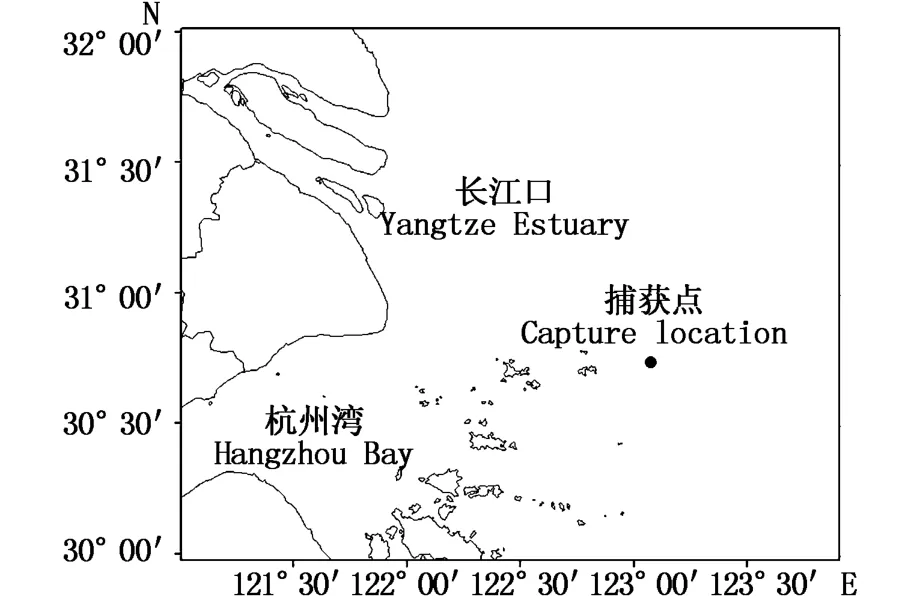
图1 采样点Fig.1 Sam p ling sites
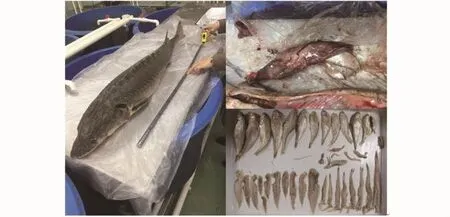
图2 中华鲟样本及其食物组成Fig.2 Sam ple of Acipenser sinensis and its diet composition

式中,F%指某种饵料生物的出现频率,因本研究中仅有中华鲟样本1 ind,故F%=100%。
2 结果与分析
2.1 肥满度与摄食强度
长江口近海中华鲟的个体大小(全长133.0 cm、体质量13.8 kg)及肥满度(0.9)均远大于长江口中华鲟幼鱼[8-10],但小于东海和黄海捕获个体[6](表1)。从摄食强度来看,本研究中华鲟与东海个体的胃充塞度均为4级,高于黄海个体;但胃饱满系数仅为33.2,远低于长江口中华鲟幼鱼(141.23~231.58)[10]。
2.2 食物组成
长江口近海中华鲟的饵料生物共有6种,其中,鱼类3种,有黄鲫(Setipinna taty)、焦氏舌鳎(Cynoglossus joyneri)和龙头鱼(Harpadon nehereus);甲壳类2种,为中华管鞭虾(Solenocera crassicornis)和口虾蛄(Oratosquilla oratoria);头足类仅四盘耳乌贼(Euprymna morsei)1种(表2;图2)。胃含物中鱼类的数量百分比、质量百分比和相对重要性指数百分比都具有绝对优势,分别达93.2%、96.7%和94.9%(表2)。
3 讨论
3.1 中华鲟的生长与肥满度
肥满度,亦称条件系数(condition factor),是鱼类体长和体重关系的一种表达方式,用作衡量鱼体的丰满程度、营养状况及其环境条件[17]。尽管无论东海或黄海中华鲟的肥满度均要高于长江口幼鱼(表1),但尚不能说明近海中华鲟在丰满程度和营养状况等方面要优于长江口幼鱼。鱼类生活史中,不同发育阶段的生长存在着等速或异速性,即鱼体各部的生长速率可以相同(等速性)或不同(异速性)[17]。研究表明,中华鲟体长的年增长量和平均增长率会随着体长的增长而呈现不同程度的递减[18]。毛翠凤等[8]研究发现,5~8月中华鲟幼鱼在长江口停留期间,其体长生长速度远大于体重生长速度。可以推断,中华鲟幼鱼入海后随着年龄和体长的增加,其体重增长速度会逐渐大于体长增长速度,这可能是近海与长江口中华鲟幼鱼肥满度差异的主要原因。近海中华鲟的肥满度及其营养状况以及环境条件的评估,还需要获取更多的研究样本及其栖息地环境因子指标进行深入的综合分析。
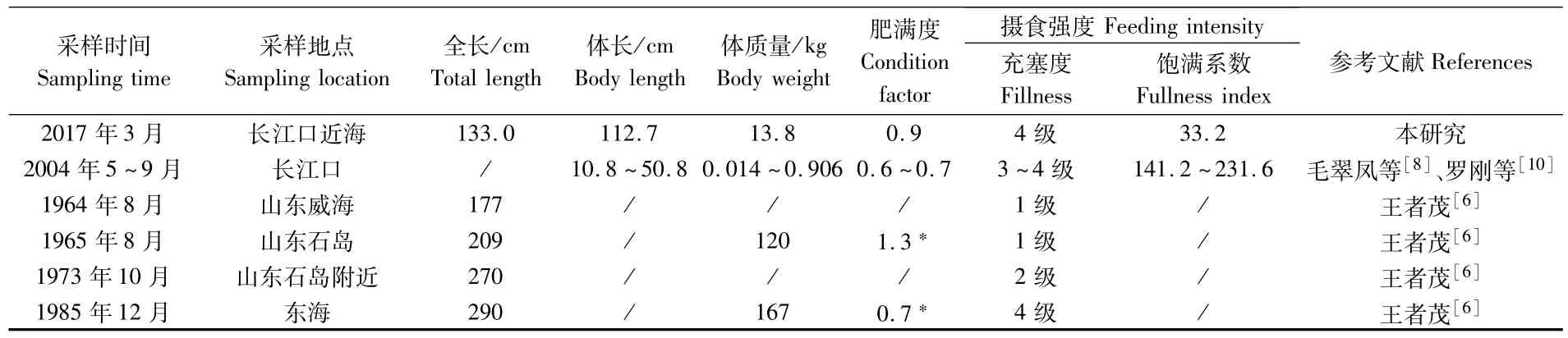
表1 中华鲟的肥满度与摄食强度Tab.1 Condition factor and feeding intensity of Acipenser sinensis
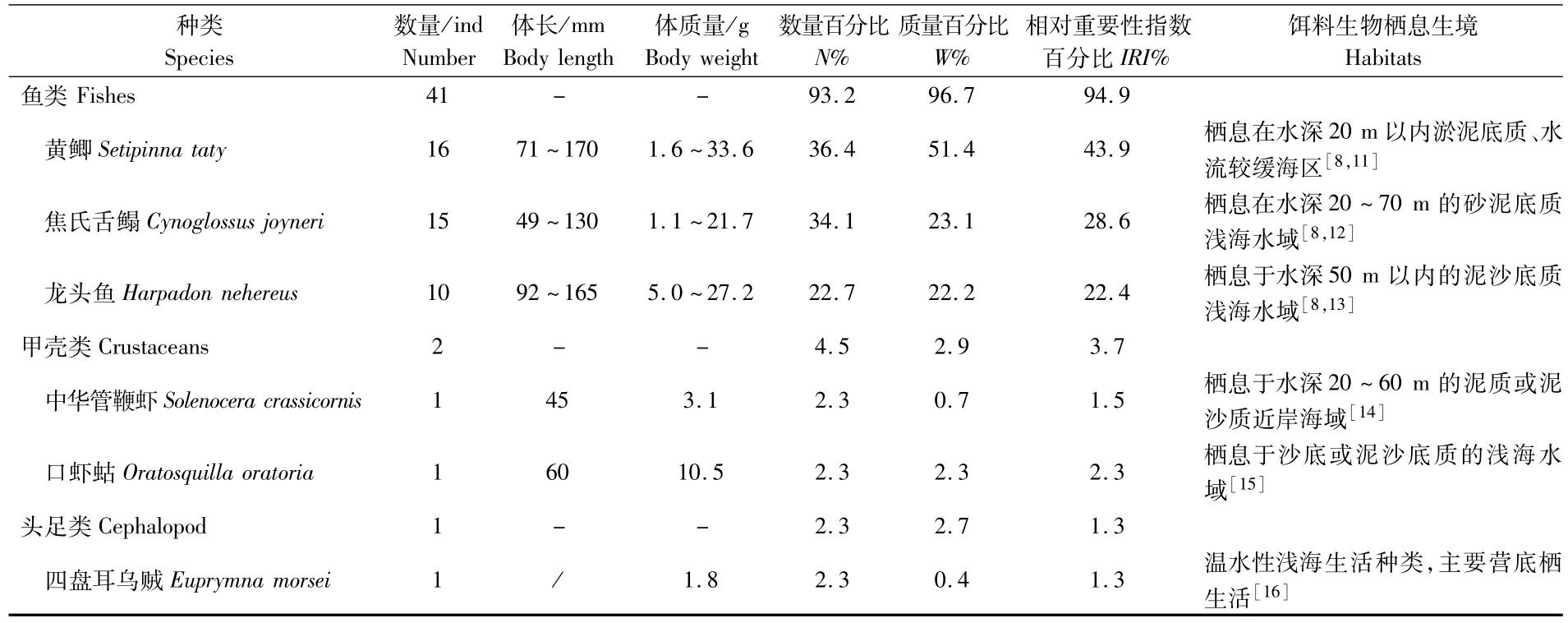
表2 春季长江口近海中华鲟的食物组成Tab.2 Food composition of Acipenser sinensis collected from the coastal waters of the Yangtze Estuary
3.2 中华鲟的摄食与食物组成
中华鲟是典型的底栖性鱼类,口裂大,下位,适于捕食底栖生物。口前部触须感知食物后,口伸长呈圆筒状把食物吸入,摄食方式属于吸吮型[6,18-19]。因此,底栖生物可能是中华鲟的主要饵料来源。
本研究发现,长江口近海中华鲟的胃饱满度达到4级,与长江口幼鱼(3~4级)[10,18]和东海成鱼(4级)[6]近似,摄食强度远高于黄海水域中华鲟的摄食强度(胃饱满度仅为1~2级)[6]。可见,长江口及东海近海无疑是中华鲟良好的索饵场。当然,不同海区间中华鲟的摄食强度与海区间饵料生物组成及其时空分布以及中华鲟不同发育阶段需求也存在着一定关系,尚需进一步加强研究。
中华鲟的食谱较为广泛,其饵料生物组成因生长发育阶段和分布水域的不同而呈现一定的差异。研究表明,长江下游(江苏常熟,浒浦江段)中华鲟稚幼鱼的饵料生物以虾蟹类为主,摄食少量黄丝藻和水生维管束植物[18];河口水域(上海崇明)中华鲟幼鱼的食物包括了12个类群的24种生物,以底栖小型鱼类、端足类和多毛类为主要食物,兼食虾类、蟹类及瓣鳃类等小型底栖动物[10];综合本研究和王者茂[6]的研究结果,洄游进入海洋生活的中华鲟,其食物包括6个类群的15种生物,主要以摄食中下层和底层的鱼类为主,其次是甲壳类和软体动物(表3)。可见,中华鲟的食性会随着生长发育阶段的不同而发生转变:由稚幼鱼阶段的以动物性食物为主的杂食性转变为亚成体和成鱼阶段的以鱼、虾蟹类为主的肉食性。另外,长江口区中华鲟幼鱼(体长10.8~50.8 cm)捕食饵料生物的最大长度不超过6.5 cm[20],小于长江口近海中华鲟(体长112.7 cm)捕食的饵料生物规格(体长4.5~17.0 cm),这说明中华鲟随着个体与摄食器官的发育完善,其捕食逐渐转向相对大型的食物类群。这一结果符合动物的“最佳索饵理论”,即捕食者总是尽可能地捕食个体较大的饵料,因为捕食大个体的饵料所获得的收益(补充的能量)要大于支出(捕食所消耗的能量),从而可以最大限度地获得能量[17,21]。而且,这种食物转换现象的出现对于扩大种群的饵料基础、满足不同发育阶段鱼类的饵料需求以及缓和它们对有限饵料资源的竞争都是十分有利的[22-23]。
3.3 中华鲟入海后的洄游分布
中华鲟入海后属索饵洄游,获得充足的饵料生物和适宜的环境条件是其洄游的最重要目的。通常,鱼类索饵洄游的特点是洄游路线、方向和时间随着饵料生物群的分布和密度、索饵鱼群数量多寡和状态而变动[17]。从近海中华鲟胃含物中饵料生物的生活习性来看,除黄鲫为中下层鱼类,其它基本属底栖生物种类,主要分布在水深不超过70 m的泥沙底质的近海浅海区(表2),且这些饵料生物在东海区生物量较高[11,13-16],这可能是中华鲟在长江口渔场和舟山渔场分布较多[18]的主要原因。陈锦辉等[4]和王友成等[5]利用PAT标志跟踪技术,研究发现中华鲟入海后沿东部沿海大陆架游动,主要分布于沿岸带15 km以内、水深11~32 m的浅海区。作者前期利用PAT标志跟踪研究也发现,中华鲟幼鱼自长江口入海后,沿东海大陆架先向南洄游至福建沿海,继而转向北洄游至黄海南部(未发表资料)。这些研究结果显示出近海中华鲟饵料生物的时空分布变化可能是其洄游分布的重要原因之一,具体尚需要进一步研究证实,这将会为中华鲟的海水保育和洄游习性研究提供重要参考资料。

表3 长江口和近海中华鲟食物组成比较Fig.3 Food comparison of Acipenser sinensis distributed in the Yangtze Estuary and the coastal area
致谢:中国水产科学研究院东海水产研究所李圣法研究员、刘勇副研究员和姜亚洲副研究员在样本信息、徐超硕士在样本测量方面给予了帮助,表示感谢。
[1] 危起伟.野生中华鲟面临灭绝风险[J].渔业致富指南,2014(20):6.
WEIQW.Extinction risk of wild Chinese sturgeon[J].FisheriesWealth Guide,2014(20):6.
[2] 李振龙.科研机构再次未监测到中华鲟自然繁殖迹象[J].中国水产,2015(2):47.LIZ L.Failed tomonitor the natural breeding of the Chinese sturgeon by scientific research institutions[J].China Fisheries,2015(2):47.
[3] ZHUANG P,ZHAO F,ZHANG T,et al.New evidence may support the persistence and adaptability of the near-extinct Chinese sturgeon[J].Biological Conservation,2016,193(2):66-69.
[4] 陈锦辉,庄 平,吴建辉,等.应用弹式卫星数据回收标志技术研究放流中华鲟幼鱼在海洋中的迁移与分布[J].中国水产科学,2011,18(2):437-442.
CHEN JH,ZHUANG P,WU JH,etal.Migration and distribution of released Acipenser sinensis in the sea based on pop-up archival tag technique[J].Journal of Fishery Sciences of China,2011,18(2):437-442.
[5] 王成友,杜 浩,刘 猛,等.厦门海域放流中华鲟的迁移和分布[J].中国科学:生命科学,2016,46(3):294-303.
WANG C Y,DU H,LIU M,et al.Migrations and distributions of Chinese sturgeon released in the sea of Xiamen[J].Scientia Sinica Vitae,2016,46(3):294-303.
[6] 王者茂.中华鲟在海中生活时期的食性初报[J].海洋渔业,1986,8(4):160-161.
WANG Z M.A preliminary report on the feeding habits of Chinese sturgeon during the sea life stage[J].Marine Fisheries,1986,8(4):160-161.
[7] 中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员会.GB/T 12763.6-2007海洋调查规范-第6部分:海洋生物调查[S].
General Administration of Quality Supervision,Inspection and Quarantine of the People’s Republic of China,Standardization Administration of the People’s Republc of China.GB/T 12763.6-2007 Specifications for Oceanographic Survey-Part 6:Marine Biological Survey[S].
[8] 毛翠凤,庄 平,刘 健,等.长江口中华鲟幼鱼的生长特性[J].海洋渔业,2005,27(3):177-181.
MAO C F,ZHUANG P,LIU J,et al.Growth of juvenile Chinese sturgeon Acipenser sinensis captured from the Yangtze River Estuary[J].Marine Fisheries,2005,27(3):177-181.
[9] 庄 平,王幼槐,李圣法,等.长江口鱼类[M].上海:上海科技出版社,2006.
ZHUANG P,WANG Y H,LIS F,et al.Fishes of the Yangtze Estuary[M].Shanghai:Shanghai Scientific&Technical Publishers,2016.
[10] 罗 刚,庄 平,章龙珍,等.长江口中华鲟幼鱼的食物组成及摄食习性[J].应用生态学报,2008,19(1):144-150.
LUO G,ZHUANG P,ZHANG L Z,et al.Diet composition and feeding habits of juvenile Acipenser sinensis in Yangtze River Estuary[J].Chinese Journal of Applied Ecology,2008,19(1):144-150.
[11] 刘 勇,程家骅,李圣法.东海区黄鲫数量分布特征的分析研究[J].海洋渔业,2004,26(4):255-260.
LIU Y,CHENG J H,LI S F.A study on the distribution of Setipinna taty in the East China Sea[J].Marine Fisheries,2004,26(4):255-260.
[12] YAMADA U,SHIRAI S,IRIE T.Names and illustrations of fishes from the East China Sea and the Yellow Sea:Japanese Chinese Korean[J].Overseas Fishery Cooperation Foundation,1995:288.
[13] 林龙山.东海区龙头鱼数量分布及其环境特征[J].上海海洋大学学报,2009,18(1):66-71.
LIN L S.Spatial distribution and environmental characteristics of Harpodon nehereus in the East China Sea region[J].Journal of Shanghai Ocean University,2009,18(1):66-71.
[14] 宋海棠,俞存根,薛利健,等.东海经济虾蟹类[M].北京:海洋出版社,2006.[M].北京:海洋出版社,2006.
SONG H T,YU C G,XUE L J,et al.The East China Sea economic shrimps and crabs[M].Beijing:China Ocean Press,2006.
[15] 俞存根,陈全震,陈小庆,等.舟山渔场及邻近海域虾蛄类的种类组成和数量分布[J].大连海洋大学学报,2011,26(2):153-156.
YU C G,CHEN Q Z,CHEN X Q,et al.Species and composition and quantity profile ofmantis shrimp around Zhoushan fishing ground[J].Journal of Dalian Ocean University,2011,26(2):153-156.
[16] 宋海棠,丁天明,徐开达.东海经济头足类资源[M].北京:海洋出版社,2009.
SONG H T,DING T M,XU K D.The East China Sea economic Cephalopod resource[M].Beijing:China Ocean Press,2009.
[17] 殷名称.鱼类生态学[M].北京:中国农业出版社,1995.
YIN M C.Fish Ecology[M].Beijing:China Agriculture Press,1995.
[18] 四川省长江水产资源调查组.长江鲟鱼类生物学及人工繁殖研究[M].成都:四川科学技术出版社,1988.
The Changjiang Aquatic Resources Survey Group,Sichuan Province.The biology of the sturgeons in Changjiang and their artificial propagation[M].Chengdu:Sichuan Scientific and Technical Publishing House,1988.
[19] 庄 平,章龙珍,罗 刚,等.长江口中华鲟幼鱼感觉器官在摄食行为中的作用[J].水生生物学报,2008,32(4):475-481.
ZHUANG P,ZHANG L Z,LUO G,et al.Function of sense organs to the feeding behavior of juvenile Chinese sturgeon captured from the Yangtze Estuary[J].Acta Hydrobiologica Sinica,2008,32(4):475-481.
[20] 罗 刚.长江口中华鲟幼鱼的食物组成、食物竞争和摄食行为[D].上海:上海海洋大学,2007.
LUO G.Food composition,feeding competition and feeding behavior of juvenile Chinese sturgeon,Acipenser sinensis,in the Yangtze River Estuary[D].Shanghai:Shanghai Ocean University,2007.
[21] GERKING SD.Feeding ecology of fish[M].San Diego:Academic Press,1994.
[22] BROOKS J L,DODSON S I.Predation,body size,and composition of plankton[J].Science,1965,150(3692):28.
[23] GROSSMAN G D.Ecological aspects of ontogenetic shifts in prey size utilization in the bay goby(Pisces:Gobiidae)[J].Oecologia,1980,47(2):233-238.
Food com position of Acipenser sinensis in the coastal waters of the Yangtze Estuary in spring
ZHAO Feng1,2,3,WANG Si-kai1,2,3,ZHANG Tao1,2,3,YANG Gang1,2,3,WANG Yu1,2,3,ZHUANG Ping1,2,3
(1.East China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Shanghai 200090;2.Scientific Observing and Experimental Station of Fisheries Resources and Environment of East China Sea and Yangtze Estuary,Ministry of Agriculture,Shanghai 200090;3.Key Laboratory of Fishery Ecology of the Yangtze Estuary,Chinese Academy of Fishery Sciences,Shanghai 200090)
One Chinese sturgeon(Acipenser sinensis)was collected in the coastal waters of the Yangtze Estuary on the 19th of March,2017,and its food composition was analyzed.The results showed that A.sinensis fed very well in the coastal waters of the Yangtze Estuary in spring,and its feeding intensity was at level four.There were six diet species in the fish stomach.Among them,there were three fish species(i.e.Setipinna taty,Cynoglossus joyneri,and Harpadon nehereus),two crustacean species(i.e.Solenocera crassicornis and Oratosquilla oratoria),and one cephalopod species(Euprymna morsei).The middle-and bottom-living fishes were the dominant prey species of Chinese sturgeon in the coastal waters based on the present and previous studies.
Acipenser sinensis;food composition;coastal waters of the Yangtze Estuary;spring
Q 178
A
1004-2490(2017)04-0427-06
2017-04-06
中国水产科学研究院基本科研业务费(2015A01YY01);农业部财政专项(2130135-探捕);修复向家坝库区渔业资源及保护长江珍稀特有物种合作项目(F13);香港海洋公园保育基金(FH04.1718)
赵 峰(1978-),男,副研究员,主要从事渔业生态与保护生物学研究。E-mail:zhaof@ecsf.ac.cn
庄 平,研究员。E-mail:pzhuang@ecsf.ac.cn
猜你喜欢
中国三峡(2017年4期)2017-06-06 10:44:23
中国三峡(2017年4期)2017-06-06 10:44:22
中国三峡(2017年4期)2017-06-06 10:44:22
中国三峡(2016年5期)2017-01-15 13:58:47
海洋渔业(2016年6期)2016-04-16 03:00:35
海洋渔业(2016年6期)2016-04-16 03:00:32
动物营养学报(2015年10期)2015-12-01 03:30:54
动物营养学报(2015年10期)2015-12-01 02:26:22
应用海洋学学报(2014年1期)2014-11-22 07:17:50
华东师范大学学报(自然科学版)(2014年3期)2014-03-11 16:18:19
