
冬虫夏草线状菌体制剂对蝠蛾幼虫感染的研究
2017-08-02 01:39贺宗毅张德利邢康康陈仕江
环境昆虫学报 2017年3期
贺宗毅,张德利,李 黎,邢康康,陈仕江*
冬虫夏草线状菌体制剂对蝠蛾幼虫感染的研究
贺宗毅1,2,3,4,张德利1,2,3,4,李 黎1,4,邢康康1,2,3,4,陈仕江1,2,3,4*
(1.重庆市中药研究院,重庆 400065;2.重庆市中药资源学重点实验室,重庆 400065;3.中国中医科学院,中药资源中心重庆分中心,重庆 400065;4.甘孜州康定贡嘎中华虫草产业有限责任公司,四川甘孜 626000)
为明确冬虫夏草线状菌体制剂的侵染力,进行了冬虫夏草线状菌体制剂对蝠蛾幼虫的感染试验。在相同条件下对蝠蛾幼虫进行人工接种,并对感染温度、菌剂用量和虫龄进行了优化。结果表明冬虫夏草线状菌体具有侵染蝠蛾幼虫的能力,优化后的感染条件为:菌体剂量为5 g(即5个标准剂量)、幼虫虫龄为3龄、培养温度为14 ℃。该结果为蝠蛾幼虫新型接种体的制备与开发提供了依据。
冬虫夏草菌;线状菌体;蝠蛾幼虫;感染
冬虫夏草Ophiocordycepssinensis(Berk.)G.H.Sungetal.(Sungetal.,2007)系我国特产珍稀名贵中药材,是冬虫夏草菌Ophiocordycepssinensis侵入蝠蛾属Hepialus.幼虫体内后形成的一种菌物药(Weietal., 2006),被誉为国菌(Zhangetal., 2015),具有补肺益肾、止血化痰之功效(Bhushanetal., 2013)。
冬虫夏草野生资源匮乏,不能满足市场需求,因此开展冬虫夏草人工繁育势在必行。蝠蛾幼虫接种技术体系是冬虫夏草人工繁育中的关键技术之一(Heetal., 2016),而蝠蛾幼虫被侵染的关键是有效的接种菌剂。目前接种菌剂来源主要有冬虫夏草菌的分生孢子和冬虫夏草菌丝体,分生孢子体作为菌剂报道较多(Lietal., 2013, 2016;Zhangetal., 2016)。由于冬虫夏草菌是极为特殊的一种高原嗜低温型真菌,其生长周期长、生长速度慢、产孢条件苛刻、极易染菌,且冬虫夏草菌多次传代菌种退化严重(Lvetal., 2016),菌剂危机显现,阻碍了冬虫夏草人工培殖的产业化,在蝠蛾幼虫接种菌剂领域寻求新的突破具有重要意义。冬虫夏草线状菌体作为蝠蛾幼虫新的菌剂应用具有培养条件较易、产量高、易于工业化生产的特点,且已有成功诱导线状菌体形成和菌剂制备的报道(Liuetal., 2008;Heetal., 2016),但用线状菌体制备的菌剂对蝠蛾幼虫的侵染效果未见报道。为明确线状菌体菌剂对蝠蛾幼虫的侵染效果,本研究测定了冬虫夏草线状菌体菌剂对蝠蛾幼虫侵染的影响,以期为蝠蛾幼虫新型接种菌剂的制备与开发提供依据。
1 材料与方法
1.1 材料与仪器
供试菌株:冬虫夏草Ophiocordycepssinensis菌种,保存于重庆市中药研究院。
供试虫种:蝠蛾幼虫为贡嘎蝠蛾HepialusgonggaensisFu et Huang幼虫,尹定华研究员鉴定。
主要仪器设备:奥林巴斯CX23显微镜;ZHWY-200B恒温摇床购自上海智诚;UV-1605型紫外可见分光光度计购自岛津;TDL-40B高速离心机购自上海安亭科学仪器厂;SW-CJ-1D超净工作台购自苏州净化设备有限公司;GZX-9070数显鼓风干燥箱购自上海博迅实业有限公司设备厂;电冰箱购自合肥荣事达电冰箱有限公司;SPX-150B-Z生化培养箱购自上海博迅实业有限公司医疗设备厂;高压灭菌锅购自上海博迅实业有限公司。
1.2 方法
1.2.1 冬虫夏草菌线状菌体菌剂的制备
冬虫夏草菌线状菌体的诱导培养:按文献(Heetal., 2016)中所述方法进行培养。
冬虫夏草菌线状菌体的制备:在超净工作台中将发酵培养的发酵液过三号药典筛,弃去固形物,得滤液;再将滤液过九号药典筛,取固形菌体,分散,备用。
线状菌体菌剂的制备:参考文献(Liuetal., 2009)制备法,经改良后:取10 mL菜籽油和4 mL邻苯二甲酸二丁酯于20 mL试管中,加入司班-80 50 μL/mL、硬脂酸铝60 mg/mL,再取上述冬虫夏草线状菌体1 g混入,经充分搅拌分散后混匀,制得1个标准剂量菌种制剂。
1.2.2 虫源饲养
幼虫为人工室内繁育贡嘎蝠蛾HepialusgonggaensisFu et Huang幼虫。蝠蛾成虫交尾后,移至产产卵箱中,待其产卵后收集变黑的卵粒,将卵粒平铺于保湿滤纸上后保温孵化,幼虫孵化后挑取幼虫至饲养盒中饲养(饲养盒中平铺珠芽蓼、人参果、圆穗蓼、胡萝卜等饲料),定期检查并更换饲料。本试验中选取2-3龄幼虫进行试验(除龄期考察外)。
1.2.3 感染率的测定
取标准剂量菌种制剂若干,选取龄期一致的贡嘎蝠蛾幼虫,采用浸渍法接种(Fengetal., 1990),每次选取30头供试幼虫,浸渍10 s,然后取出,用灭菌后的干滤纸吸去虫体表面残余的悬浮液,放入13℃、湿度60%培养箱中饲养,每个处理重复3次,每7天观察记录幼虫体内是否有虫菌体形成,以此计算幼虫感染率(φ):

1.2.4 实验设计及数据处理
单因素实验:首先对影响感染效果的因素分别进行了单因素试验。显著性分析采用多重比较得出,同一列数据中不同字母表示数据存在显著差异,同一列数据若含有相同字母表示不存在显著差异。数据采用spss 19.0软件进行差异显著性分析。
正交实验:为优化感染条件及感染效果,将单因子实验的因素指标进行正交实验考察。各因素选取水平情况、试验结果及分析。显著性分析采用数据采用spss 19.0软件进行差异显著性分析。
2 结果与分析
2.1 单因素实验
2.1.1 不同龄期幼虫对侵染效果的影响
幼虫免疫活性与幼虫发育时期具有相关性(Wuetal., 2010),其体壁附着菌剂的能力和被菌体分泌的胞外酶降解的程序亦不同(Pengetal., 2009),为了解较适宜的幼虫侵染龄期,在其它条件一致的情况下,分别考察了不同龄期幼虫对感染率的影响,结果如表1所示。

表1 不同龄期幼虫感染效果Table 1 The results of instar on Hepialus moths larva infection
注:多重比较显著性检验水平,同一行字母不同表示差异显著(P<0.05)。Note:The different letters indicate there were significant differences (P<0.05).
由表1可得知,幼虫龄期不同,其感染率相差较大。随着幼虫的生长,感染率逐渐上升,3龄幼虫时达到最高感染率26.7%,显著优于其它龄期的侵染率(P<0.05)。随着幼虫进一步生长老熟,感染率有所下降。1龄和5龄幼虫未检测出被感染的幼虫,可能是较嫩或老熟幼虫不利于菌剂感染。
2.1.2 菌体剂量对蝠蛾幼虫侵染效果的影响
分别考察了不同菌体用量配制的菌剂对蝠蛾幼虫侵染效果的影响,其它条件相同,结果见图2。

表2 菌体剂量对蝠蛾幼虫侵染效果的影响Table 2 The effect of filamentous mycelium biomass on Hepialus moths larva infection
注:多重比较显著性检验水平,同一行字母不同表示差异显著(P<0.05)。表2-3同。Note:The different letters indicate there were significant differences (P<0.05).Same to Table 2-3.
实验菌剂所用菌体线状、质轻、分支多、易扭结团聚,因此需用助悬试剂将菌丝分散开来,较多的菌种用量虽可增加侵染的机率,但过多的菌体易发生扭结、团聚,影响侵染率。从表2可看出,菌体用量增大会提高幼虫的感染率,当线状菌体用量为4 g时,其感染率显著优于其它菌种用量的效果(P<0.05)。随着菌体用量继续增加,线状菌体在分散剂中出现团聚现象,进而影响了感染率。故配制菌剂时菌体用量为4 g(即4个标准剂量)。
2.1.3 温度对侵染效果的影响
冬虫夏草菌来源于高原特有生物冬虫夏草,冬虫夏草主产区平均温度长年低于20 ℃,其对温度较敏感(Xingetal., 2008)。因此进行菌体制剂感染实验时对温度的筛选极其重要。其它条件相同,分别研究了不同温度条件下菌体制剂对蝠蛾幼虫感染效果的影响,结果见表3。
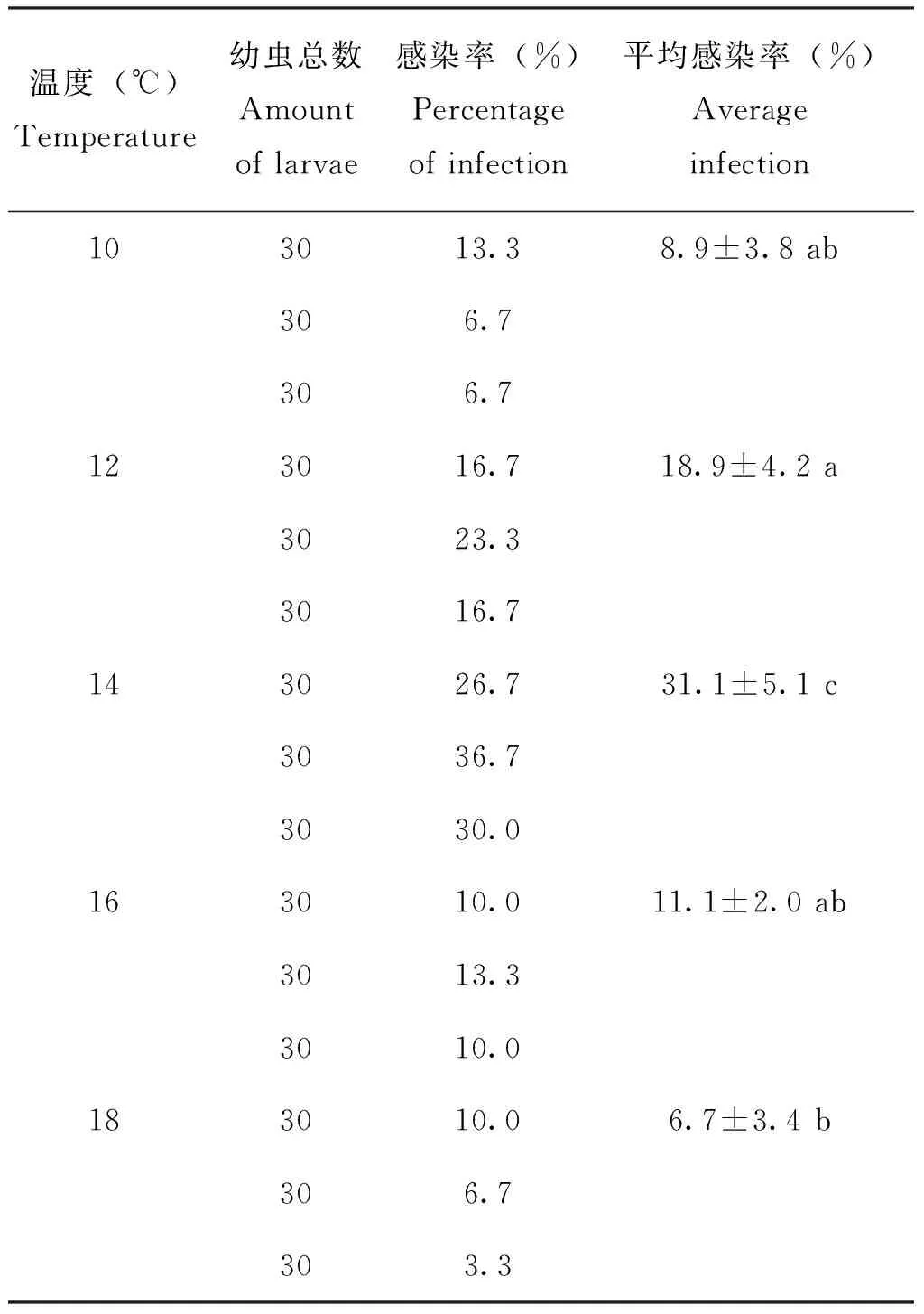
表3 温度对蝠蛾幼虫侵染效果的影响Table 3 The effect of temperature on Hepialus moths larva infection
随着温度的升高菌体顶端生长萌发较快,感染率逐渐升高,当温度超过16℃时蝠蛾幼虫出现生长受阻,能检测到虫菌体的幼虫越来越少,并陆续出现死亡现象,说明温度的升高影响到蝠蛾幼虫生理生长。当培养温度为14℃时,幼虫感染率显著优于其它温度培养下的效果(P<0.05),故选择14℃作为培养温度。
2.2 正交实验
在明确菌体制剂具有侵染力的情况下,为确定主影响因子和较优的侵染条件,对不同龄期幼虫、菌种用量、侵染温度3个因素进行了正交实验优化。根据单因素中3个因素的试验结果进行L4(23)正交实验。各因素选取水平情况见表4,试验及分析结果见表5。
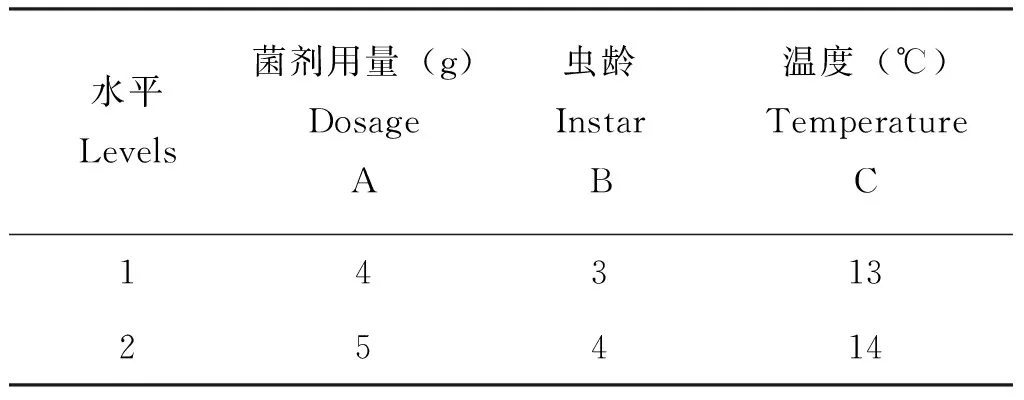
表4 正交实验因素水平Table 4 The factor level oforthogonal experiment

表5 正交试验设计及结果分析Table 5 The orthogonal experiment design and results
注:表中数据为三次重复平均值±标准差。Note: SE, Data in the same column with different capital.
从表5中的结果可表明:培养温度是影响蝠蛾幼虫感染率的最显著因素,其余两个因素的显著性影响程度依次为:幼虫虫龄>菌体剂量。影响蝠蛾幼虫感染率的最佳条件组合为:A2B1C2,即菌体剂量为5 g(即5个标准剂量),幼虫虫龄为3龄,培养温度为14℃。
3 结论与讨论
试验结果表明具有活性的线状菌体制剂对蝠蛾幼虫具有侵染力,优化确定了影响蝠蛾幼虫感染率的主因素并获得了最优培养条件:菌体剂量为5 g(即5个标准剂量)、幼虫虫龄为3龄、培养温度为14℃。
冬虫夏草侵染及形成过程极其复杂,现有研究虽表明冬虫夏草菌是冬虫夏草的唯一无性型菌种(Shenetal., 1983;Liuetal., 1989;Lietal., 2000),但在学术界仍存在较大争议。如近年来研究认为在冬虫夏草成熟过程中,冬虫夏草菌与蝙蝠蛾拟青霉存在DNA共存及竞争性生长,表明冬虫夏草是蝙蝠蛾拟青霉和冬虫夏草菌共同的结果(Zhuetal., 2007;Yangetal., 2008;Gaoetal., 2011;Zhuetal., 2012)。在冬虫夏草分子生物学研究领域不断有新的证据证实野生冬虫夏草是多种真菌、多种突变基因型冬虫夏草菌与蝠蛾幼虫形成的物种复合体,是可培养真菌以及更多不可培养真菌形成的菌物群区系(Zhangetal., 2010a, 2010b)。
本试验研究结果表明线状菌体制剂对蝠蛾幼虫虽有感染力,但感染率均较低,未达到产业化生产的要求,今后工作将重点关注于如何提高侵染率。由于整个实验仍处在冬虫夏草高原产区和使用灭菌土培的形式饲养,在部分环节中是否存在其它土栖真菌或因子的干预,有待进一步确定。同时,线状菌体是如何侵入幼虫体内及侵入期间、侵入后幼虫自身存在何种形态的免疫应答等科学问题则有待进一步的研究。
References)
Gao L, Li XH, Zhao JQ,etal.Detection of multipleOphiocordycepssinensismutants in premature stroma ofCordycepssinensisby Massarray SNP MALDI-TOF mass spectrum genotyping [J].JournalofPekingUniversity(Health Sciences), 2011, 43 (2): 259-266.[高凌, 李晓红, 赵建晴, 等.MassARRAY单核苷酸多态性MALDI-TOF质谱基因分型法检测未成熟冬虫夏草子座中多个冬虫夏草菌突变基因型[J].北京大学学报(医学版), 2011, 43 (2): 259-266]
He ZY, Li L, Zhang DL,etal.The research on the proliferative culture ofOphiocordycepssinensisfilamentous mycelium [J].JournalofEnvironmentalEntomology, 2016, 38 (6): 1205-1213.[贺宗毅, 李黎, 张德利, 等.冬虫夏草菌丝状菌源增殖培养的研究[J].环境昆虫学报, 2016, 38 (6): 1205-1213]
Lei YY, Lv LH, He YR,etal.Correlation between biological characteristics ofBeauveriabassianaand its virulence toPlutellaxylostella[J].ChineseJournalofBiologicalControl, 2010, 26 (2): 143-148.[雷妍圆, 吕利华, 何余容, 等.球孢白僵菌生物学特性与其对小菜蛾致病力相关性分析[J].中国生物防治, 2010, 26 (2): 143-148]
Li JH, Xie F, Wang C,etal.The microstructure and formation of the conidia ofHirsutellasinensis[J].ChineseJournalofMicroecology, 2013, 25 (7): 775-781.[李建宏, 谢放, 王灿, 等.中国被毛孢分生孢子形成过程显微结构的研究[J].中国微生态学杂志, 2013, 25 (7): 775-781]
Li YL, Luo HC, Hu M.Research on the artificial cultivation of the conidia byOphiocordycepssinensis[J].FoodResearchandDevelopment, 2016, 37 (3): 187-190.[李玲玲, 罗合春, 胡敏.人工培育冬虫夏草分生孢子的研究[J].食品研究与开发, 2016, 37 (3): 187-190]
Liu HB, Wang GX, Mao XB,etal.Optimization on oil suspension formulation ofCordycepssinensisspawn [J].Microbiology, 2009, 36 (10): 1557-1563.[刘洪波, 王贵学, 毛先兵, 等.正交优化冬虫夏草菌种油混悬制剂[J].微生物学通报, 2009, 36 (10): 1557-1563]
Liu HB, Wang GX, Zhu HL,etal.The microstructure and formation of the conidia ofHirsutellasinensis[J].ChineseJournalofBiologicalControl, 2008, (s1): 106-110.[刘洪波, 王贵学, 朱华李, 等.冬虫夏草菌种油混悬剂的研究[J].中国生物防治学报, 2008, (s1): 106-110]
Peng GL, Xue JW, Liu WM,etal.Role of protease chitinase ofVerticilliumlecaniiin infecting scale insect cuticle[J].ChineseJournalAppliedandEnvironmentalBiology, 2009, 15 (2): 220-225.[彭国良, 薛皎亮, 刘卫敏, 等.蜡蚧轮枝菌入侵蚧虫表皮过程中蛋白酶和几丁质酶的作用[J].应用与环境生物学报, 2009, 15 (2): 220-225]
Shen NY, Zeng L, Zhang XC,etal.The separation ofOphiocordycepssinensisfungi [J].EdibleFungi, 1983, 5 (1): 1-5.[沈南英, 曾璐, 张显耻, 等.冬虫夏草真菌的分离[J].食用菌, 1983, 5 (1): 1-5]
Shrestha B, Zhang WM, Zhang YJ,etal.What is the Chinese caterpillar fungusOphiocordycepssinensis(Ophiocordycipitaceae) [J].Mycology, 2010, 1 (4): 228-236.
Wei XL, Yin XC, Guo YL,etal.Analyses of molecular systematics onCordycepssinensisand its related taxa [J].Mycosystema, 2006, 25 (2): 192-202.[魏鑫丽, 印象初, 郭英兰, 等.冬虫夏草及其相关类群的分子系统学分析 [J].菌物学报, 2006, 25 (2): 192-202]
Wu BS.The Difference of Immunocomptence during Development in the Mealworm Beetles,Tenebriomolitor(Insecta Coleptera) [D].Suzhou: Suzhou University, 2010.[吴必顺.黄粉虫发育过程中免疫活性差异研究[D].苏州: 苏州大学, 2010]
Xing XK, Guo SX.The structure and histochemistry of sclerotia ofOphiocordycepssinensis[J].Mycologia, 2008, 100 (4): 616-625.
Yang JL, Xiao W, He HX,etal.Molecular phylogenetic analysis ofPaecilomyceshepialiandCordycepssinensis[J].ActaPharmaceuticaSinica, 2008, 43 (4): 421-426.[杨金玲, 肖薇, 何惠霞, 等.蝙蝠蛾拟青霉与冬虫夏草关系的分子系统学研究[J].药学学报, 2008, 43 (4): 421-426]
Zhang YJ, Li EW, Wang CS,etal.Ophiocordycepssinensis, the flagship fungus of China: Terminology, life strategy and ecology [J].Mycology, 2012, 3 (1): 2-10.
Zhang YJ, Sun BD, Zhang S,etal.Mycobiotal investigation of naturalOphiocordycepssinensisbased on culture-dependent investigation [J].Mycosystema, 2010a, 29 (4): 518-527.[张永杰, 孙炳达, 张姝, 等.分离自冬虫夏草可培养真菌的多样性研究[J].菌物学报, 29 (4): 518-527]
Zhang YJ, Zhang S, Wang M,etal.High diversity of the fungal community structure in naturally occurringOphiocordycepssinensis[J].PLoSONE, 2010b, 5 (12): 1-8.
Zhang ZY, Liang GH, Liang L,etal.Effects of medium and environmental conditions on the sporulation ofOphiocordycepssinensisin solid fermentation [J].Mycosystema, 2016, 35 (4): 440-449.[张宗耀, 梁关海, 梁蕾, 等.培养基及培养条件对冬虫夏草固体发酵产分生孢子的影响[J].菌物学报, 2016, 35 (4): 440-449]
Zhu JS, Guo YL, Yao YS,etal.Coexistence ofHirsutellasinensisandPaecilomyceshepialiDNA during maturation ofCordycepssinensisand changes in fungal competitive proliferation capability and chemical profiles [J].JournalofFungalResearch, 2007, 5 (4): 214-224.[朱佳石, 郭英兰, 姚艺桑, 等.冬虫夏草成熟过程中冬虫夏草和蝙蝠蛾拟青霉DNA共存及竞争增殖力、化学成分变化[J].菌物研究, 2007, 5 (4): 214-224]
Zhu JS, Zhao JG, Gao L,etal.Dynamically altered expressions of at 6Ophiocordycepssinensismutants in the stroma ofCordycepssinensis[J].JournalofFungalResearch, 2012, 10 (2): 100-112.[朱佳石, 赵家刚, 高凌, 等.至少6个突变基因型冬虫夏草菌在冬虫夏草子座中表达的动态变化[J].菌物研究, 2012, 10 (2): 100-112]
The infection research on theOphiocordycepssinensisfilamentous mycelium toHepialusmoths larvae
HE Zong-Yi1,2,3,4,ZHANG De-Li1,2,3,4,LI Li1,4,XING Kang-Kang1,2,3,4,CHEN Shi-Jiang1,2,3,4*
(1.Chongqing Academy of Chinese Materia Medica, Chongqing 400065, China; 2.Chongqing Key Laboratory of Chinese Medicine Resources, Chongqing 400065, China; 3.Chongqing Sub-center of the National Resource Center for Chinese Materia Medica, China Academy of Chinese Medical Sciences, Chongqing 400065, China; 4.Ganzi Tibetan Autonomous Prefecture Kangding GonggaOphiocordycepssinensisIndustry CO.LTD., Ganzi 626000, Sichuan Province, China)
In order to clear the infection of theOphiocordycepssinensisfilamentous mycelium to theHepialusmoths larvae, the infection ofO.sinensisfilamentous mycelium toHepialusmoths larvae was tested by artificial fed at the same environment and condition.Then, the infected conditions were optimized by orthogonal experiment design.The results showed that the infected conditions were amount ofO.sinensisfilamentous mycelium 5 g, instar 3th, temperature 14℃, In summery, theO.sinensisfilamentous mycelium had the infection to theHepialusmoths larvae.The result had provided theoretical basis to the reforming ofO.sinensisfilamentous mycelium inoculant preparation.
Ophiocordycepssinensis; filamentous mycelium;Hepialusmoths larva; infect
贺宗毅,张德利,李黎,等.冬虫夏草线状菌体制剂对蝠蛾幼虫感染的研究[J].环境昆虫学报,2017,39(3):673-678.
重庆市科技平台项目(cstc2014ptyjd10001);重庆市科技惠民计划项目(2013gs500102-D2014-3);重庆市社会民生科技创新专项(cstc2015shmszx120032);重庆市科技攻关计划项目(cstc2013yykfB110002)
贺宗毅,男,1986年生,重庆人,硕士研究生,助理研究员,研究方向为冬虫夏草人工培殖,E-mail: cqhcmthzy2004@163.com
*通迅作者Author for correspondence,E-mail:shijiangchen@163.com
Received: 2017-03-10;接受日期Accepted: 2017-04-10
Q965
A
1674-0858(2017)03-673-06
猜你喜欢
山东冶金(2022年2期)2022-08-08
发明与创新·小学生(2021年12期)2021-01-06
军事文摘(2020年22期)2021-01-04
天津农林科技(2020年3期)2020-08-13
园林科技(2020年2期)2020-01-18
今日农业(2019年11期)2019-08-13
山东冶金(2019年1期)2019-03-30
现代农业(2016年5期)2016-02-28
山东青年(2016年2期)2016-02-28
汉语世界(The World of Chinese)(2016年3期)2016-01-10
