
库布齐沙地人工生物结皮区植被恢复状况
2017-07-05 02:28吴沛沛杨矿涛
安徽农业科学 2017年14期
关键词:植被恢复
吴沛沛 杨矿涛

摘要[目的]调查库布齐沙地人工生物结皮区高等植被恢复状况,揭示人工生物结皮形成后高等植被群落演替过程。[方法]采用样方法对不同接种年份的人工生物结皮区高等植被的种类、数量、盖度进行系统全面的调查。[结果]共发现42种高等植物,分属于31属10科,优势种为油蒿、沙蓬、绳虫实、猪毛蒿、羊草、披碱草。随着人工生物结皮发育演替时间的延长,高等植被的种类、数量、盖度、生物量、物种多样性和均匀度也逐渐增大。[结论]在人工结皮区,高等植被群落的演替顺序为油蒿、沙蓬、猪毛蒿、羊草群落。
关键词库布齐沙地;植被恢复;生物结皮
中图分类号S181.3;X171.4文献标识码A文章编号0517-6611(2017)14-0060-03
Abstract[Objective]By investigating vegetation restoration at manmade biological soil crusts areas in Kubuqi Desert,succession process of vegetation was revealed.[Method]Based on the survey of community plots,species,biomass and coverage of vegetation were investigated within different inoculation of manmade biological crusts areas.[Result] 42 species vascular plants(belonging to 31 genera and 10 families,respectively)were found,and the dominant species were Artemisia ordosica,Agriophyllum squarrosum,Corispermum declinatu,Artemisia scoparia,Leymus chinensis,and Elymus dahuricus.Along with succession of manmade biological soil crusts,there were increases in vascular vegetations species,abundances,coverages,biomass,species diversity and evenness.[Conclusion]Moreover,successions order of vascular vegetations were Artemisia ordosica, Agriophyllum squarrosum, Artemisia scoparia, Leymus chinensis.
Key wordsKubuqi Desert;Vegetation restoration;Biological soil crusts
在植被稀少的荒漠半荒漠地區,生物结皮能够适应高温、高盐碱、强辐射、风沙侵蚀等恶劣环境,从而很好地存活和大量分布。它们能够固定沙丘,减少风沙侵蚀,改善土壤水分平衡,增加土壤营养,因此受到国内外学者的广泛关注[1-2]。在腾格里沙漠、甘肃河西走廊地区、古尔班通古特沙漠、库布齐沙漠,藻类工作者先后进行了生物结皮的空间分布、种类组成、藻类种类、胶结机理和细微结构以及荒漠藻类在固沙成土等研究[3-5]。同时,从自然生物结皮中分离筛选到适合于固沙的荒漠藻类。刘永定[6]提出了利用荒漠藻类固沙的理念,并逐步完善成熟,形成了荒漠藻人工生物结皮综合固沙技术。人工生物结皮形成后,它们的先锋拓殖作用改善了周围的生态环境,为荒漠-半荒漠生态系统向草地生态系统的演替提供了条件。笔者通过调查不同接种年份的荒漠藻人工生物结皮区,对高等植被恢复状况进行系统的生态调查,以揭示人工生物结皮形成后高等植被群落演替过程。
1研究区域与方法
1.1研究区域研究区域位于库布齐沙地人工生物结皮接种1年区、2年区、4年区、5年区、6年区和7年区。选择北坡、南坡和丘间地为试验样地(每块样地选取6个1 m×1 m的样方)。记录样方内高等植被的种类、数量、高度、盖度等内容。
1.2试验方法
1.2.1相对重要值。物种重要值(Important Value,IV)=相对密度+相对频度+相对盖度,其中IV>1.5的物种为优势种,IV>0.8的物种为亚优势种,IV>0.1的物种为伴生种,IV<0.1的物种为偶见种。
相对密度=(某一物种的个体数/群落中所有物种个体数)×100%
相对频度=(某一物种出现的样方数/样方总数)×100%
相对盖度=(某一物种的分盖度/群落植被的总盖度)×100%
1.2.2多样性指数。
Shannon-Wiener指数:H=-(PilnPi)
Simpson指数:D=1-(Ni/N)2
Pielou均匀度指数:E=-PilnPi/lnS
Margalef丰富度指数:D=(S-1)/lnN
式中,S为物种总数;N为物种总个体数;Ni为第i种个体数;Pi为物种i的个体数在群落全部个体数中所占比例。
1.3统计分析各指标之间的方差分析运用one-way ANOVA在0.05水平进行分析,数据分析在SPSS 16.0软件中进行。根据优势种和亚优势种的重要值,对不同样地群落组成用Canoco 5.0软件进行主成分分析。
2结果与分析
2.1不同年份人工生物结皮区高等植被的生物量和盖度变化由表1可知,隨着微生物修复时间的延长,植被的盖度也逐渐增大,1年区植被盖度最小,7年区的植被盖度最大。各年份的人工结皮区,均呈现丘间地最大,北坡次之,南坡最小的特点。1年区植被盖度不足10.00%,各立地条件之间植被盖度无显著差异(P>0.05)。2年区比1年区植被盖度增加了0.5~1.0倍,但是与1年区无显著差异(P>0.05)。4年区植被盖度是1年区植被盖度的约3.0倍,且丘间地与北坡植被盖度差异不大,而北坡和丘间地与南坡的差异有所扩大。5年区植被盖度比4年区增加了约1.0倍,北坡和丘间地植被盖度是北坡植被盖度的1.5倍。6年区北坡的植被盖度比5年区有轻微下降,南坡和丘间地植被盖度仍有大幅增加。7年区丘间地的植被盖度高达56.77%,与1年区植被盖度相比,增加了约6.0倍。
各年份人工结皮区植被的生物量从大到小依次为丘间地、北坡、南坡(表1),且植被的生物量随结皮发育演替时间的推延而增大。1年区各立地条件之间植被的生物量无显著差异(P>0.05)。2、4年区植被的生物量北坡显著大于南坡(P<0.05)。5年区南坡的生物量比4年区增加了65.38%,而丘间地植被的生物量约为南坡的2.0倍。6年区丘间地植被生物量最大,而在南坡的生物量同比5年区接种区是有所下降,北坡植被生物量与5年区无显著差异(P>0.05)。7年区南坡的生物量显著大于6年区南坡(P<0.05),但与5年区南坡无显著差异(P>0.05);丘间地和北坡的植被生物量与6年区相同立地条件的植被生物量相比略微下降,但与6年区的植被生物量无显著差异(P>0.05)。
2.2不同年份人工生物结皮区高等植被群落中的物种组成在人工生物区共发现高等植物42种,分属于31属10科,其中有80.95%的物种属于禾本科(7种)、菊科(14种)、豆科(6种)和藜科(7种),且物种个体数量较多,出现的频率较高,盖度也较大。萝藦科(2种)、紫草科(1种)、紫葳科(1种)、旋花科(1种)、蓼科(2种)、蒺藜科(1种)这6科的物种仅在一两个样方出现,个体数量均在10个以下,相对盖度不足1%。在1~4年区,高等植物的物种数逐渐增加了4.4倍;4年以后的区高等植被种类变化不大,但群落中各物种的重要值有所变化,优势种也在更替。根据物种组成以及优势种的不同,一些物种形成各种不同的小群落,主要有油蒿、大赖草、沙蓬、绳虫实、沙蓬+绳虫实、猪毛蒿、羊草、披碱草、羊草+披碱草、披碱草+拂子茅、拂子茅等一些小群落。
1年区的优势种为油蒿,大赖草、绳虫实、雾冰藜为伴生种,沙蓬为偶见种,这些物种均为旱生型或者超旱生型。2年区出现了2种豆科植物,猪毛蒿为优势种,羊草和沙蓬为亚优势种。4年区的高等植物种类比2年区增加了1.0倍,优势种仍是猪毛蒿,但是羊草的重要值有所增加,说明该地区正在向羊草群落演替;菊科植物的物种数由2年区的2种增加到9种,藜科植物由2年的3种增加至7种;另外,在丘间地还发现了中生型的菟丝子,主要寄生在绳虫实和大籽蒿的植物体上。5年区的高等植被群落组成与4年区差别不大,但是猪毛蒿的重要值开始下降。6年区出现了猪毛蒿和羊草2种优势种共存的局面;亚优势种为披碱草,而且拂子茅的重要值也较高,为0.534,说明该地区的蒿类植物群落正在被禾本科植物群落取代。7年区的优势种为羊草,猪毛蒿和披碱草为亚优势种,旱中生型的大籽蒿和中生型的狭叶青蒿及野艾蒿的重要值都增加了1.0倍以上。
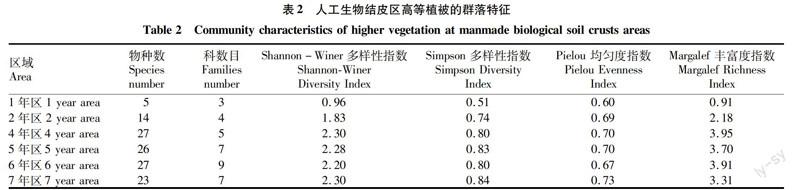
2.3不同年份人工生物区高等植被群落特征由表2可知,随着人工结皮的发育演替,高等植物的多样性逐渐增加,Shannon-Winer多樣性指数和Simpson多样性指数的变化趋势基本一致,而Pielou均匀度指数也呈现出逐渐增大的趋势。这说明在人工结皮区,人工结皮的发育演替与高等植被群落的恢复呈现互相促进的关系。人工生物结皮在流沙表面形成后,沙土表面逐渐稳定,其水分营养状况也逐渐改善,为高等植物的生长提供了有利条件。而高等植物的存在又为生物结皮提供一定的遮阴作用,使得一些地区的水肥条件逐渐改善,促进了藓类的生存和发育,从而促进了生物结皮向藓类结皮演替。
随着人工结皮的发育演替,高等植被的物种数目逐渐增加,由1年区的5种增加到4年區的27种。而后虽然高等植物的物种数几乎没有变化,但由原来5个科增加到9个科,这是由于5、6年区高等植物群落正在经历由猪毛蒿群落向羊草群落的演替,猪毛蒿群落和许多伴生种都存在其中,因此其种数和科数都比较多。而7年的生物结皮区,羊草群落逐渐占优势,使得群落内羊草(如山苦荬、披碱草和拂子茅等)物种的数目和盖度逐渐增加;而猪毛蒿群落的伴生种蓝刺头、披针叶黄华、苦豆子、苦马豆消失。因此,该地区的物种数减少到23种,所属的科也减少到7个科(表2)。Margalef丰富度指数的变化过程也很好地说明了这个问题。
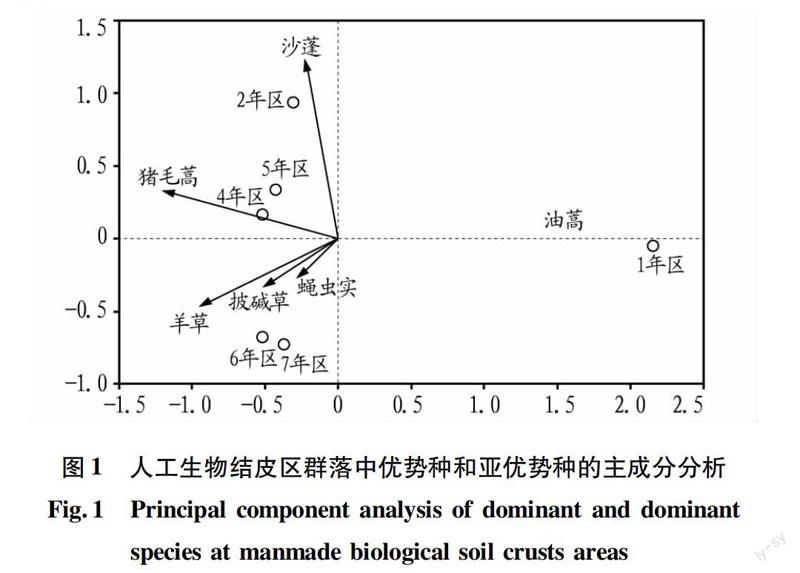
将群落中重要值大于0.8的6个物种油蒿、沙蓬、绳虫实、猪毛蒿、羊草、披碱草与各接种年份生物结皮区进行主成分分析(图1)。发现第一、二序轴解释了物种数据的97.80%。图中带箭头的射线代表物种,空心圆点代表各年份人工结皮区的样方。将物种的射线延长,6个样方垂直投影到物种的射线上,以投影点到物种实心箭头处相对距离为标准,沿着箭头方向为增大,反之为减小。在人工结皮区,高等植被群落沿着油蒿→沙蓬→猪毛蒿→羊草群落的顺序进行演替。图中2个物种之间夹角的余弦代表物种之间的相关性,夹角越大,相关性越小。羊草与绳虫实和披碱草的夹角很小,说明羊草与绳虫实和披碱草的相关性很大,绳虫实和披碱草是羊草的重要伴生种。羊草与猪毛蒿呈45°的锐角,说明羊草与猪毛蒿也呈正相关。而羊草、沙蓬、油蒿三者之间为钝角,说明这3个物种之间呈微弱的负相关。而猪毛蒿和羊草与油蒿几乎呈180°,说明它们和油蒿呈显著负相关。
3结论
在荒漠半荒漠地区,退化的高等植被群落能以较快的速度进行重新组织和演替,其根本原因在于植物物种具有拓殖能力和群落资源过剩。拓殖能力是植物的本能,过剩的资源则是高等植被演替的物质条件[7]。如果系统中存在过剩的资源,群落中就会有新的物种加入或某些种群增长。结皮的形成、发育、演替及资源的积累,为植被的演替提供了物质基础。随着结皮的发育演替,土壤中碳、氮、磷和有机质等营养物质逐渐增加,细菌、真菌数量也逐渐增加,它们通过生命活动促进了各种物质的转化,这就为荒漠生态系统提供了过剩的资源。同时由于人工生物结皮稳定了土表,增加了表面粗糙度,因此土壤种子库在结皮形成后增加[8],这就为高等植物的拓殖提供了可能。一旦条件适宜,种子萌发生长,新的物种出现。
在流沙表面接种荒漠藻类后,人工生物结皮在1个月左右的时间形成,使松散的沙粒相互胶结在一起,使流沙表面逐渐被固定[5]。随着人工结皮的发育演替,高等植物的多样性逐渐增加,Shannon-Winer多样性指数和Simpson多样性指数的变化基本一致,而Pielou均匀度指数也呈现出逐渐增大的趋势。这说明在人工结皮区,人工结皮的发育演替与高等植被群落的恢复呈现互相促进的关系。人工生物结皮在流沙表面形成后,沙土表面逐渐稳定下来,其水分营养状况也逐渐改善,为高等植物的生长提供了有利条件。而高等植物的存在又为生物结皮提供一定的遮阴作用,使得一些地区的水肥条件逐渐改善,促进了藓类的生存和发育,从而促进生物结皮向藓类结皮演替。
在温带和寒温带的沙漠地区,从裸露的沙丘演替到较稳定的森林群落,约需要1 000年的时间[9]。而人对生物群落演替的影响远远超过其他所有的自然因素,由此人为参与可以加速生态系统中群落演替的进程。在没有接种人工结皮的地区,油蒿就能够在流动沙丘旺盛生長,随着生物结皮的形成,油蒿的生长能力降低,大量油蒿逐渐枯死,被其他物种所代替[10-11]。在1年区为优势种的油蒿,随着结皮的演替发育,其优势度逐渐下降,到了7年区,重要值仅为0.079。值得一提的是4年区牛心朴子的出现,是荒漠化过程中最后阶段的指示种,是荒漠草原沙化标志之一。如果油蒿群落被牛心朴子侵占,则生态系统向荒漠化方向逆向演替[12]。而4年区猪毛蒿占优势,羊草为亚优势种;至6年区,猪毛蒿和羊草共同成为亚优势种,牛心朴子消失,说明高等植被群落正在向羊草群落演替,该过程为正向演替。总体来看,随着接种年份的增加,人工生物结皮区高等植被群落的演替顺序依次为油蒿、沙蓬、猪毛蒿、羊草。
参考文献
[1] BELNAP J,PHILLIPS S L,WITWICKI D L,et al.Visually assessing the level of development and soil surface stability of cyanobacterially dominated biological soil crusts[J].J Arid Environ,2008,72(7):1257-1264.
[2] ELDRIDGE D J,FREUDENBERGER D,KOEN T B.Diversity and abundance of biological soil crust taxa in relation to fine and coarsescale disturbances in a grassy eucalypt woodland in eastern Australia[J].Plant and soil,2006,281(1):255-268.
[3] 刘永定,黎尚豪.土壤藻类及其生理生态[J].水生生物学报,1993,17(3):272-277.
[4] 胡春香,刘永定,张德禄,等.荒漠藻类拓殖演替及其影响因子[J].应用生态学报,2003(14):11-12.
[5] LAN S B,WU L,ZHANG D L,et al.Analysis of environmental factors determining development and succession in biological soil crusts[J].Sci Total Environ,2015,538:492-499.
[6] 劉永定.荒漠蓝藻环境生物学与生物土壤结皮固沙[M].北京:科学出版社,2013:199-212.
[7] 王炜,刘钟龄,郝敦元,等.内蒙古草原退化群落恢复演替的研究:I.退化草原的基本特征与恢复演替动力[J].植物生态学报,1996,20(5):449-459.
[8] 苏延桂,李新荣,张景光,等.生物土壤结皮对土壤种子库的影响[J].中国沙漠,2006,26(6):997-1001.
[9] OLSON J S.Rates of succession and soil changes on Southern Lake Michigan sand dunes[J].Botanical Gazette,1958,119(3):125-170.
[10] 杨宝珍,董学军,高琼,等.油蒿(Artemisia ordosica)的蒸腾作用及其群落的水分状况[J].植物生态学报,1994,18(2):161-170.
[11] LAN S B,WU L,ZHANG D L,et al.Desiccation provides photosynthetic protection for crust cyanobacteria Microcoleus vaginatus from high temperature[J].Physiol Plant,2014,152(2):345-354.
[12] 程晓莉,安树青,钦佩,等.鄂尔多斯草地退化过程中植被地上生物量空间分布的异质性[J].生态学报,2003,23(8):1526-1532.
猜你喜欢
绿色科技(2016年22期)2017-03-15
绿色科技(2017年3期)2017-03-14
现代园艺(2016年13期)2016-09-02
绿色科技(2015年6期)2015-08-05
