
人为干扰对油松天然林空间分布格局的影响1)
2017-08-07 06:22刘宪钊王金龙李卫珍陆元昌国红邢海涛
东北林业大学学报 2017年3期
刘宪钊 王金龙 李卫珍 陆元昌 国红 邢海涛
(中国林业科学研究院资源信息研究所,北京,100091) (山西省黑茶山国有林管理局) (中国林业科学研究院资源信息研究所)
人为干扰对油松天然林空间分布格局的影响1)
刘宪钊 王金龙 李卫珍 陆元昌 国红 邢海涛
(中国林业科学研究院资源信息研究所,北京,100091) (山西省黑茶山国有林管理局) (中国林业科学研究院资源信息研究所)
以山西黑茶山3种不同干扰强度的油松天然林为研究对象,采用g(r)函数点格局方法,分析不同干扰强度下,油松天然林径级结构和多样性差异,以及优势种群油松不同生长阶段个体(幼树、小树、中树和大树) 的空间分布点格局。结果表明:油松径级结构呈单峰型,中径级的林木个体数较多。辛普森多样性和香农-维纳多样性随干扰强度的增加而逐渐减少;0~1 m尺度上,不同生长阶段个体呈聚集分布,随着种群发育(幼树-小树-中树-大树) ,油松天然林的空间格局趋于随机分布,这体现了种群的一种生存策略或适应机制。
黑茶山;油松;树种多样性;点格局
Heicha Mountain;Pinustabulaeformis; Tree species diversity; Point pattern
群落组成与空间格局是分析植物种群特征、种群间相互作用及种群与环境关系的重要基础[1-2],为研究物种共存机制提供了重要信息[3-4]。种群的空间格局对其生长、繁殖、死亡、再生、资源利用及其干扰等有显著的影响[5]。植物种群的空间格局在不同发育阶段、不同生境条件下会发生变化,当种群受到不同程度的外界干扰时,由于生境变化程度和不同种群自然恢复存在差异性,不同生物学特征的种群所具有的空间分布格局也存在差异[6]。种群的空间格局与其所处的空间尺度密切相关[7],在某些尺度上可能是聚集性分布,而在另外一些尺度上则可能变成随机分布或均匀分布[8]。常规方法仅能解决单一尺度下的空间分布格局,点格局分析方法(如Ripley’s K函数)以植物分布的空间坐标为基础,可以分析多尺度下的种群格局和空间关联分析[9-10],但其具有累积性,会混淆大尺度与小尺度的效应[11],g(r)函数是在K函数的基础上衍生而来,是K函数的概率密度函数,能有效剔除K函数小尺度上的累积效应而得到广泛应用[12]。
油松(Pinustabulaeformis)为我国特有树种,其耐干旱、耐瘠薄、自然分布广,是华北山区主要树种之一,也是温带针叶林中分布最广的植物种群[13]。山西黑茶山自然保护区是油松天然林的典型分布区之一,对维持华北北部山区生态系统稳定性具有重要意义[14],由于历史及社会原因,该地区天然林均不同程度地受到人为活动的干扰。目前山西省油松林的研究主要以中部太岳山林区为主[15],山西省北部黑茶山油松边缘区天然林种群组成和分布格局的研究尚属空白。本文运用点格局分析方法探讨了不同干扰程度、不同发育阶段的油松个体,在不同空间尺度下的分布规律,以期为油松天然林的保护与可持续利用提供理论依据。
1 研究区概况
研究区位于黑茶山林区南部的南阳山林场。地理坐标为东经111°15′~111°36′,北纬37°53′~38°11′。南阳山林场气候属暖温带季风型大陆性气候,四季明显,冬季少雪干旱,春季多风干燥,夏季雨量集中的特点。气候温和干燥,年平均气温约7.1 ℃,日均温≥10 ℃的年积温2 351 ℃左右,年降水量600 mm,多集中在7—9月份,年日照时间2 800 h,无霜期128 d左右。主要气候灾害有干旱、霜害等。全区共有野生种子植物81科332属687种,其中:裸子植物3科7属9种,被子植物78科325属678种(双子叶植物69科269属576种,单子叶植物9科56属102种)[14]。
2 研究方法
2.1 野外数据采集
在全面踏查的基础上,2015年7月在黑茶山林区南阳山林场选取3种不同干扰强度的油松天然林分别建立40 m×40 m方形样地各1块,调查时,将样地分为64个5 m×5 m的小样方进行调查,调查胸径(D)大于等于1 cm的所有木本植物,挂牌并定位,记录树种名称、胸径、树高、枝下高、冠幅等,灌草层记录各物种的高度、盖度、多度。
2.2 点格局分析和数据处理
由于油松天然林实测年龄较为困难,本研究采用"空间代替时间"的伪时间序列方法来研究油松天然林种群动态[16]。根据研究区油松林径级分布和林场关于油松天然林的档案材料,按照胸径(D)将油松划分为径级I(幼树,0≤D<4 cm)、径级II(小树,4 cm≤D<12 cm)、径级III(中树,12 cm≤D<20 cm)、径级IV(大树,D≥20 cm)共4个生长阶段,分析油松的空间格局。
点格局分析法是将每个个体视为二维空间的一个点,以全部个体组成的二维点图为基础进行格局分析。采用点格局分析方法中的单变量的g(r)函数分析不同生长阶段油松种群空间格局[17]。其中,Ripley’s K函数为:

(1)
式中,A为样方面积,λ为模型估计参数,指样方内物种个体密度,Wij为边界效应修正,dij指两随机点间的距离,I为指示函数。当dij≤r时,I=1.0,当dij>r时,I=0。
g(r)是从K函数衍生出来的,主要以环代替K函数中的圆,计算过程没有累积效应。即:
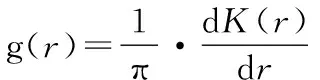
(2)
当g(r)=1.0时,个体分布显示为完全空间随机分布,g(r)>1.0时为聚集分布,g(r)<1.0时为均匀分布。
用Monte-Carlo方法循环99次,产生置信度为99%的包迹线,以检验两个树种的分布格局。数据分析过程使用软件R 3.2.2完成[18]。
3 结果与分析
3.1 不同干扰强度下油松天然林径级结构
对3种不同干扰强度的油松天然林调查样地林木株数进行统计分析,强、中、弱3种干扰程度下油松天然林密度分别为1 665、2 401、3 376株/hm2。由表1可知,强度干扰下,油松天然林从小到大4个径级的密度比约为1∶62∶32∶4;中度干扰和弱度干扰情况下,该比例分别变为3∶73∶21∶3和1∶83∶13∶3。从目前径级结构来看,3种油松天然林分,虽然均表现为近正态分布的稳定型种群,但弱度干扰的油松天然林幼树和中树、大树资源结构远不及中度和强度干扰下的油松天然林分,林分密度过大抑制林下更新的发育,最终影响油松天然林后续的更新交替。

表1 山西省黑茶山不同干扰强度油松天然林径级分布
3.2 不同干扰强度下油松天然林多样性分析
由表2可知,辛普森多样性和香农-维纳多样性随干扰强度的增加而逐渐减少,即使中度干扰条件下的油松天然林内,其它树种的种类少于强度干扰油松天然林。中度干扰油松林内树种均匀度最高,其次为弱度干扰和强度干扰。树种丰富度以弱度干扰油松林最高(6种)、其次为强度干扰(5种),中度干扰油松林树种丰富度最低(4种)。树种优势度以强度干扰最高,并随干扰程度的增加而减低。

表2 不同干扰强度油松天然林树种多样性
3.3 不同干扰强度下油松天然林空间分布格局
在0~1 m尺度上,幼树、小树、中树和大树在强度干扰下均表现为从聚集分布,1~10 m为随机分布,在10 m的尺度上则呈现均匀分布趋势。
在中度干扰下,幼树、小树、中树和大树表现为在0~1 m尺度下,聚体趋势先降后升,1~10 m从聚集分布转变为随机分布。
在弱度干扰下,幼树、小树、中树和大树也呈现与强度干扰和中度干扰同样的分布趋势,且在0~1 m尺度呈弱的聚集分布。

图2 不同干扰强度油松种群不同生长阶段个体的点格局分布
4 结论与讨论
以径级分布为特征的水平结构是森林群落稳定和生长发育状况的重要指标[19],由于在实际测量中树木的年龄结构难以获得,而同一环境下油松的胸高直径与年龄、生活史存在明显的正相关关系[13,15],油松林直径分布特征常被用来描述其种群和结构的稳定性。本研究中,不同干扰程度的油松天然林在径级II和径级III的个体数量最多,属于稳定型种群。但弱度干扰的油松天然林幼树资源远不及中度和强度干扰下的油松林,林分密度过大会抑制林下更新的发育,最终影响油松天然林后续的更新交替,在后续的抚育经营中需特别关注。
干扰通过改变植物群落内的环境条件、物种组成和多样性等,进而改变植物群落的结构和功能,影响其演替进程甚至演替方向[20]。目前,Connell[21]提出的"中度干扰假说"(中等程度的干扰有利于群落多样性达到高水平)得到广泛的认可。本研究中油松天然林树种多样性表现出随干扰强度的增加而减小的现象,而中度干扰只是在物种的均匀度上高于弱度和强度干扰。油松天然林的封育保护一定程度上维持了群落的多样性,但无法实现树种多样性的提高,科学合理的经营性干扰可能成为提高该地区油松天然林树种多样性的一个主要方式。
一般认为,小尺度上的种群分布多为种子的传播限制及物种自身生物学特性作用的结果,大尺度分布格局则更多受到环境异质性的影响。本研究中,油松的空间分布受到种子扩散限制和环境异质性的影响,在样地0~1 m的尺度中呈现聚集分布,而在1~10 m的尺度上表现为随机分布。种群以聚集分布形式占有和利用环境资源,有利于维持种群自身稳定,油松为阳性树种,种子在成熟扩散过程中,部分种子因物理作用随机分散母树周围,当雨季来临,种子因水淹时间过长而失去萌发能力,影响其扩散。
植物的空间分布格局可能受到自然资源(光、水分、土壤养分)的斑块性分布影响,导致物种容易在一些资源丰富的地块呈现出聚集分布,油松和其它阔叶树种属伴生关系,能够长期共存。但长期的封育造成油松天然林密度过大,个体间的挤压和竞争影响其生长发育,同时几乎郁闭的林冠层也阻碍了其它树种的落种、萌发和更新生长。为了提高该区油松天然林群落的稳定性、多样性,促进林分健康发育,有必要开展积极有效的抚育经营活动。
[1] HE F, DUNCAN R P. Density-dependent effects on tree survival in an old-growth Douglas fir forest[J]. Journal of Ecology,2000,88(4):676-688.
[2] JOHN R, DALLING J W, HARMS K E, et al. Soil nutrients influence spatial distributions of tropical tree species[J]. Proceedings of the National Academy of Sciences,2007,104(3):864-869.
[3] LOREAU M, NAEEM S, INCHAUSTI P, et al. Biodiversity and ecosystem functioning: current knowledge and future challenges[J]. Science,2001,294:804-808.DOI:10.1126/science.1064088.
[4] TILMAN D, REICH P B, KNOPS J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment[J]. Nature,2006,441,629-632.DOI:10.1038/nature04742.
[5] HUBBELL S P. The unified neutral theory of biodiversity and biogeography[M]. Princeton: Princeton University Press,2001.
[6] 杨梅,林思祖,王鸿.甜槠群落主要种群空间分布格局对人为干扰的响应[J].中南林学院学报,2005,25(5):33-35.
[7] WIEGAND T, MOLONEY K A. Rings, circles, and null-models for point pattern analysis in ecology[J]. Oikos,2004,104(2):209-229.
[8] STOYAN D, PENTTINEN A. Recent applications of point process methods in forestry statistics[J]. Statistical Science,2000,15(1):61-68.
[9] RIPLEY B D. The second-order analysis of stationary point processes[J]. Journal of Applied Probability,1976,13(2):255-266.
[10] DIGGLE P J. Statistical analysis of spatial point patterns[M]. 2nd ed. London:Arnold,2003.
[11] CONDIT R, ASHTON P, BUNYAVEJCHEWIN S, et al. The importance of demographic niches to tree diversity[J]. Science,2006,313:98-101.DOI:10.1126/science.1124712.
[12] ILLIAN J, PENTTINEN A, STOYAN H, et al. Statistical analysis and modelling of spatial point patterns[M]. West Sussex: Wiley,2008.
[13] 张赞,赵秀海,张春雨.华北油松林油松种群的空间分布格局[J].东北林业大学学报,2009,37(11):43-46.
[14] 郝晓鹏,上官铁梁.山西黑茶山自然保护区种子植物区系地理成分分析[J].山西大学学报(自然科学版)2009,32(S1):123-126.
[15] 黄三祥,张赞,赵秀海.山西太岳山油松种群的空间分布格局[J].福建林学院学报,2009,29(3):269-273.
[16] 洪伟,柳江,吴承祯.红锥种群结构和空间分布格局的研究[J].林业科学,2001,37(1):6-10.
[17] 张俊艳,成克武,臧润国.海南岛热带天然针叶林主要树种的空间格局及关联性[J].生物多样性,2014,22(2):129-140.
[18] 吴喜之.复杂数据统计学方法-基于R的应用[M].3版.北京:中国人民大学出版社,2015.
[19] 宾粤,叶万辉,曹洪麟,等.鼎湖山南亚热带常绿阔叶林20公顷样地幼苗的分布[J].生物多样性,2011,19(2):127-133.
[20] 毛志宏,朱教君.干扰对植物群落物种组成及多样性的影响[J].生态学报,2006,26(8):2695-2699.
[21] Connell J H. Diversity in tropical rain forest and coral reefs[J]. Science,1978,199:1302-1310.
1)林业科技推广示范项目([2014]07号);北京市自然科学基金项目(8152033)。
刘宪钊,男,1981年11月生,中国林业科学研究院资源信息所,副研究员。E-mail:lxz9179@caf.ac.cn。
2016年6月13日。
S718.54
责任编辑:王广建。
Spatial Distributions ofPinustabulaeformisNature Forest under Human-caused Disturbance//Liu Xianzhao(Institute of Forest Resource Information Techniques, Chinese Academy of Forestry, Beijing 100091, P. R. China); Wang Jinlong, Li Weizhen(Heicha Mountain National Forest Authority); Lu Yuanchang, Guo Hong, Xing Haitao(Institute of Forest Resource Information Techniques, Chinese Academy of Forestry)//Journal of Northeast Forestry University,2017,45(3):13-16.
With the investigation data from three types ofPinustabulaeformisnature forest with different disturbing degrees in Heicha Mountain, Shanxi Province, we studied the diameter structure and community diversity of these types forests. We discussed the spatial pattern ofP.tabulaeformisat different growth stages by g(r) function. The diameter structure was presented a unimodal distribution and middle age individuals accounted more. The Simpson and Shannon-Wiener diversity index were decreased with the increasing of the disturbing degree. All trees had a clustered distribution across a range of scales (0-1 m). Varied with growth stage (saplings-small trees-middle tree-big trees), the spatial pattern tended to be random, which showed the survival strategy and suitability in certain degree.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
南方农业·下旬(2022年4期)2022-05-24
山西林业(2021年2期)2021-07-21
中国水土保持科学(2019年6期)2019-04-26
作文大王·低年级(2019年3期)2019-04-19
心声歌刊(2018年6期)2019-01-29
红土地(2018年7期)2018-09-26
现代园艺(2017年22期)2018-01-19
西北植物学报(2015年9期)2015-07-05
