
针对猪瘟病毒E2蛋白的嵌合猪源化单克隆抗体的表达及抗病毒活性鉴定
2017-09-03 08:50陈姝承孙慧敏李素刘平黄马吉飞仇华吉
生物工程学报 2017年8期
陈姝承,孙慧敏,李素,刘平黄,马吉飞,仇华吉
1 天津农学院 动物科学与动物医学学院,天津 300384 2 中国农业科学院哈尔滨兽医研究所,黑龙江 哈尔滨 150069
针对猪瘟病毒E2蛋白的嵌合猪源化单克隆抗体的表达及抗病毒活性鉴定
陈姝承1,2,孙慧敏2,李素2,刘平黄2,马吉飞1,仇华吉1,2
1 天津农学院 动物科学与动物医学学院,天津 300384 2 中国农业科学院哈尔滨兽医研究所,黑龙江 哈尔滨 150069
陈姝承, 孙慧敏, 李素, 等. 针对猪瘟病毒E2蛋白的嵌合猪源化单克隆抗体的表达及抗病毒活性鉴定. 生物工程学报,2017, 33(8): 1235–1243.
Chen SC, Sun HM, Li S, et al. Expression and antiviral activity of a chimeric porcinized monoclonal antibody (cHQ06)against E2 protein of classical swine fever virus. Chin J Biotech, 2017, 33(8): 1235–1243.
猪瘟 (Classical swine fever,CSF) 是严重危害养猪业的一种烈性传染病,常造成巨大的经济损失,是世界动物卫生组织要求必须申报的动物疫病之一。猪瘟的病原是猪瘟病毒 (Classical swine fever virus,CSFV),CSFV的结构蛋白由衣壳蛋白 (C) 和囊膜糖蛋白 (Erns、E1、E2) 构成。E2蛋白是CSFV主要的保护性抗原,可以诱导机体产生中和抗体,从而抵抗CSFV的感染。此前,本团队制备了一株针对CSFV E2蛋白的鼠源单克隆抗体HQ06。文中将HQ06抗体重链和轻链可变区基因与猪源恒定区基因嵌合后克隆至真核表达载体,利用中国仓鼠卵巢 (CHO) 细胞制备一株针对 CSFV E2蛋白的嵌合猪源化单克隆抗体 cHQ06。应用ELISA、Western blotting试验证实了cHQ06与CSFV E2蛋白具有良好的反应性;中和试验结果表明cHQ06可以中和CSFV。综上所述,本研究应用CHO细胞稳定表达了具有良好反应性和中和活性的针对CSFV E2蛋白的嵌合猪源化单克隆抗体cHQ06,为研究CSFV E2蛋白结构、功能以及开发新型的CSFV诊断和治疗制剂奠定基础。
猪瘟病毒,单克隆抗体,基因工程抗体,嵌合表达,悬浮培养
猪瘟 (Classical swine fever,CSF) 是严重危害养猪业的一种烈性传染病,常造成巨大的经济损失,该病被世界动物卫生组织 (OIE) 列为必须申报的一类动物疫病[1]。CSF的病原是猪瘟病毒 (Classical swine fever virus,CSFV),CSFV是有囊膜的单股正链RNA病毒,基因组长约 12.3 kb,是黄病毒科瘟病毒属的成员[2]。该基因组仅编码一个大的开放阅读框,其编码一个由3 898个氨基酸组成的多聚蛋白。该多聚蛋白被宿主细胞编码的信号肽酶和自身编码的蛋白酶Npro、NS2及NS3切割为4种结构蛋白 (C和Erns、E1、E2) 及8种非结构蛋白 (Npro、P7、NS2、NS3、NS4A、NS4B、NS5A 和 NS5B)[3-4]。
衣壳蛋白C和糖蛋白Erns、E1、E2是CSFV的结构蛋白[5]。E2为Ⅰ型跨膜蛋白,其氨基端为胞外区,羧基端为跨膜区,将E2蛋白锚定在囊膜上[6]。E2蛋白在病毒生命周期中有重要作用,影响病毒吸附[7]、组织嗜性[8]和病毒毒力[9]。此外,E2是CSFV的主要保护性抗原蛋白,诱导机体产生中和抗体来抵抗病毒的感染[10]。E2蛋白在氨基端有B、C、D、A四个抗原结构域,构成两个独立的抗原区,按B/C和A/D的顺序排列 (aa 690–800 和 766–865)[11-13]。
E2蛋白的抗原特性已经被许多单克隆抗体所鉴定[14-16]。在本实验室前期的工作中,我们制备了一株鼠源E2蛋白单克隆抗体HQ06,它可以特异性识别CSFV C株和石门株E2蛋白的线性表位,该单克隆抗体重链为免疫球蛋白 G(IgG) 1型、轻链为к型[17]。
为了克服产生单克隆抗体的杂交瘤细胞不稳定且抗性易丢失等问题,在本研究中,我们利用单细胞克隆技术获得HQ06重链 (HC)、轻链 (LC) 的基因,将其与猪源抗体恒定区基因相融合,克隆至真核表达载体,并在CHO细胞上稳定、高效表达针对CSFV E2蛋白的嵌合猪源化单克隆抗体 (cHQ06)。通过检测实验证实cHQ06与CSFV E2蛋白具有良好的反应活性。而且,通过中和试验证实其可以抑制CSFV的感染。因此,本研究证明,针对CSFV E2蛋白的嵌合猪源化单克隆抗体cHQ06可以稳定、高效表达,为研究 CSFV E2蛋白结构、功能以及开发新型的 CSFV诊断和治疗制剂奠定基础。
1 材料与方法
1.1 细胞、病毒和质粒
人胚肾细胞 (HEK293T)、猪肾细胞 (PK-15)购自美国标准菌种收藏中心;中国仓鼠卵巢细胞 (CHO) 购于中国科学院细胞库。悬浮HEK293细胞由哈尔滨动物生物制品国家工程研究中心有限公司提供。CSFV石门株在PK-15细胞中传代并用Reed-Muench公式计算病毒滴度[18]。慢病毒载体pFUGW及辅助质粒psPAX2、pMD2.G由 Addgene提供。表达抗体重链和轻链的真核表达质粒pcDNA-HC和pcDNA-LC由哈尔滨兽医研究所猪烈性传染病创新团队保存。
1.2 主要试剂
Prime STAR®HS DNA聚合酶、限制性内切酶BamHⅠ、EcoRⅠ、T4 DNA连接酶、DNA 上样缓冲液等购自TaKaRa公司;目的质粒特异性引物由博仕生物公司合成。HiTrap Protein A HP预装柱 (5 mL) 购于GE Healthcare公司;分子量为10 kDa的超滤管购于Merck公司。辣根过氧化物酶 (HRP) 标记的兔抗猪免疫球蛋白 G(IgG) H&L (Abcam 公司);鼠抗猪 IgG (BD Pharmingen公司);HRP标记的山羊抗鼠的IgG、3,3′,5,5′-四甲基联苯胺 (TMB) 底物、4′,6-二脒基-2-苯基吲哚 (DAPI)、伊文斯蓝 (Evans blue)、异硫氰酸荧光素 (FITC) 标记的 IgG均购于Sigma-Aldrich公司。
1.3 重组质粒的构建
以pcDNA-HC和pcDNA-LC质粒为模板,扩增cHQ06重链 (HC) 和轻链 (LC) 基因。琼脂糖凝胶电泳鉴定核酸分子量正确后,将目的条带胶回收并于37 ℃酶切4 h,将其与同样酶切后的pFUGW质粒载体于16 ℃连接8 h,制备重组质粒pFUGW-HC和pFUGW-LC。引物序列见表1。
1.4 共表达 cHQ06抗体重链和轻链的细胞系的构建
将质粒pFUGW-HC和pFUGW-LC以及辅助质粒pMD2.G和psPAX2共转染到HEK293T细胞中,制备表达抗体HC和LC的慢病毒。将重组慢病毒按照感染复数 (MOI) 为 10感染CHO细胞。48 h之后对细胞计数,应用有限稀释法筛选单细胞克隆,收取其上清用双抗体夹心酶联免疫吸附试验 (DAS-ELISA) 进行检测,通过检测结果筛选出cHQ06抗体细胞系。
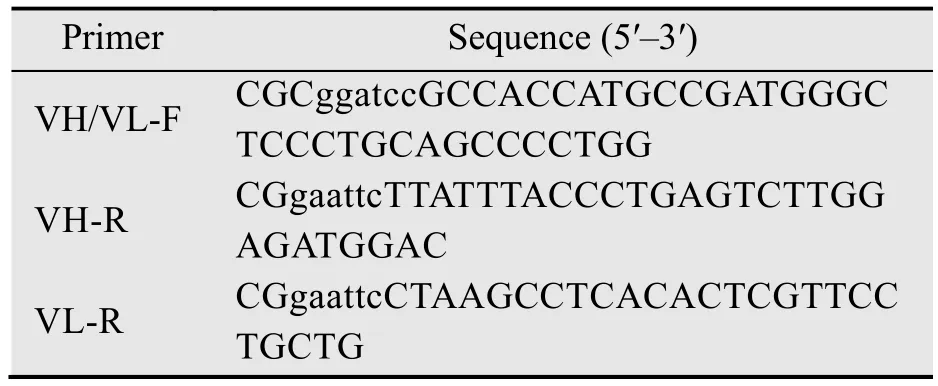
表1 cHQ06-HC和cHQ06-LC特异性引物序列Table 1 Specific primer sequences for amplifying cHQ06-HC and cHQ06-LC
1.5 cHQ06的表达和纯化
收集CHO-cHQ06细胞系培养上清,使用制备型液相层析系统 ÄKTA explorer 对抗体进行纯化。蛋白洗脱后应用非还原性8%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳 (SDS-PAGE) 检测抗体组装,并应用Western blotting等方法分析cHQ06的组装以及与兔抗猪IgG的反应性。
1.6 DAS-ELISA
将纯化的鼠抗猪IgG经1∶250稀释,在4 ℃条件下,以每孔100 μL包被ELISA板12 h。包被完成后,每孔加入150 µL PBST清洗1次,然后将含有5% FBS的5%脱脂乳加入到ELISA孔板中,每孔100 μL,37 ℃作用2 h。将纯化的cHQ06经2倍系列稀释加入到孔板中,于37 ℃作用1 h后,弃掉抗体,用PBST洗涤3次。将板子中的 PBST吸干,加入用辣根过氧化物酶(HRP) 标记的兔抗猪 IgG H&L (1∶4 000),于37 ℃继续孵育1 h。应用PBST洗涤3次后加入TMB底物显色,15 min后加入2 mol/L H2SO4终止反应,最后应用酶标仪在450 nm波长处检测吸光度。
1.7 应用间接ELISA检测cHQ06与 CSFV E2蛋白的反应活性
分别应用BHK-21细胞表达的CSFV石门株E2蛋白、悬浮HEK293细胞表达的C株E2蛋白或纯化的CSFV 石门株包被ELISA板,每孔包被4 ng。在4 ℃放置12 h,然后用5%脱脂乳于37 ℃封闭2 h。将纯化的cHQ06经2倍系列稀释加入孔板中,于37 ℃孵育1 h。应用PBST洗涤3次,加入用HRP标记的兔抗猪IgG H&L(1∶4 000),于37 ℃孵育1 h,TMB底物显色后应用酶标仪在450 nm波长处检测吸光度。
1.8 应用 Western blotting检测 cHQ06与CSFV E2蛋白反应活性
将BHK-21细胞表达的CSFV石门株E2蛋白和悬浮的HEK293细胞表达的C株E2蛋白进行SDS-PAGE,将蛋白转印至硝酸纤维素 (NC)膜后,用5%脱脂乳于室温孵育2 h。应用PBS洗涤1次后,将NC膜放至HQ06腹水 (1∶1 000)及纯化的cHQ06中,室温孵育1.5 h。用PBST洗涤 5次,每次 5 min,洗涤完毕后,分别于HRP标记的山羊抗鼠 (1∶1 000)、兔抗猪 IgG(1∶2 500) 中室温孵育1 h。应用PBST洗涤后,使用多功能荧光化学发光成像分析系统进行扫描,保存灰白图。
1.9 病毒中和试验
将纯化的cHQ06抗体2倍系列稀释,做6个稀释梯度和4个重复,与每孔中100 TCID50的CSFV石门株等体积混合,于37 ℃温箱中孵育2 h。应用冷无水乙醇于-20 ℃冰箱中固定细胞15 min,然后将CSFV阳性血清1∶100稀释,加入每孔,于37 ℃孵育2 h后,应用PBS洗涤5次,每次5 min。洗涤结束后加入1∶100稀释的FITC标记的IgG和1∶1 000稀释的伊文斯蓝,放入37 ℃温箱,孵育1 h。孵育结束应用PBST清洗5次,再加入1∶1 000稀释的DAPI将细胞核染色,37 ℃放置10 min,用PBS清洗5次后,应用荧光显微镜分析抗体中和效价。
1.10 统计学分析
应用GraphPad Prism 5软件分析所有的数据。误差线表示图中所有平均值()的标准偏差(s)。
2 结果与分析
2.1 cHQ06重链、轻链重组质粒的构建
以质粒pcDNA-HC和pcDNA-LC为模板,分别应用特异性引物分别扩增 cHQ06-HC、LC目的基因,如图1A所示,重链大小为1 500 bp,轻链大小为 750 bp,表明目的片段已被成功克隆。重组质粒经BamHⅠ和EcoRⅠ双酶切后在1 500 bp和750 bp处出现特异条带,测序结果显示,其与目标序列一致。结果表明,质粒pFUGW-HC和pFUGW-LC构建成功 (图1B)。
2.2 过表达cHQ06细胞系的筛选
将转导后的细胞接种于96孔板中,应用有限稀释法筛选单克隆细胞,显微镜下共观察到有26株单克隆细胞,收取其上清,应用双抗体夹心ELISA检测抗体与兔抗猪IgG的反应性。检测结果表明,6号、21号和25号细胞克隆的培养上清与兔抗猪的 IgG反应性较强,有可能为阳性细胞 (图2A)。将这3株细胞扩大培养,收取上清进行纯化,双抗体夹心ELISA检测结果显示,6号细胞克隆的培养上清与兔抗猪 IgG反应性最好,且呈剂量依赖性 (图 2B)。
2.3 cHQ06猪源抗体活性的检测
将纯化后的6号细胞克隆的培养上清 (浓度为1 mg/mL) 进行非还原性的SDS-PAGE分析,结果表明 cHQ06抗体可成功组装成为 160 kDa大小的蛋白,与猪源抗体蛋白的分子量一致 (图3A)。为了进一步证明cHQ06抗体是否是猪源化抗体,我们应用 8%非还原性 SDS-PAGE进行Western blotting试验,在160 kDa处检测到抗体蛋白,这表明,cHQ06可以和抗猪IgG抗体反应(图3B)。以上结果证明,cHQ06抗体成功组装,并且可被兔抗猪 IgG识别,已成功获得猪源化的E2蛋白单克隆抗体。
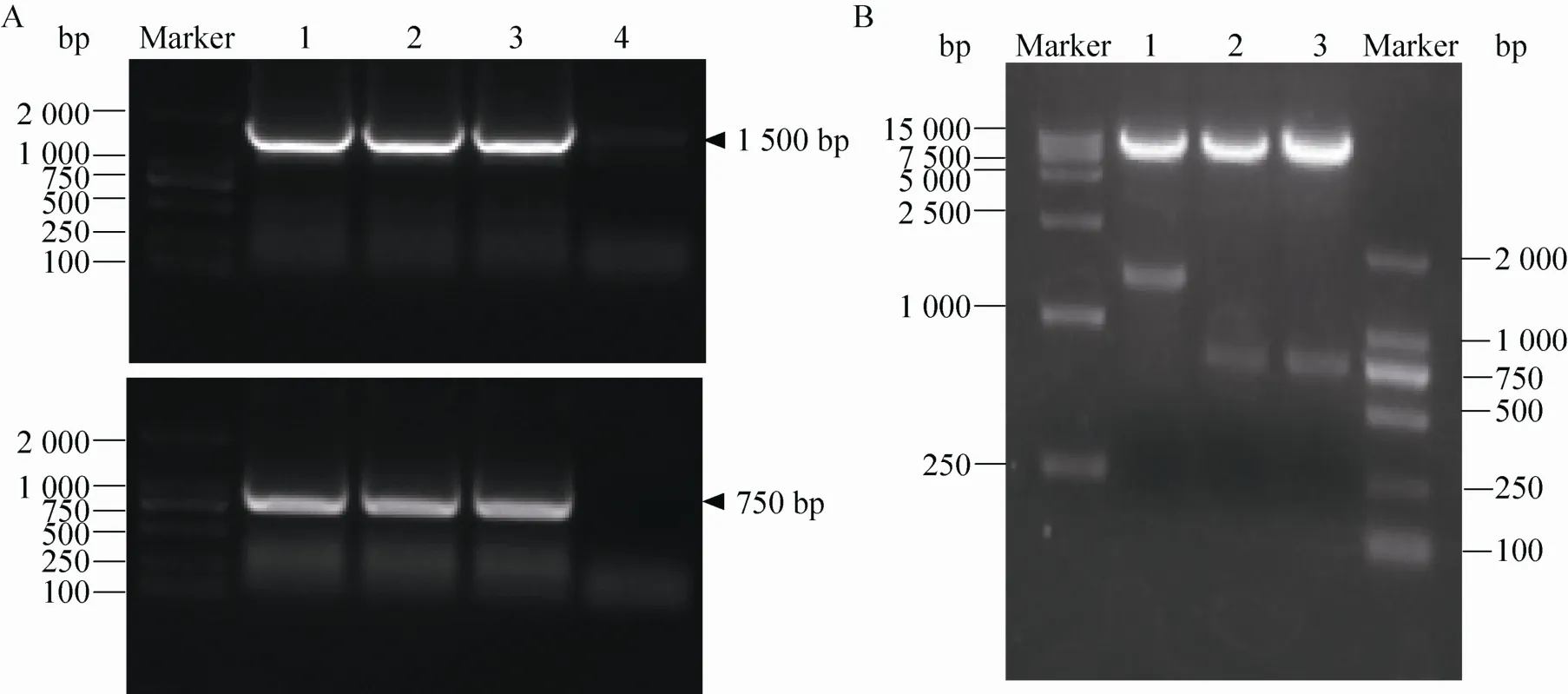
图1 重组质粒pFUGW-HC和pFUGW-LC的构建Fig. 1 Construction of recombinant plasmids pFUGW-HC and pFUGW-LC. (A) Amplification results of cHQ06 heavy chain and light chain by RT-PCR; 1–3: the gene of HC and LC; 4: negative control. (B) Identification of plasmids pFUGW-HC and pFUGW-LC; 1: pFUGW-HC; 2: pFUGW-LC; 3: pFUGW.
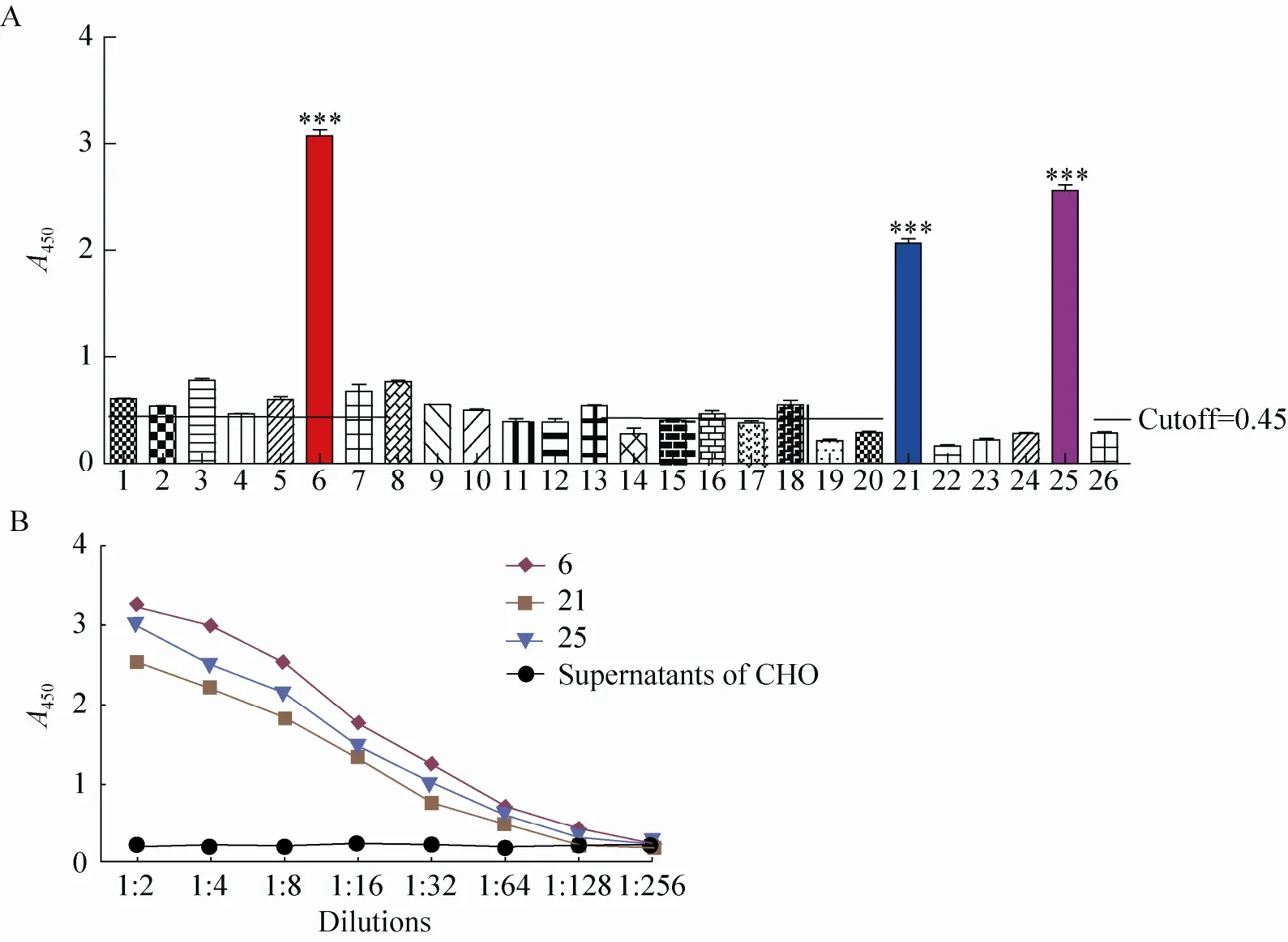
图2 cHQ06细胞系的筛选Fig. 2 Screening of cHQ06 cell line. (A) Screening of positive cell clones of cHQ06 using double-antibody sandwich ELISA. (B) Reactivity of different cell clones of cHQ06 with rabbit anti-pig IgG in a dose-dependent manner.
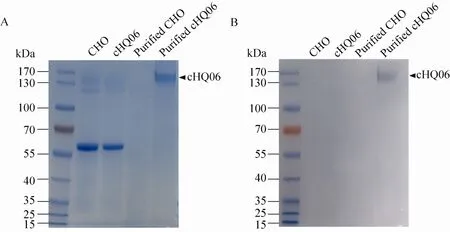
图3 cHQ06猪源抗体活性的检测Fig. 3 Activity measurement of porcinized antibody cHQ06. (A) The generation of cHQ06 was examined using SDS-PAGE. (B) The reactivity of cHQ06 and rabbit anti-pig IgG was examined using Western blotting.
2.4 cHQ06反应性的鉴定
应用间接 ELISA和 Western blotting验证cHQ06抗体和 CSFV E2蛋白的反应性。间接ELISA结果表明,cHQ06抗体与不同表达系统表达的E2蛋白均具有良好的反应性,并且呈剂量依赖性 (图 4A)。为了进一步证明 cHQ06抗体的特异性,应用 Western blotting试验检测CSFV C株和石门株E2蛋白与cHQ06抗体的反应性,并用鼠源HQ06 (浓度为1 mg/mL) 作为阳性对照。结果表明,cHQ06抗体识别CSFV E2蛋白的能力强于亲本抗体HQ06 (图4B),综上所述,cHQ06抗体与CSFV E2蛋白有良好的反应性。
2.5 cHQ06抗体对猪瘟病毒的中和作用
HQ06可以识别 CSFV E2蛋白表面的772LFDGTNP778表位[19]。为了鉴别 cHQ06是否具有中和病毒的能力,将CSFV以100 TCID50与等体积的2倍系列稀释cHQ06抗体进行孵育,然后感染PK-15细胞,48 h后进行间接免疫荧光试验 (IFA)。结果显示,在 1∶2、1∶4和 1∶8的cHQ06稀释孔中,未检测到CSFV的特异性荧光灶,且在1∶16的cHQ06稀释孔中,只检测到少量荧光灶,这表明,cHQ06中和效价为1∶12 (图5)。
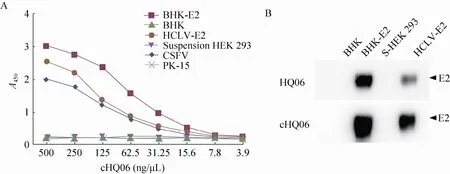
图4 cHQ06抗体的反应性检测Fig. 4 Detection of the reactivity of cHQ06 with the CSFV E2 protein. (A) Indirect ELISA. (B) Western blotting.

图5 用中和试验检测cHQ06抗体对CSFV的中和作用Fig. 5 Neutralizing activity of cHQ06 against CSFV infection determined by neutralization test. Scale bars represent 400 μm.
3 讨论
在蛋白纯化方面,我们使用制备型液相层析系统ÄKTA explorer纯化抗体蛋白,现已有很多应用ÄKTA纯化蛋白的报道[24-25]。传统的蛋白纯化方法费时、费力,且蛋白容易降解,洗脱效果不佳。ÄKTA explorer相比于传统纯化蛋白的方法更高效,可在短时间内,在大量流入样品的情况下,根据紫外峰值快速精准地收取纯化的目的蛋白,最大程度地避免抗体蛋白的降解。
在中和试验中,我们应用 IFA进行检测,确定了该抗体的中和效价。试验方法参考了欧盟猪瘟诊断标准操作手册,该抗体在 1 mg/mL浓度下的中和效价为1∶12,中和效价不高,可能是由于单克隆抗体只能识别一个抗原表位,所以中和效价偏低。
我们已经证实了该抗体的反应性和抑制病毒复制的能力,并且有较好的亲和力。因此,cHQ06在检测方面的应用具有很高的研究价值。在下一步研究中,我们计划应用cHQ06抗体建立竞争ELISA检测方法,为开发新型CSFV抗体检测试剂奠定基础。
REFERENCES:
[1] Ji W, Guo Z, Ding NZ, et al. Studying classical swine fever virus: making the best of a bad virus.Virus Res, 2015, 197: 35–47.
[2] Becher P, Ramirez RA, Orlich M, et al. Genetic and antigenic characterization of novel pestivirus genotypes: implications for classification.Virology, 2003, 311(1): 96–104.
在知识经济时代,教育是实现社会主义现代化的基础,肩负着提高劳动者素质和培养专门人才的重要任务。在乡村振兴战略中,师生首先要对乡村振兴的地位与作用、意义与内涵有较深的认识与了解;第二要有吃苦的工作精神和踏实的干劲;第三要有良好的理论水平和自觉的动手能力;第四要具有与乡村干部和广大农民进行沟通的能力;第五要有团队精神。只有这五点做到了,才能做好做实乡村振兴中的事情,而这正是同学们综合素质的体现。
[3] Collett MS, Moennig V, Horzinek MC. Recent advances in pestivirus research. J Gen Virol, 1989,70(2): 253–266.
[4] Thiel HJ, Stark R, Weiland E, et al. Hog cholera virus: molecular composition of virions from a pestivirus. J Virol, 1991, 65(9): 4705–4712.
[5] Rümenapf T, Meyers G, Stark R, et al. Molecular characterization of hog cholera virus. Arch Virol(Suppl), 1991, 3: 7–18.
[6] Li Y, Wang JM, Kanai R, et al. Crystal structure of glycoprotein E2 from bovine viral diarrhea virus.Proc Natl Acad Sci USA, 2013, 110(17):6805–6810.
[7] Wang Z, Nie YC, Wang PG, et al. Characterization of classical swine fever virus entry by using pseudotyped viruses: E1 and E2 are sufficient to mediate viral entry. Virology, 2004, 330(1):332–341.
[8] Reimann I, Depner K, Trapp S, et al. An avirulent chimeric Pestivirus with altered cell tropism protects pigs against lethal infection with classical swine fever virus. Virology, 2004, 322(1):143–157.
[9] Risatti GR, Borca MV, Kutish GF, et al. The E2 glycoprotein of classical swine fever virus is a virulence determinant in swine. J Virol, 2005,79(6): 3787–3796.
[10] Beer M, Goller KV, Staubach C, et al. Genetic variability and distribution of classical swine fever virus. Anim Health Res Rev, 2015, 16(1): 33–39.
[11] Hulst MM, Westra DF, Wensvoort G, et al.Glycoprotein E1 of hog cholera virus expressed in insect cells protects swine from hog cholera. J Virol, 1993, 67(9): 5435–5442.
[12] König M, Lengsfeld T, Pauly T, et al. Classical swine fever virus: independent induction of protective immunity by two structural glycoproteins. J Virol, 1995, 69(10): 6479–6486.
[13] Wensvoort G, Terpstra C, de Kluijver EP, et al.Antigenic differentiation of pestivirus strains with monoclonal antibodies against hog cholera virus.Vet Microbiol, 1989, 21(1): 9–20.
[14] Sandomenico A, Leonardi A, Berisio R, et al.Generation and characterization of monoclonal antibodies against a cyclic variant of hepatitis C virus E2 epitope 412–422. J Virol, 2016, 90(7):3745–3759.
[15] Angus AG, Patel AH. Immunotherapeutic potential of neutralizing antibodies targeting conserved regions of the HCV envelope glycoprotein E2.Future Microbiol, 2011, 6(3): 279–294.
[16] Edward S, Sands JJ. Antigenic comparisons of hog cholera virus isolates from Europe, America and Asia using monoclonal antibodies. Dtsch Tierarztl Wochenschr, 1990, 97(2): 79–81.
[17] Hou Q, Peng WP, Sun Y, et al. Expression of the truncated E2 protein-encoding gene of classical swine fever virus in Escherichia coli and preparation of a monoclonal antibody against E2 protein. Chin Vet Sci, 2008, 38(1): 1–5 (in Chinese).侯强, 彭伍平, 孙元, 等. 猪瘟病毒 E2蛋白主要抗原区编码基因的原核表达及其单克隆抗体的制备. 中国兽医科学, 2008, 38(1): 1–5.
[18] Reed LJ, Muench H. A simple method of estimating fifty percent endpoints. Am J Epidemiol,1938, 27(3): 493–497.
[19] Peng WP, Hou Q, Xia ZH, et al. Identification of a conserved linear B-cell epitope at the N-terminus of the E2 glycoprotein of classical swine fever virus by phage-displayed random peptide library. Virus Res, 2008, 135(2): 267–272.
[20] Steplewski Z, Sun LK, Shearman CW, et al.Biological activity of human-mouse IgG1, IgG2,IgG3, and IgG4 chimeric monoclonal antibodies with antitumor specificity. Proc Natl Acad Sci USA, 1988, 85(13): 4852–4856.
[21] Shaw DR, Khazaeli MB, Sun LK, et al.Characterization of a mouse/human chimeric monoclonal antibody (17-1A) to a colon cancer tumor-associated antigen. J Immunol, 1987,138(12): 4534–4538.
[22] Hashimoto Y, Tada M, Iida M, et al. Generation and characterization of a human-mouse chimeric antibody against the extracellular domain of claudin-1 for cancer therapy using a mouse model.Biochem Biophys Res Commun, 2016, 477(1):91–95.
[23] Ji S, Jiang M, Yan B, et al. The chimeric monoclonal antibody MHCSZ-123 against human von Willebrand factor A3 domain inhibits high-shear arterial thrombosis in a Rhesus monkey model. J Hematol Oncol, 2017, 10(1): 111.
[24] Yoo D, Provchy J, Park C, et al. Automated high-throughput protein purification using an ÄKTA purifier and a CETAC autosampler. J Chromatogr A, 2014, 1344: 23–30.
[25] Wang FM, Yu F, Zhao MH. A method of purifying intact complement factor H from human plasma.Protein Expr Purif, 2013, 91(2): 105–111.
(本文责编 陈宏宇)
Expression and antiviral activity of a chimeric porcinized monoclonal antibody (cHQ06) against E2 protein of classical swine fever virus
Shucheng Chen1,2, Huimin Sun2, Su Li2, Pinghuang Liu2, Jifei Ma1, and Huaji Qiu1,2
1 College of Animal Science and Veterinary Medicine, Tianjin Agricultural University, Tianjin 300384, China 2 Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences (CAAS), Harbin 150069, Heilongjiang, China
Classical swine fever (CSF), one of OIE-listed diseases, is a highly contagious and economically important disease of pigs. Classical swine fever virus (CSFV) is the causative agent of CSF. The capsid (C) protein and the glycoproteins Erns, E1 and E2, are structural components of the virus. E2 is the most immunogenic protein of the CSFV glycoproteins, inducing neutralizing antibodies that provide protection against lethal CSFV challenge. In a previous study,we developed a murine MAb HQ06 against the E2 protein of CSFV. In this study, the variable region genes from HQ06 and constant regions gene of swine antibody are fused and cloned into the eukaryotic expression vectors to establish a cell line which can stably express a chimeric porcinized MAb (cHQ06) against E2 in CHO cell. The purified cHQ06 antibody protein was determined to be successfully generated, which exhibited high reactivity between cHQ06 and the E2 protein of CSFV by enzyme-linked immunosorbent assay (ELISA) and Western blotting. More importantly, we investigated the neutralizing activity of cHQ06 against CSFV. In conclusion, this study generated cHQ06 for efficient and stable production which can be used against to develop novel diagnostic assays, investigate the structure and function of the E2 protein and generate novel preparations of diagnosis and treatment.
classical swine fever virus, monoclonal antibody, genetic engineering antibody, chimeric expression, suspension culture
June 24, 2017; Accepted: July 25, 2017
s:Huaji Qiu. Tel/Fax: +86-451-51051708; E-mail: huajiqiu@hvri.ac.cn Jifei Ma. Tel: +86-22-23784590; E-mail: hbmjfts@126.com
Supported by: National Natural Science Foundation of China (No. 31672537).
国家自然科学基金(No. 31672537) 资助。
猜你喜欢
今日农业(2022年14期)2022-09-15
现代畜牧科技(2021年9期)2021-10-13
猪业科学(2021年3期)2021-05-21
中国现代医药杂志(2020年10期)2020-12-14
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
现代检验医学杂志(2016年3期)2016-11-15
湖南畜牧兽医(2016年3期)2016-06-05
华南农业大学学报(2015年5期)2015-12-04
医学研究杂志(2015年3期)2015-06-10
