
厚壳贻贝在不同温度下的干露耐受性研究
2017-08-16 10:23:11孙煜阳童巧琼詹萍萍徐善良王春琳
生物学杂志 2017年4期
孙煜阳, 童巧琼, 王 文, 詹萍萍, 程 亮, 徐善良, 王春琳
(宁波大学 海洋学院, 宁波 315211)
厚壳贻贝在不同温度下的干露耐受性研究
孙煜阳, 童巧琼, 王 文, 詹萍萍, 程 亮, 徐善良, 王春琳
(宁波大学 海洋学院, 宁波 315211)
为了解不同温度及保湿与干燥状态下厚壳贻贝稚贝的耐干露能力,分别在低温1℃、2℃、4℃和8℃,高温30℃、32℃和34℃的保湿和干燥条件下,测定壳长为(4.43±0.76)mm和(10.16±0.43)mm稚贝的耐干露能力。结果表明:1)在相同湿度条件下,厚壳贻贝稚贝干露后存活率与干露时间呈显著负相关(P<0.05)。 1℃~8℃条件下,厚壳贻贝干露半致死时间长达45~91 h,而温度为30℃~34℃时,干露的半致死时间为17~23 h,低温时的耐干露能力是高温时的3倍以上。2)大规格稚贝的耐干露能力优于小规格稚贝。小规格厚壳贻贝4℃组的半致死时间为70~79 h,大规格4℃组的半致死时间为76~91 h。3)稚贝在保湿条件下的存活率明显高于干燥条件(P<0.05)。4)低温干露后,返水缓变温处理组的厚壳贻贝存活率显著高于急变温处理组,而高温干露后则情况相反。综上所述,以4℃保湿干露为厚壳贻贝稚贝的最佳运输条件,可保证24 h内90%以上的存活率。
厚壳贻贝;耐干露能力;高温耐受性;温度;湿度
厚壳贻贝(Mytiluscoruscus),俗称海红,隶属于软体动物门(Mollusca)、瓣鳃纲(Lanellibranchia)、 异柱目 (Anisomyaria)、贻贝科(Mytidea),在我国黄海、渤海及东海的离岸岛礁上较为常见。贻贝常以海水中的一些微生物及有机碎屑为食,以足丝附着在附着物上为生。厚壳贻贝是海产贝类中生长速度最快的种类之一,因此其也是我国较为常见的水产经济物种之一。近年来,厚壳贻贝的人工育苗及养殖技术已经获得突破,并在浙江嵊泗县形成贻贝养殖主产区。
干露时间是指水生生物脱离水体后暴露在空气中能存活的时间,又称“露空时间”。当水生生物处于干露状态下,细胞内外水分平衡因为生物体内水分逐渐蒸发而被打破,导致细胞失水萎缩。同时,水生生物在失水状态下不能直接从环境中获得充足的氧气并排出二氧化碳,因此会造成血液中pH失去恒定性而导致生物窒息性死亡[1]。已有研究文献报道了各种环境条件下双壳贝类的耐干露能力结果表明:干露温度、湿度以及干露时间对贝类存活率产生影响[2-6];太平洋牡蛎、墨西哥湾扇贝、虾夷扇贝在高温干燥条件下存活率明显下降[7-9];硬壳蛤存活率随着时间的增加而逐渐下降[10]。但关于厚壳贻贝稚贝耐干露相关研究至今未见报道。在厚壳贻贝养殖的苗种运输过程及收获环节中都会面临干露的问题,生产上由于缺乏对其稚贝耐干露能力的了解,在苗种长途运输中常出现大批死亡,造成巨大的经济损失,如何提高贻贝干露过程中的存活率,成了养殖者十分关心的问题。因此,本研究对厚壳贻贝稚贝生产运输过程中干露时间的长短掌控具有重要的理论指导意义。
1 材料与方法
1.1 材料
本实验样品取自浙江省嵊泗县枸杞贻贝养殖基地,共采集大、小两种规格的稚贝样品各5000个。实测两组壳长(精确至0.01 mm)分别为(4.43±0.76)mm、(10.16±0.43)mm。稚贝置于30 L水槽中暂养5 d,水温为(22±2)℃,与海区相近,海水盐度28。每天以适量角毛藻投喂稚贝,隔日吸污并换水1/2。
1.2 方法
1.2.1 低温条件下稚贝耐干露能力比较
从暂养的大、小两种规格稚贝中各随机选出1200个健康活泼个体,每一规格随机分成4组,每组300个,用湿纱布包裹,分别置于3 L保湿塑料盒中。在温度设定为1℃、2℃、4℃和8℃的恒温培养箱中分别放入大、小稚贝各一组恒温培育。在培育时间到6、12、18、24、30、36、42、48、60和72 h时,分别从各组中快速取出20个稚贝返水,其中10个稚贝分别置于1℃、2℃、4℃及8℃相应同温的海水中,自然回升至20℃室温,为温度缓变组。将另10个稚贝直接投入20℃的海水中,为温度急变组。24 h后观察并统计缓变与急变各组的死亡率与存活率。稚贝死亡的界定以解剖镜下观察双壳张开不能闭合为标准,即认定该稚贝已死亡。
1.2.2 高温条件下稚贝耐干露能力比较
试验方法同上,以小规格稚贝为材料,培养箱温度分别设定为30℃、32℃及34℃。
1.2.3 保湿和干燥条件下稚贝的存活率的比较
随机取出暂养的大规格稚贝700个,平均随机分成14组,每组50个。其中7组为保湿处理组,用湿润纱布包裹,另7组为干燥处理组,用干燥纱布包裹,处理后样品按组分别放入相应1 L聚乙烯塑料瓶中。在温度设定为0℃、5℃、10℃、15℃、20℃、25℃及30℃的7个恒温箱中分别各放入一组保湿、干燥处理的稚贝。在12、24、36和48 h时各瓶取20个,放入与培养箱同温海水,并自然回升至室温20℃静置24 h,计算各组稚贝存活率。
1.3 数据处理
稚贝存活率(%)=(实际存活数/取样数)×100%
2 结果与分析
2.1 低温条件下厚壳贻贝稚贝的耐干露能力
在1℃、2℃、4℃及8℃环境中,两种规格稚贝随时间延长的存活率变化如图1、图2所示。总体来看,稚贝在低温条件下的干露时间越长存活率越低。
图1-A为小规格稚贝干露后温度缓变处理组,干露24 h后,4个低温组的稚贝存活率仍在90%以上,且4个温度组之间的存活率无显著性差异(P>0.05);干露30 h后存活率开始显著降低;至36 h时,4组稚贝存活率仅为70.12%、77.02%、72.45%和72.72%,组间也开始出现了显著性差异(P<0.05);而在干露24 h后的稚贝的存活率与干露48 h后的稚贝存活率有极其显著的差异(P<0.01),1℃组跌至31.39%,2℃组为50.23%,8℃组40.70%,而4℃组最好,尚有64.86%存活率。此后1℃和8℃组存活率继续快速下跌,2℃组稍有下降,4℃组基本稳定;至72 h实验结束时,4℃组的存活率分别是1℃、2℃和8℃组的6倍、1.5倍和2倍。
图1-B是小规格稚贝干露后温度急变处理组存活率变化的结果,由图可见,在干露24 h内,在低温条件下的4个温度组,存活率不存在显著性差异(P>0.05),但存活率均已降至86%左右;干露36 h时,各组存活率下降显著,各温度组之间也出现显著差异(P<0.05),以4℃组最高,为75.75%;48 h时4℃组依然保持优势。此后1℃组存活率急剧下降,至72 h时,1℃组仅剩12.57%个体存活,4℃组仍有一半存活,2℃、8℃组的存活率介于1℃和4℃之间。
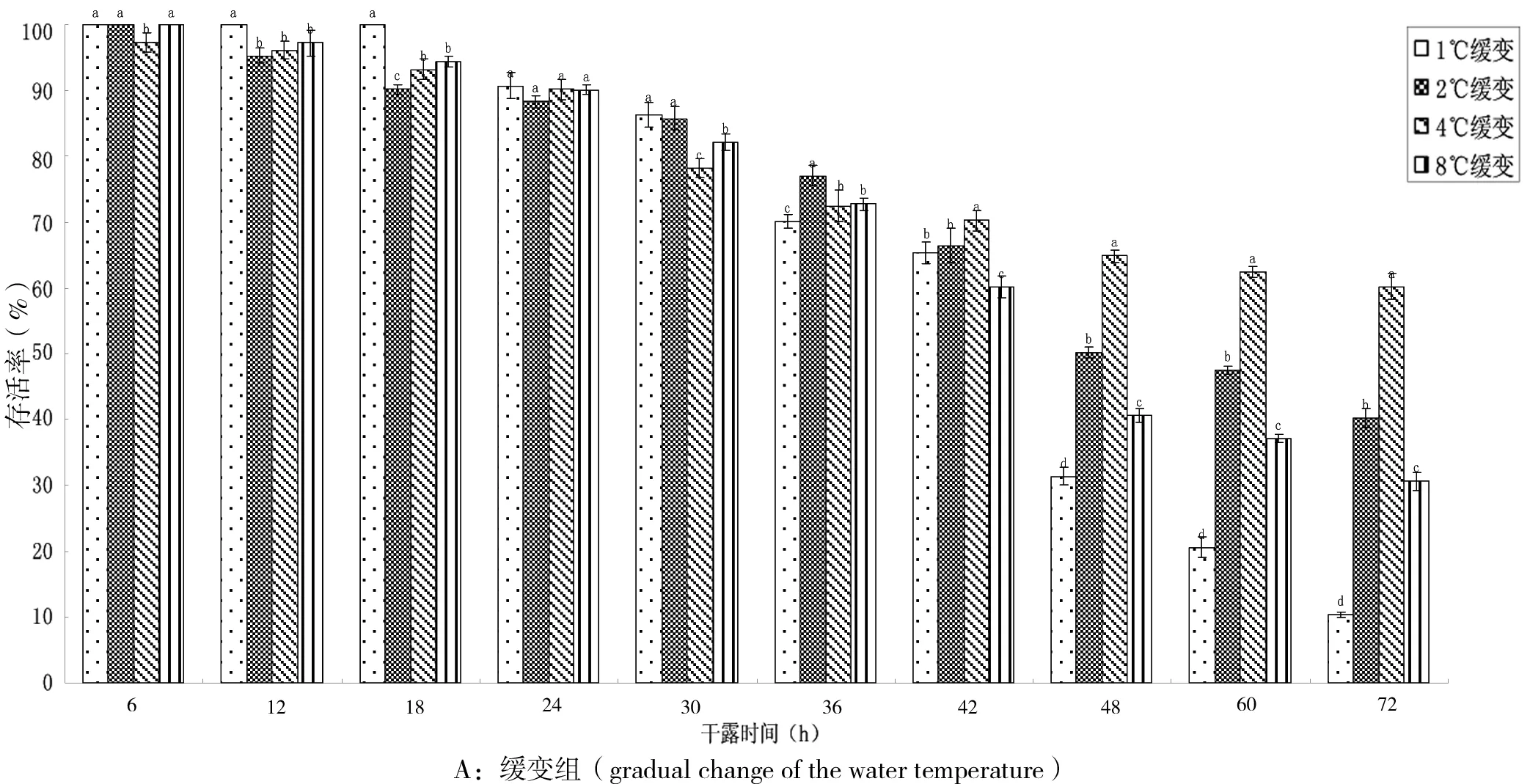
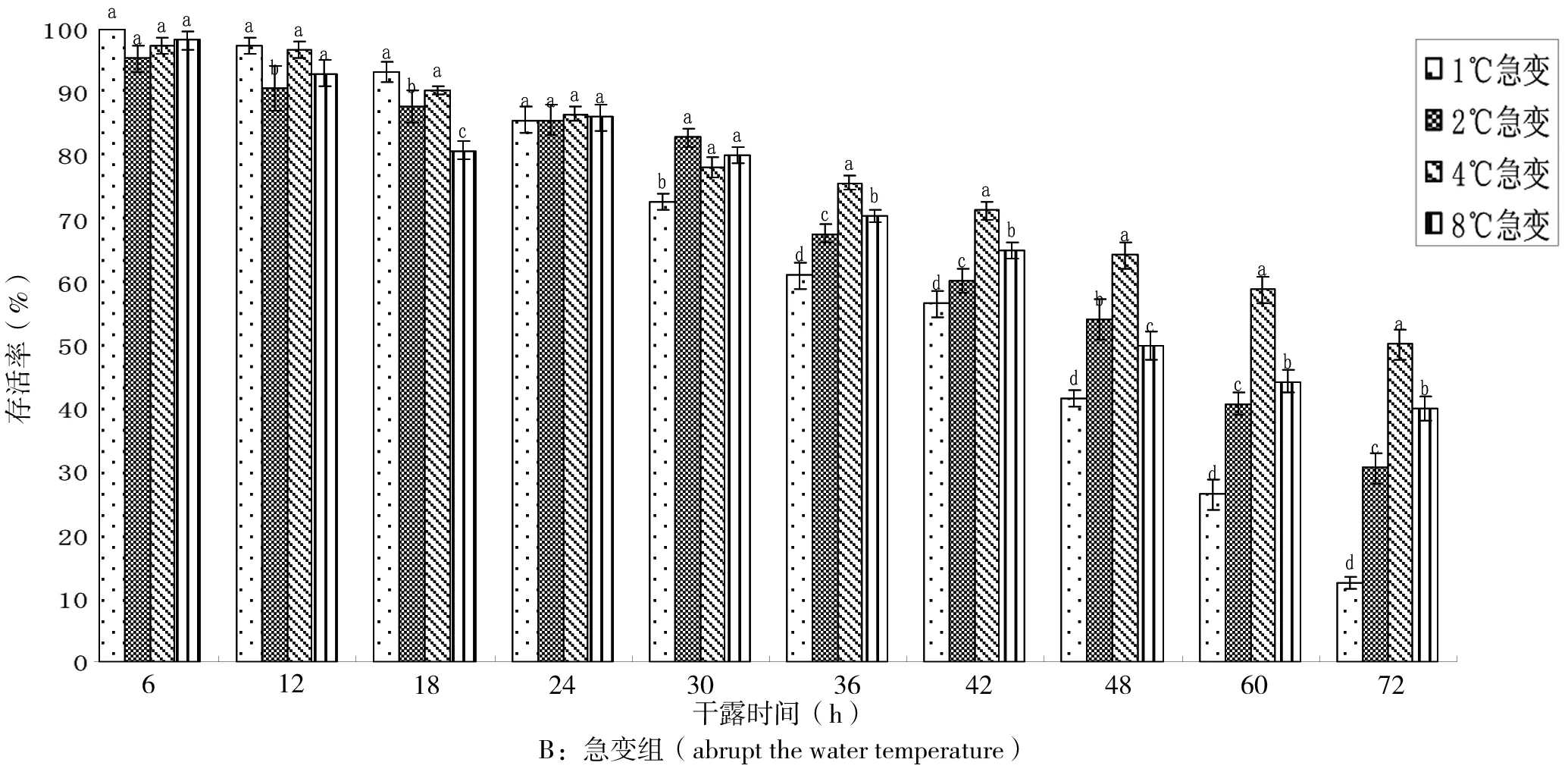
图1 低温条件下小规格稚贝在不同干露时间的存活率
从图2-A的大规格稚贝干露后缓变组存活率看,干露24 h 4个温度组稚贝存活率均为100%;30 h开始各组出现死亡,36 h时1℃和2℃组存活率显著低于4℃和8℃组(P<0.05),48 h时1℃组的存活率显著低于其他3组(P<0.05)仅为61.82%,其中4℃组存活率最高,为77.12%;至72 h实验结束时,除4℃组仍有60.23%存活率,其他3组均低于50%。
大规格稚贝干露后急变组存活率如图2-B所示。干露12 h内4个温度组的存活率均保持在100%;在24 h时,各组存活率维持在98.85%~90.13%;干露36 h后,1℃和2℃组存活率分别跌至75.02%和77.02%,显著低于4℃和8℃组(P<0.05);1℃组在48 h时存活率已近50%,此时4℃和8℃组仍有71%左右;至实验结束72 h时,1℃组存活率最低,为30.52%,其他3组均在50%左右,其中以4℃组最佳。
2.2 高温条件下稚贝的耐干露能力比较
稚贝在30℃、32℃、34℃高温条件下不同干露时间的存活率如图3所示。结果表明,稚贝在高温条件下耐干露能力明显不及低温时耐干露能力,随干露时间的延长,稚贝存活率迅速降低,干露42 h后返水存活率为0%。


图2 低温条件下大规格稚贝在不同干露时间的存活率
图3-A结果显示,3个高温组在干露6 h后温度缓变返水条件下,存活率为92.21%~100%,12 h时34℃组已降为74.25%,3组差异显著(P<0.05);18 h时32℃组存活率最高,为82.21%;24 h时34℃组急降为10.12%,极显著低于30℃和 32℃两组(P<0.01);干露30 h 3组样品存活率均跌至20%以下;干露36 h时30℃、32℃组尚有少量存活,但34℃组已经全部死亡。
图3-B可见,3个高温组在干露6 h后急变温度下,稚贝存活率无显著差异(P>0.05);12 h时30℃组的存活率仍高达90.01%,但34℃组已降至60.13%,组间存活率差异显著(P<0.05);18 h时34℃组已跌至41.22%,32℃组为63.21%,而30℃组依然维持在87.22%;干露24 h后,30℃组尚有60%的存活率,而此时32℃和34℃两组稚贝的存活率急剧下降至20%左右;30 h后,3组的存活率均已低于10%;36 h时, 30℃和32℃组存活率仅为5.08%和6.38%,34℃组稚贝已全部死亡。
2.3 稚贝在干燥与保湿条件下干露能力比较
稚贝在干燥与保湿条件下耐干露能力见图4和5。在保湿环境中,24 h内除30℃组存活率已低于50%,其余6个温度组全部存活;干露48 h后,5℃~25℃实验组稚贝存活率均维持在95%以上,显著高于0℃和30℃实验组(P<0.05)。干燥环境中,稚贝干露后存活率明显不如保湿条件下存活率,干露36 h时,0℃和30℃两组稚贝已全部死亡;48 h时,5℃组存活率最高,达到38.93%,而其他实验组存活率均低于10%。干露36 h以内,保湿条件与干燥条件下,稚贝存活率差异不显著(P>0.05),而超过36 h的两种条件下存活率差异显著(P<0.05)。

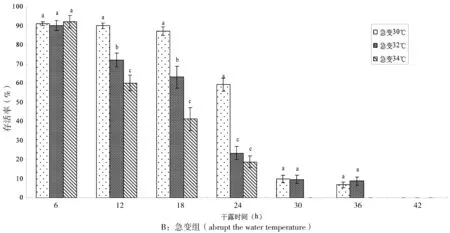
图3 高温条件下小规格稚贝在不同干露时间的存活率
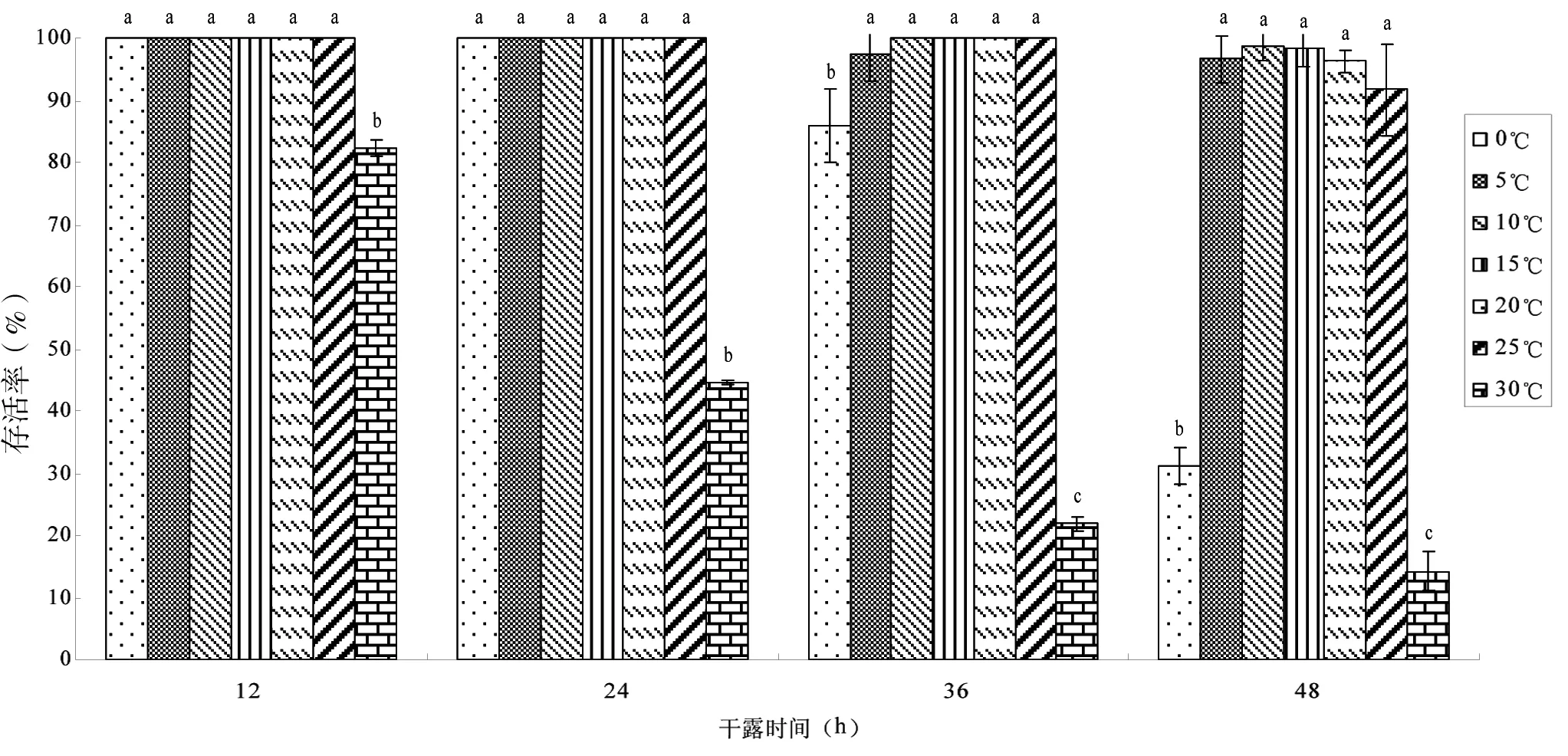
图4 稚贝在保湿条件下干露不同时间的存活率
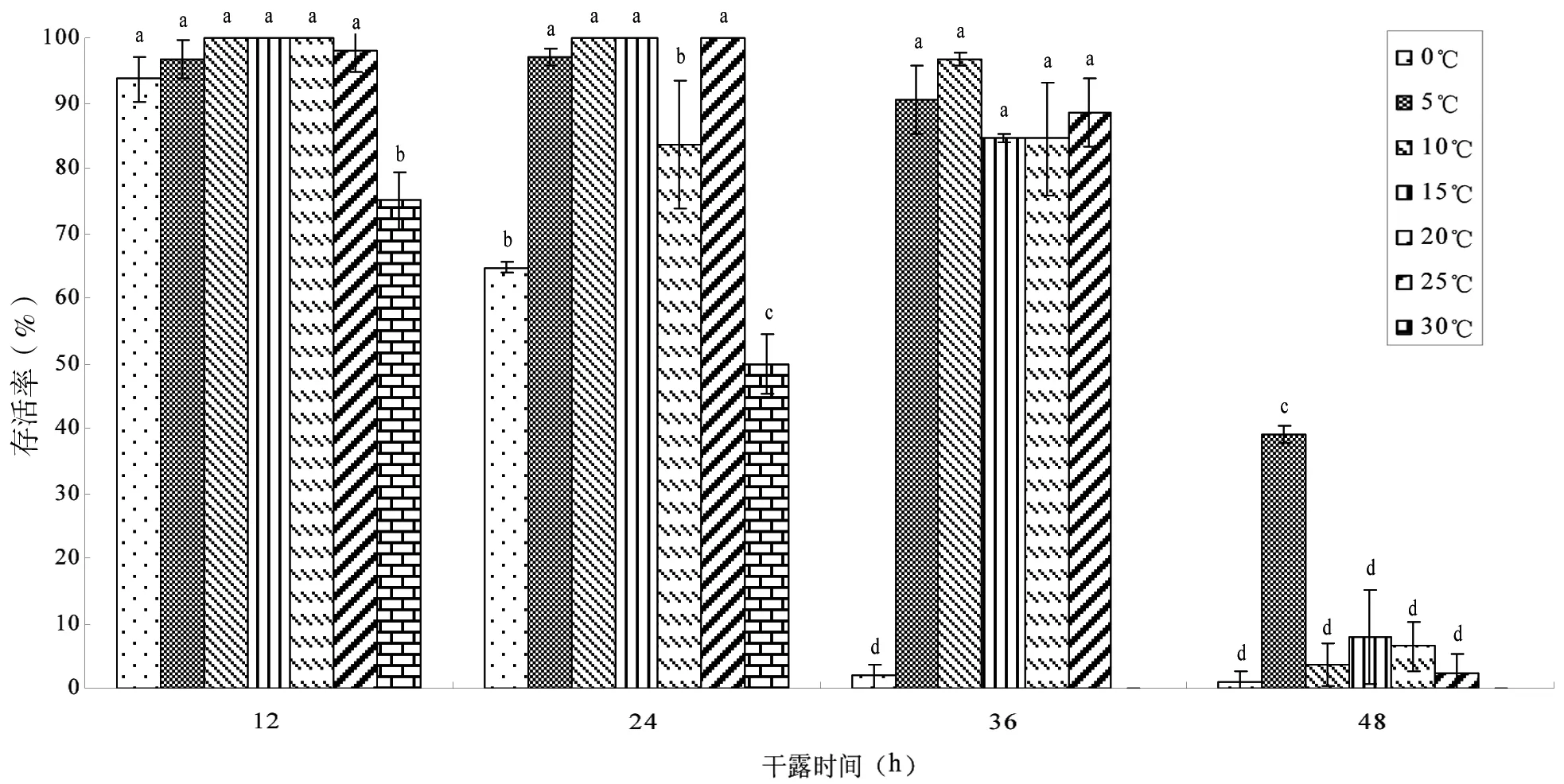
图5 稚贝在干燥条件下干露不同时间的存活率
2.4 干露半致死时间
由上述图1~图5可得厚壳贻贝稚贝干露时间与存活率的回归方程,并进一步计算得到各温度下的干露半致死时间见表1。
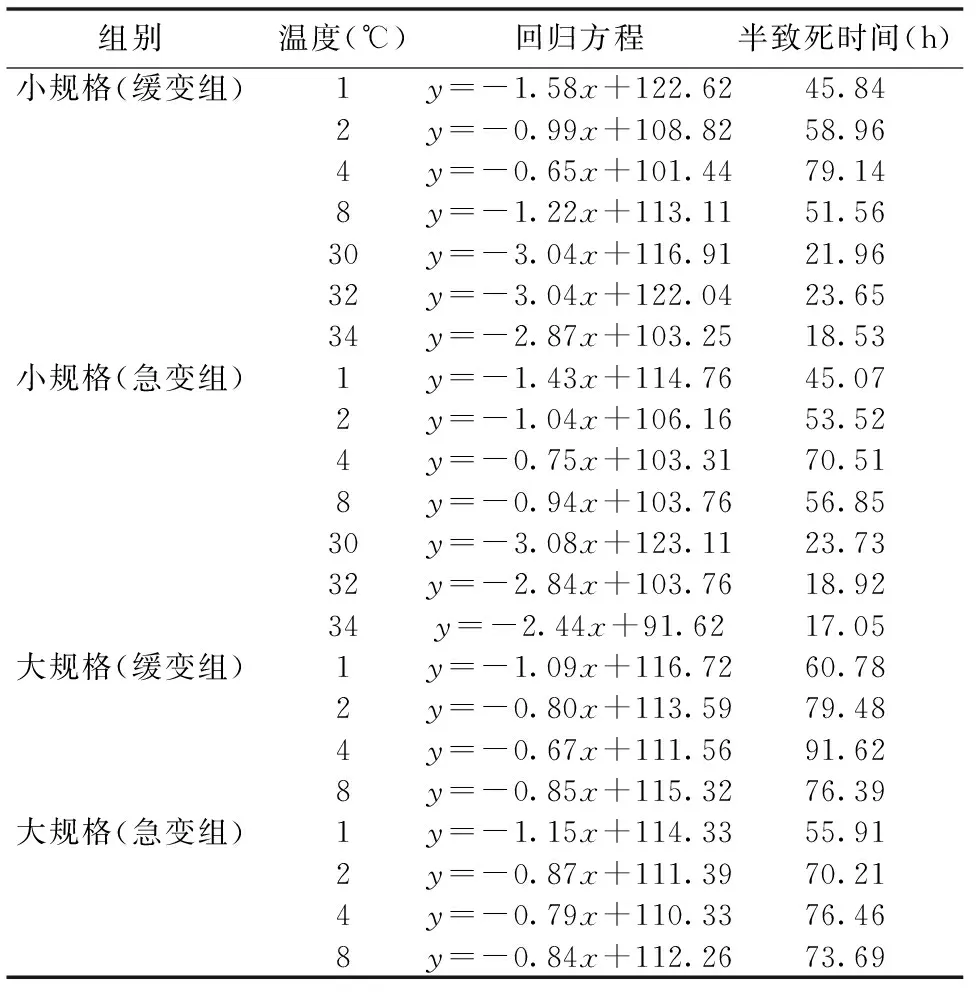
表1 不同干露条件下厚壳贻贝稚贝半致死时间
由表1可见,在1℃~4℃低温干露下,厚壳贻贝稚贝的半致死时间随着温度的升高而增加,小规格稚贝4℃组的半致死时间为70~79 h,大规格稚贝4℃组的半致死时间为76~91 h,且8℃组的半致死时间不及4℃组;而高温组半致死时间随着温度的升高而缩短,最长半致死时间仅为23~24 h;同时可知,大规格稚贝耐干露能力优于小规格稚贝(P<0.05)。且无论低温干露还是高温干露,返水时温度缓变组均优于温度急变组。
3 讨论
在干露后,双壳贝类的外套膜会处于闭合状态,而为保证其细胞呼吸时内外气体交换和新陈代谢的正常进行,其软体部仍保留一定的水分。贝类的露空时间由其保水能力及其对低含水量的耐受性来决定[10]。于瑞海等指出:扇贝在其他条件完全相同时,因两壳不能完全闭合,干露时间越长,软体部失去水分的就越多,干露后的成活率就越低,这与牡蛎和文蛤等双壳类不同[2]。稚贝软体部组织水分过度蒸发的情况在湿润状态下,会明显好转,进而显著提高其耐干露能力[11]。但是不同双壳贝类的结构、生理和生态特点各有不同,因此它们的耐干露能力也存在一定的差异。厚壳贻贝的双壳可以完全闭合,从而能够长时间地将水分锁在外套膜中,因此相比于扇贝等双壳不能完全闭合的贝类,厚壳贻贝具有更强的耐干露能力。有研究表明常温下,相较于施氏獭蛤稚贝的9 d耐干露时间[12],墨西哥扇贝稚贝离水后的干露时间不超过15 min,其耐干露能力较差[13]。硬壳蛤稚贝[10]和太平洋牡蛎稚贝[7]的干露时间分别为100 h和50 h左右。本研究表明,厚壳贻贝稚贝在1℃~8℃低温保湿条件下,能够维持50%以上存活率的干露时间为 45~91 h,而在30℃~34℃高温保湿条件下,能够维持50%以上存活率的干露时间为17~23 h。由此可见,厚壳贻贝稚贝在低温条件下的耐干露能力是高温时的3倍以上,这与厚壳贻贝属于冷温性种类,自然分布的最南端到福建平潭,且夏天海水温度较长时间超过31℃就会脱苗死亡的事实相符合[14]。本实验也证实,无论低温干露还是高温干露处理,稚贝的存活率与干露时间都呈显著负相关,这是因为随着干露时间增加,细胞也会加速失水,最终导致其窒息性死亡。有研究表明,在干露条件下,牡蛎为发挥调节缺氧适应的作用,会表达过量的HIF-1,同时也会降低磷酸葡糖异构酶和磷酸果糖激酶等有氧呼吸酶的表达[15],这说明,降低代谢速率可能是贝类在干露条件下为维持生存,减少能量消耗的一条途径[16]。对于同种贝类,由于其个体成长环境、个体大小等原因,耐干露能力也会有所不同。图4、图5以及表1显示,厚壳贻贝稚贝规格大小也直接关系到其耐干露能力,小规格稚贝耐干露能力显著劣于大规格稚贝。该结论也与于瑞海等对不同发育期太平洋牡蛎干露成活率的研究结果相近[7],说明稍大规格的稚贝拥有更加完善的生理代谢调节机能。
温度是影响水生变温动物生长和生存的重要环境因子之一。厚壳贻贝生长适宜水温为4℃~31℃,最适水温为15℃~27℃,最适盐度是24.6~35.8[14]。因此,本研究观察到低温1℃~8 ℃和高温30℃~34℃对稚贝耐干露能力具有显著影响。于瑞海等[2]研究发现,湿度相同时,海湾扇贝在低温条件下的存活率明显高于高温组;曹井志等[17]通过探索厚壳贻贝低温无水保活技术,发现厚壳贻贝成贝能够在-1.5℃下存活;也有研究认为厚壳贻贝能够在35℃条件下存活,但对高温的耐受力极差[18]。高温条件下稚贝的呼吸代谢强度较大,能量消耗及细胞水分散失都会加快,导致稚贝死亡速率加快。因此,为利于稚贝存活,使其被迫进行厌氧代谢并减缓代谢速率,可以保持一定的低温,并保持一定的湿度。本研究结果表明,以4℃保湿干露为厚壳贻贝稚贝的最佳运输条件,在干露时间控制在24 h内即可以保证稚贝90%以上的存活率。
干露后的返水温度在同类研究中容易被忽视,本研究发现低温组返水时缓变温处理组存活率显著高于急变温处理组,而高温组则相反。这是由于稚贝由低温条件突然进入20℃常温水中,使低温干露条件下形成的低能耗代谢与无氧呼吸模式来不及转变,返水后细胞开始有氧代谢,体内能量消耗瞬间增大,导致代谢失衡,造成大量死亡;而高温32℃~34℃下,稚贝已经处于极限温度,返水缓变温度反而延长了高温对其组织细胞及酶系统的损害时间,故出现缓变组存活率低于急变组的现象。厚壳贻贝这种较强的耐干露能力也为其苗种的长距离运输提供了方便,生产上经常采用厢式冷藏车干露保湿运输,能保证较高的苗种运输存活率,若能同时采取下海前温度适应过渡,会进一步提高苗种的下海成活率。
[1]杨 凤,谭文明,闫喜武,等. 干露及淡水浸泡对菲律宾蛤仔稚贝生长和存活的影响[J]. 水产科学, 2012, 31(3):143-146.
[2]于瑞海,辛 荣,赵 强,等. 海湾扇贝不同发育阶段耐干露的研究[J]. 海洋科学, 2007, 31(6):6-9.
[3]曹 琛,匡少华,姜 超,等. 干露及盐度变化对毛蚶苗的影响试验[J]. 河北渔业, 2007(5):38, 51.
[4]马贵范,杨 凤,郭文学,等. 中国蛤蜊和四角蛤蜊对干露和淡水浸泡的耐受性比较[J]. 大连海洋大学学报, 2013, 28(1):44-48.
[5]张 涛,杨红生,刘保忠,等. 环境因子对硬壳蛤Mercenariamercenaria稚贝成活率和生长率的影响[J]. 海洋与湖沼, 2003, 34(2):142-149.
[6]宁军号,张向向,常亚青,等. 几种生态因子对成体偏顶蛤存活的影响[J]. 大连海洋大学学报, 2015, 30(3):285-290.
[7]于瑞海,王昭萍,孔令锋,等. 不同发育期的太平洋牡蛎在不同干露状态下的成活率研究[J]. 中国海洋大学学报(自然科学版), 2006, 36(4):617-620.
[8]刘志刚,王 辉,栗志民,等. 墨西哥湾扇贝高起始致死温度的研究[J]. 中国水产科学, 2007, 14(5):778-785.
[9]郝振林,唐雪娇,丁 君,等. 不同高温水平对虾夷扇贝存活率、耗氧率和体腔液免疫酶活力的影响[J]. 生态学杂志, 2014, 33(6):1580-1586.
[10]李忠泓,王国栋. 硬壳蛤稚贝对淡水浸泡、干露和低温的耐受能力[J]. 水产科学, 2004, 23(6):14-16.
[11]蔡英亚,黄翔鹄,吴洞科. 红树蚬的生态观察[J]. 热带海洋, 1995, 14(1):94-98.
[12]刘 超,郭景兰,彭张明,等. 施氏獭蛤稚贝对高温和干露的耐受性研究[J]. 水产科学, 2015, 34(3):169-173.
[13]杨红生,张 涛,王 萍,等.温度对墨西哥湾扇贝耗氧率及排泄率的影响[J]. 海洋学报, 1998, 20(4):91-96.
[14]叶 鹏,蔡厚才,庄定根,等. 南麂海区野生贝类增养殖种类初步筛选[J]. 渔业现代化, 2006, 33(4):26-28.
[15]张琳琳. 长牡蛎防御相关基因的进化和表达调控分析[D].青岛:中国科学院研究生院(海洋研究所), 2012.
[16]张继红,方建光,梁翻鹏. 低温对栉孔扇贝能量收支的影响[J]. 中国水产科学, 2002, 9(1):48-51.
[17]曹井志,徐 若,包建强. 厚壳贻贝低温无水保活技术[J]. 安徽农业科学, 2008, 36(10):4248-4249.
[18]黄振华,柯爱英,王瑶华,等. 生态因子对厚壳贻贝眼点幼虫生长和存活的影响[J]. 水产养殖, 2015, 36(1):41-44.
Study on enduring exposure and desiccation ability ofMytiluscoruscusin different temperature and humidity conditions
SUN Yu-yang, TONG Qiao-qiong, WANG Wen, ZHAN Ping-ping, CHENG Liang, XU Shan-liang, WANG Chun-lin
(School of Marine Science, Ningbo University, Ningbo 315211, China)
In order to understand the tolerance ofMytiluscoruscusto dry exposure in different temperature and humidity or dry conditions, we measured the juvenile with the shell length of (4.43±0.76)mm and (10.16±0.43)mm. The experiments were performed at low temperature (1℃,2℃, 4℃ and 8℃) and high temperature (30℃, 32℃ and 34℃) with the condition of being dry and moist, respectively. The results showed that the survive rate of juveniles after exposure and desiccation had a negative correlation with the time of the tolerance under the same humidity (P<0.05). And the semilethal time was 45-91 h at 1-8℃, while 17-23 h at 30-34℃. The endurance of juveniles to exposure and desiccation under low temperature was more than triple under high temperature. The ability to survival of large size ofM.coruscuswas stronger than that of the small size one. The semilethal time of small size juveniles was 70-79 h, while the semilethal time of lager size was 76-91 h at 4℃. The survival rate ofM.coruscusunder moisturizing conditions is higher than that in drying conditions (P<0.05). The survival rate about the group of rewet with slow change in temperature was higher than the quick change in temperature group after air expose in low temperature, but the result after high temperature exposure was the opposite. Research results that the optimal transportation condition ofMytiluscoruscuswas air exposured under moisture condition at 4℃. In this way, the survival rate could be guaranteed more than 90% in 24 h.
Mytiluscoruscus; ability of enduring exposure and desiccation; high temperature tolerance; humidity; temperature
2016-08-02;
2016-10-31
国家星火计划项目(2015GA701003);宁波市科技专项(2015C10006);浙江省贻贝产业科技特派员服务团队项目
孙煜阳,海洋资源与环境专业,E-mail:562871097@qq.com
徐善良,教授,硕士生导师,从事海洋生物研究,E-mail:xushanliang@nbu.edu.cn
Q495
A
2095-1736(2017)04-0042-05
doi∶10.3969/j.issn.2095-1736.2017.04.042
猜你喜欢
检察风云(2024年10期)2024-05-24 17:38:09
建材发展导向(2021年11期)2021-07-28 06:57:22
烹调知识(2020年10期)2020-10-20 05:47:44
当代水产(2020年10期)2020-03-17 07:02:48
当代水产(2019年8期)2019-10-12 08:57:26
大连海洋大学学报(2017年3期)2017-06-19 18:56:11
河北渔业(2016年5期)2016-09-08 10:31:08
水产养殖(2016年2期)2016-03-28 11:47:52
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:42
河北渔业(2015年6期)2015-06-24 11:19:13
