
基于微卫星标记的苹果绵蚜种群遗传结构动态
2017-06-19 18:24:10杨明明谭秀梅魏成民张永晓万方浩周洪旭青岛农业大学农学与植物保护学院山东省植物病虫害综合防控重点实验室山东青岛6609中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室北京0093
生物安全学报 2017年2期
杨明明, 谭秀梅, 魏成民, 张永晓, 万方浩,, 周洪旭*青岛农业大学农学与植物保护学院,山东省植物病虫害综合防控重点实验室,山东 青岛 6609; 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 0093
基于微卫星标记的苹果绵蚜种群遗传结构动态
杨明明1, 谭秀梅1, 魏成民1, 张永晓1, 万方浩1,2, 周洪旭1*
1青岛农业大学农学与植物保护学院,山东省植物病虫害综合防控重点实验室,山东 青岛 266109;2中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193
【目的】苹果绵蚜是我国重要的入侵害虫,对苹果生产造成了严重危害。近年来,苹果绵蚜扩散面积增大,危害加重。了解苹果绵蚜入侵过程中的分子生态变化,可为该虫的综合防控提供依据。【方法】利用微卫星标记技术,选择6个微卫星位点对山东省6个地区(烟台、威海、青岛、潍坊、聊城、泰安)2012—2015年苹果绵蚜种群遗传结构变化规律进行分析。【结果】2012—2015年,山东省6个地区的苹果绵蚜遗传多样性随时间推移逐渐降低。其中,2012年的遗传多样性极显著高于2013—2015年;2013—2015年之间虽然差异不显著,但随时间推移,等位基因观测值(Na)和期望杂合度(He)等遗传多样性指数有逐渐降低的趋势。通过无限等位基因模型、双相突变模型和逐步突变模型分析发现,6个地区的苹果绵蚜均经历了瓶颈效应,是遗传多样性降低的主要原因。对6个微卫星位点分析发现,Erio20、 Erio75和Erio78扩增到的苹果绵蚜等位基因数量以及这3个位点的多样性指数随时间推移逐渐降低。【结论】Erio20、 Erio75和Erio78是引起苹果绵蚜遗传多样性降低的主要多态位点。苹果绵蚜可能进化出了“超级克隆”基因型,因此,其可在我国适应不同生态环境并增大扩散范围。
苹果绵蚜; 入侵害虫; 微卫星; 遗传多样性; 动态
苹果绵蚜Eriosomalanigerum(Hausmann),别名绵蚜、血色蚜虫、赤蚜、白毛虫,属半翅目瘿绵蚜科绵蚜属,是一种重要的世界检疫性害虫,也是我国重要的入侵生物之一(郭小侠等,2011; 贺春玲等,2004; 王兴亚等,2011; 吴海军等,2007; Ruiz-Montoyaetal.,2015)。苹果绵蚜通常聚集在苹果树剪锯口和伤口、树皮的裂缝、新梢叶腋、果枝顶部、果柄、果实梗洼和萼洼处以及浅根部进行刺吸危害,降低果实的品质与产量,严重威胁苹果生产(王西存等,2011; Ateyyat & Alantary,2009; Havilletal.,2006; Kelmetal.,2004; Nicholasetal.,2005)。20世纪五六十年代,我国通过连续施用药剂、释放天敌日光蜂Phelinusmali(Haldeman)和种植抗性品种等措施,将苹果绵蚜控制在了较低水平,但自90年代中期开始,苹果绵蚜又在我国进一步扩散蔓延 (Ateyyat & Alantary,2010),呈现自东向西扩大危害的态势。近年来,该虫已传到江苏(赵增锋,2012)、河北(张福芹,2002)、天津(Zhouetal.,2013)、山西(董锦花等,2008)、河南(李定旭等,2003)、新疆等地(王朴等,2007; 于江南等,2008),并且在新疫区危害日趋严重。
生物为了生存而适应环境,种群遗传多样性和等位基因分布可能随着环境因素的变化而变化,因此,遗传多样性能够反应生物对环境变化的适应能力(段云,2013)。随着分子生物学在昆虫学中的应用日趋广泛,遗传多样性在农业害虫防治中发挥了重要作用。近年来,全球环境条件发生较大变化,害虫遗传动态规律成为研究热点。Lavanderoetal.(2009)以智利中部的27个苹果绵蚜种群为研究对象,发现8对微卫星引物对苹果绵蚜均具有多态性,每个位点含3~11个等位基因,观测杂合度为0.41~0.93,证明8对微卫星引物可用于苹果绵蚜种群遗传研究。Zhouetal.(2015)通过微卫星标记中国24个苹果绵蚜种群,发现山东、辽宁和邻近地区的19个苹果绵蚜种群具有相似的遗传多样性和遗传结构,形成一个不同于云南和新疆的庞大的随机交配种群。Timmetal.(2005)运用扩增片段长度多态性(amplified fragment length polymorphism,AFLP)手段对南非苹果绵蚜孤雌生殖种群的遗传结构及遗传多样性进行研究,发现苹果绵蚜种群表现低水平的遗传变异。然而,有关苹果绵蚜遗传结构动态的报道较少。
本文利用微卫星标记(simple sequence repeat,SSR)技术,研究山东省6个地区苹果绵蚜2012—2015年遗传多样性变化情况,以期为该虫的综合防控提供依据。
1 材料与方法
1.1 苹果绵蚜样本采集
于2012年7月、2013年7月、2014年8月、2015年8月,在山东省青岛市、聊城市、烟台市、潍坊市、泰安市、威海市6个地区采集苹果绵蚜成虫。将采集到的24个种群共412个苹果绵蚜样本放入离心管中,浸泡在100%无水乙醇中,在-20 ℃冰箱中保存待用。
1.2 苹果绵蚜DNA提取
苹果绵蚜DNA提取参照Sunnucks & Hales (1996)和Weir & Cockerham (1984)的方法,稍有改动。先将单头苹果绵蚜放入0.2 mL离心管中,加入10 μL裂解液(0.45% Tween-20、pH=8.4的0.45% NP-40、10 mmol·L-1Tris-HCl、50 mmol·L-1KCl),用10 μL封口枪头将苹果绵蚜研磨碎,再加入90 μL裂解液。65 ℃温浴15 min,95 ℃加热10 min后,储存于-20 ℃以下备用。
1.3 SSR的PCR扩增、电泳及检测
SSR的PCR所用引物(表1)设计参照Lavanderoetal. (2009)的方法。反应体系(13 μL):10×Buffer (+Mg2+) 1.30 μL,dNTPs 1.04 μL,上、下游引物分别0.26 μL,ddH2O 7.94 μL,Taq酶(PC1100,Solarbio,5 U·μL-1) 0.2 μL,模板2 μL。反应程序:94 ℃预变性4 min;94 ℃变性30 s,50~60 ℃复性45 s,72 ℃延伸1 min,共35个循环;最后72 ℃ 7 min。PCR产物用1%琼脂糖凝胶电泳分离,经溴化乙锭染色后在紫外凝胶成像仪上观察,选取电泳效果好的产物送上海生物工程有限公司进行STR(short tandem repeat)检测。
1.4 数据处理
利用软件Popgene (version 1.32)计算每个种群的等位基因观测值(number of alleles observed,Na)、等位基因有效值(number of effective alleles,Ne)、期望杂合度(expected heterozygosity,He)和Nei′s期望杂合度(Nei′s expected heterozygosity,Nei),并对成对种群遗传分化指数FST与基因流情况Nm进行分析。其中,基因流根据公式Nm=(1/FST-1)/2计算得出(Yehetal.,1999);利用FSTAT 2.9.3.2 计算各个种群的基因多样性(heterogeneous stock,Hs)和等位基因丰富度(allelic richness,Ar)(Franksetal.,2010);利用GENEPOP v3.4计算群体间的遗传分化指数FST(Weir & Cockerham,1984);利用BOTTLENECK检验偏离突变—漂流平衡(Cornuet & Luikart,1996),通过3个突变模型(双相突变模型、无限等位基因模型和逐步突变模型)检测不同支系的每个种群是否存在瓶颈效应。双相突变模型默认参数设置为30%来自无限等位基因模型与70%来自逐步突变模型。利用软件STRUCTURE (Pritchardetal.,2000)对所有种群进行贝叶斯聚类分析,进行2万次的burn-in和5万次的MCMC (Markov Chain Monte Carlo)运算。分析时,从1到20共计算20个K值,每个K值重复6次;利用Evanno (2005)的方法计算分组数ΔK。

表1 苹果绵蚜的6个SSR位点及其对应的引物序列信息Table 1 Information of six SSR E. lanigerum loci and their primer sequences
2 结果与分析
2.1 山东省6个地区苹果绵蚜种群的遗传多样性
由图1可以看出,随着时间推移,6个地区的苹果绵蚜遗传多样性指数均表现逐渐降低的趋势。在P<0.05水平上,泰安、聊城种群的Na和Ne无显著差异,泰安种群2012年的其他遗传多样性指数均显著高于2013—2015年,聊城种群2012和2013年的其他遗传多样性指数均显著高于2014—2015年;威海和青岛种群的Na和Ne也无显著差异,且2个地区苹果绵蚜种群2012年的其他遗传多样性指数均显著高于2013—2015年;潍坊种群的Ne无显著差异,2012年的Na、Hs、He和Nei值均显著高于2013—2015年,2012—2013年的Ar显著高于2014—2015年;烟台种群的Na无显著差异,2012—2014年的Ar显著高于2015年,2012年其他遗传多样性指数均显著高于2013—2015年。总体而言,6个地区苹果绵蚜2012年的遗传多样性显著高于2013—2015年。
2.2 山东省6个地区苹果绵蚜种群的遗传分化
对6个地区苹果绵蚜不同年份种群间遗传分化指数FST进行显著性(P<0.05)分析(表2)可知,2012年烟台种群FST与其他种群具有显著差异,其余种群间差异均不显著;2013年威海与烟台种群、威海与潍坊种群、威海与泰安种群FST差异显著,其余种群间差异均不显著;2014—2015年6个地区苹果绵蚜种群的FST均没有显著差异。种群间的基因流结果显示,2012—2015年6个地区间Nm的范围分别为0.594~1.012、0.902~1.004、0.931~1.005、1.000~1.005,其中,2012年的基因流变化范围最大。这说明苹果绵蚜种群在不同地区间没有遗传分化,基因流水平基本一致。
2.3 山东省6个地区苹果绵蚜种群遗传动态
分别对不同地区不同年份苹果绵蚜的瓶颈效应进行检测(表3),发现在无限等位基因模型、双相突变模型和逐步突变模型下,泰安苹果绵蚜种群2014和2015年均表现显著的杂合度过剩(威尔克克森检测P<0.05);威海和潍坊种群2012、2014和2015年在3个模型下都表现显著的杂合度过剩,2013年在无限等位基因模型和双相突变模型下表现显著的杂合度过剩;烟台种群2012和2015年在3个模型下都表现显著的杂合度过剩;青岛种群2015年在3个模型下均表现显著的杂合度过剩, 2012—2014年在无限等位基因模型和双相突变模型下均表现显著的杂合度过剩;聊城种群2014年在3个模型下均表现显著的杂合度过剩, 2012年在无限等位基因模型和双相突变模型下表现显著的杂合度过剩,2013年在无限等位基因模型下表现显著的杂合度过剩。总之,2012—2015年苹果绵蚜6个地理种群均经历了不同程度的瓶颈效应。
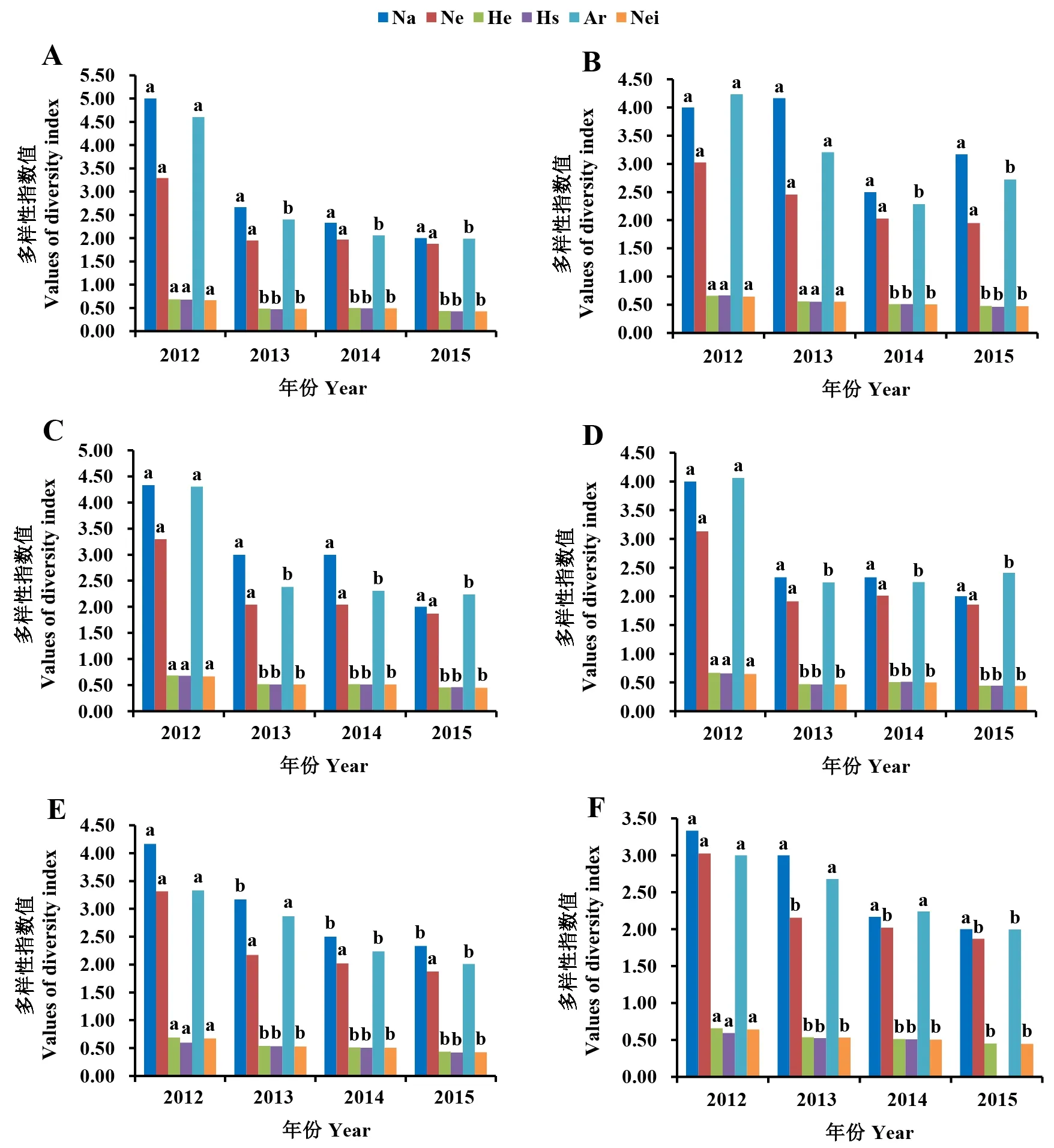
A:泰安市;B:聊城市;C:威海市;D:青岛市;E:潍坊市;F:烟台市。Na:等位基因观测值;Ne:等位基因期望值;He:期望杂合度;Hs:基因多样性;Ar:等位基因丰富度;Nei:Nei′s 期望杂合度。同种颜色柱上不同字母表示不同年份间差异显著(P<0.05)。A: Taian City; B: Liaocheng City; C: Weihai City; D: Qingdao City; E: Weifang City; F: Yantai City. Na: Number of alleles observed; Ne: Number of effective alleles; He: Expected heterozygosity; Hs: Heterogeneous stock; Ar: Allelic richness; Nei: Nei′s expected heterozygosity. Different letters on the column with the same color indicate significant differences among different years.图1 山东省2012—2015年6个地区苹果绵蚜种群多样性指数 Fig.1 The genetic diversity of E. lanigerum in six areas of Shandong Province during 2012-2015
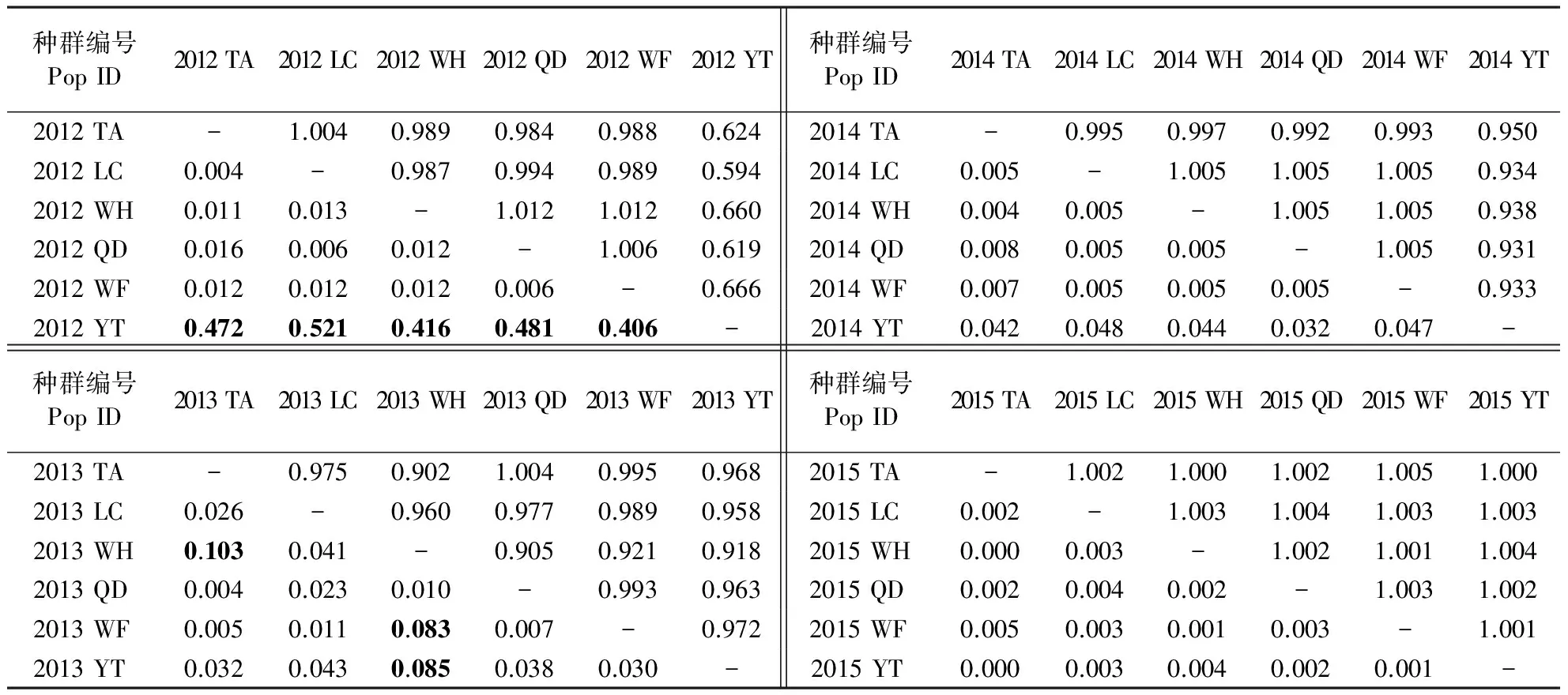
表2 2012—2015年山东省6个地区苹果绵蚜成对遗传分化指数FST(对角线下方)与基因流情况Nm(对角线上方)Table 2 Pairwise FST matrix (below diagonal) and gene flow Nm (above diagonal) of E. lanigerum population in six areas of Shandong Province during 2012-2015
TA:泰安市;LC:聊城市;WH:威海市;QD:青岛市;WF:潍坊市;YT:烟台市。粗体的数字表示差异显著(P<0.05)。基因流根据公式Nm=(1/FST-1)/2计算得出。
TA: Taian City; LC: Liaocheng City; WH: Weihai City; QD: Qingdao City; WF: Weifang City; YT: Yantai City. Bold numbers indicate significant differences (P<0.05). Gene flow is calculated according to formulaNm=(1/FST-1)/2.
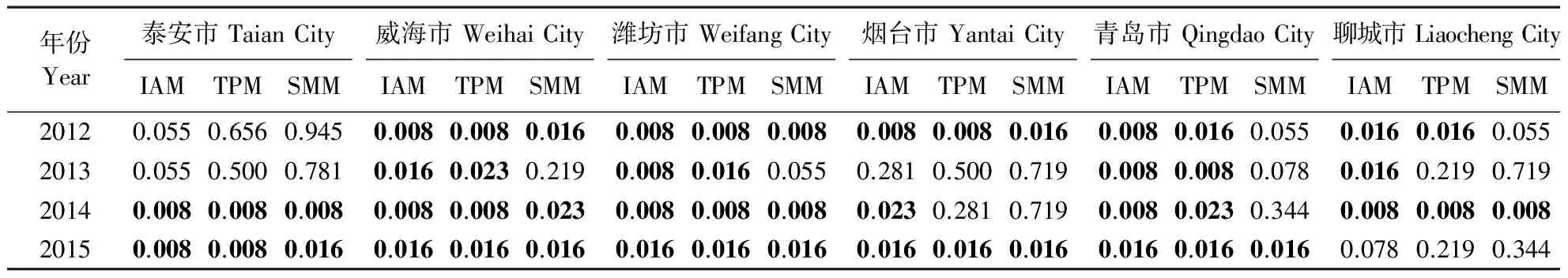
表3 采用IAM、TPM和SMM 3个模型对种群内杂合度过剩的检测Table 3 Within-population tests for heterozygosity excess according to the IAM, TPM and SMM models
IAM:无限等位基因模型;TPM:双相突变模型;SMM:逐步突变模型。粗体的数字表示杂合度过剩显著(P<0.05)。
IAM: Infinite allele model; TPM: Two-phased model of mutation; SMM: Stepwise mutation model. Bold numbers indicate significant heterozygosity excess (P<0.05).
如图2所示,当K=4时,ΔK为最大值,在STRUCTURE软件聚类分析中,最佳分组数K=4。通过STRUCTURE软件分析表明, 2012年6个地区主要为红色和绿色,应聚为一类,2013—2015年6个地区比较相近,主要为蓝色和黄色,应聚为一类(图3),与遗传多样性分析结果一致。
对6个地区苹果绵蚜不同位点不同年份等位基因的遗传多样性变化进行显著性分析(表4),发现E20、E75和E78 3个位点变化明显。2012—2015年E20位点的杂合度水平随时间推移表现降低的趋势。在P<0.05水平上,E20扩增的等位基因中,2012年的Ar、He和Nei均显著高于2013—2015年,且2013和2014年的值显著高于2015年。2012年E75和E78的Ar、He和Nei也均显著高于2013—2015年,2013—2015年之间的差异均不显著。总体而言,2012—2015年苹果绵蚜在E20、E75和E78 3个位点的遗传多样性随时间推移表现逐渐降低的趋势。
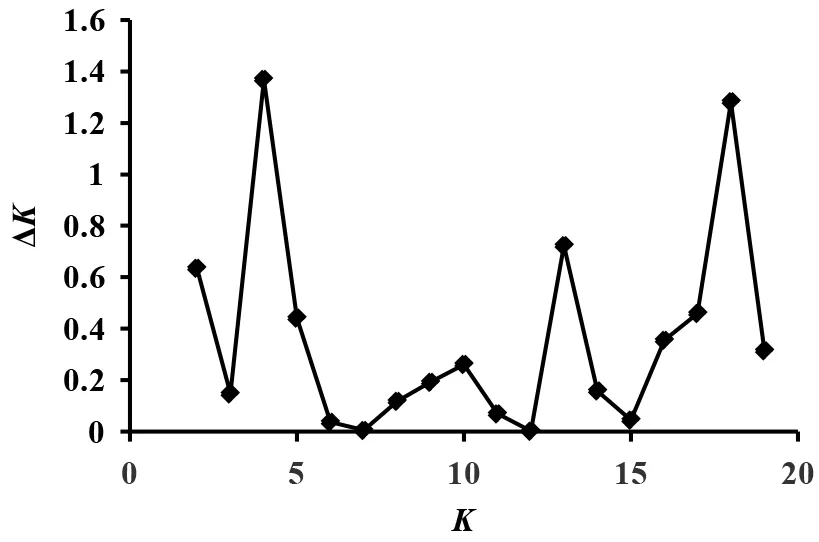
图2 ΔK值的散布图Fig.2 Scatter plots of ΔK
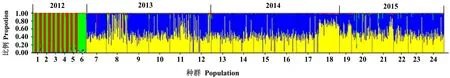
1、7、13、19:泰安市;2、8、14、20:聊城市;3、9、15、21:威海市;4、10、16、22:青岛市;5、11、17、23:潍坊市;6、12、18、24:烟台市。 1,7,13,19: Taian City; 2,8,14,20: Liaocheng City; 3,9,15,21: Weihai City; 4,10,16,22: Qingdao City; 5,11,17,23: Weifang City; 6,12,18,24: Yantai City.图3 基于STRUCTURE软件的2012—2015年山东省6个地区苹果绵蚜种群的贝叶斯聚类分析(K=4)Fig.3 Bayesian clustering analysis of E. lanigerum populations during 2012-2015 using STRUCTURE
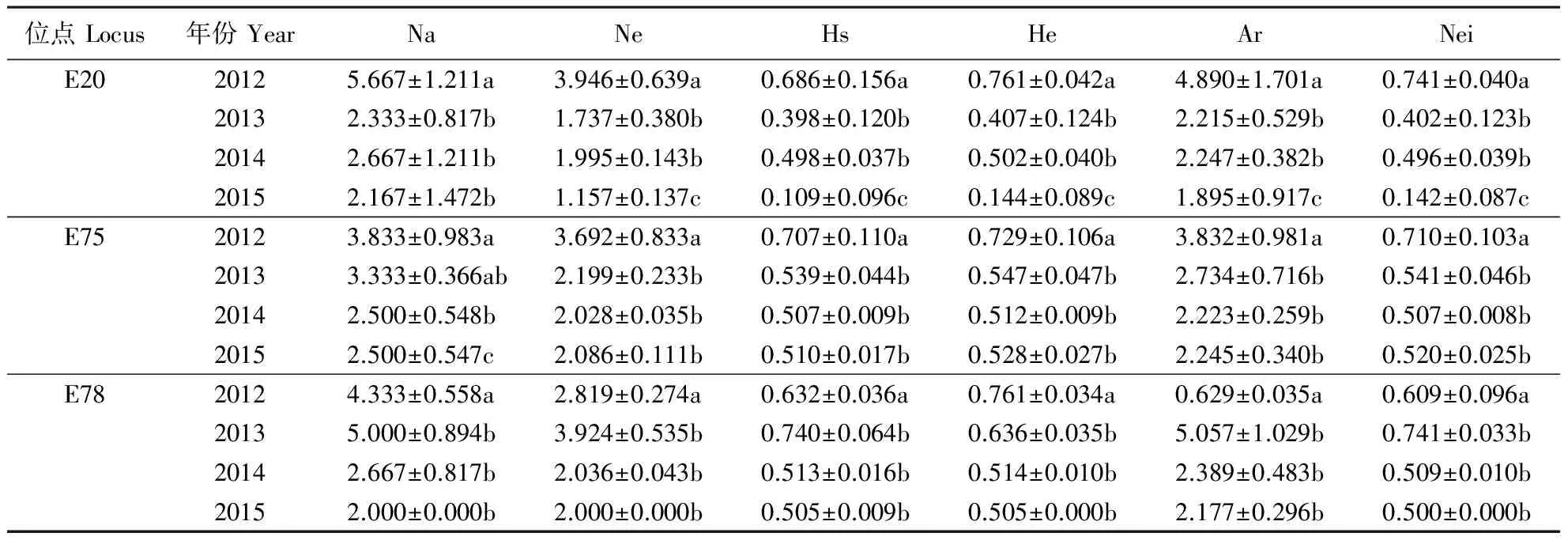
表4 2012—2015年苹果绵蚜不同位点的遗传多样性Table 4 The genetic diversity of E. lanigerum using different loci during 2012-2015
Na:等位基因观测值;Ne:等位基因期望值;Hs:基因多样性;He:期望杂合度;Ar:等位基因丰富度;Nei:Nei′s 期望杂合度。同列数据后不同字母表示同个位点在不同年份间差异显著(P<0.05)。
Na: Number of alleles observed; Ne: Number of effective alleles; Hs: Heterogeneous stock; He: Expected heterozygosity; Ar: Allelic richness; Nei: Nei′s expected heterozygosity. Different letters in the same column indicate significant differences among different years at the same locus (P<0.05).
3 讨论
苹果绵蚜对环境表现出广泛适应性,在传入后经历了严重的瓶颈效应,进化出“超级克隆(super clone)”种群(Zhouetal.,2015)。本文通过分析苹果绵蚜4年间的遗传多样性,发现2012—2015年6个地区苹果绵蚜种群的遗传多样性有降低趋势,且均经历了瓶颈效应。遗传多样性降低不利于生物入侵(Figueroaetal.,2005; Tsutsuietal.,2000)。Tsutsuietal.(2000)通过微卫星对阿根廷蚂蚁Linepithemahumile(Mayr)的遗传变异进行研究,发现阿根廷蚂蚁经过瓶颈效应后能成功入侵新的领地。Figueroaetal.(2005)对入侵智利的麦长管蚜Sitobionavenae(Fabricius)种群研究发现,入侵智利的麦长管蚜种群遗传多样性降低,可能是种群在初传入后经历了严重瓶颈效应,进化出“超级克隆”基因型,增强了对环境的适应能力。因此,笔者认为,苹果绵蚜在我国可能进化出了“超级克隆”基因型。Zhouetal.(2015)通过微卫星研究发现,我国北方8个省份主要苹果种植区的苹果绵蚜种群形成一个不同于云南和新疆的单一的“supercolony”。 因此,苹果绵蚜传入我国一个世纪以来,经过长期的环境适应,为增强其生态适应能力,出现了超级克隆大种群,这可能是近十几年来苹果绵蚜再次大暴发并扩散蔓延的原因之一。
苹果绵蚜遗传多样性的降低与E20、E75和E78 3个位点的变化有关。在遗传距离的分析中,同一年份中6个地区种群的遗传距离差异均不显著。E20、E75和E78 3个位点遗传多样性变化规律与苹果绵蚜遗传多样性随时间的变化规律一致:2012年E20、 E75和E78 3个位点的遗传多样性指数显著高于2013—2015年,而其他3个位点(E3、E25和E72)的遗传多样性指数随时间变化没有明显增减趋势。因此,在苹果绵蚜遗传多样性变化中,E20、E75和E78 3个位点起关键性作用,可能与这3个位点的近缘连锁基因有关,这需要进一步研究。
此外,日光蜂是苹果绵蚜的重要内寄生蜂(周洪旭等,2010)。基于微卫星标记研究苹果绵蚜和日光蜂的不同地理种群,发现苹果绵蚜入侵途径与日光蜂扩散途径基本一致(Zhouetal.,2014a,2014b),二者之间可能存在一定的协同进化关系。因此认为,苹果绵蚜遗传多样性的降低还可能与其专性寄生蜂日光蜂田间寄生作用有关,这种内在关系尚需进一步研究。
董锦花, 程斌, 陆俊姣, 马瑞燕, 2008. 检疫性害虫苹果绵蚜在山西发生危害的调查与研究. 山西大学学报(自然科学版), 28(2): 156-158.
段云, 2013. 中国麦红吸浆虫不同地理种群的遗传结构及遗传多样性研究. 博士学位论文. 北京: 中国农业科学院.
郭小侠, 崔俊锋, 陈川, 石勇强, 2011. 苹果绵蚜风险分析和风险管理. 北方园艺 (13): 135-136.
贺春玲, 田海燕, 毛永珍, 2004. 我国苹果绵蚜发生及防治研究进展. 陕西林业科技 (1): 34-38.
李定旭, 陈根强, 李文亮, 张志勇, 司雨, 2003. 河南省苹果绵蚜发生现状及其防治对策. 植物检疫, 17(3): 149-151.王朴, 刘霞, 唐永清, 王佳武, 薛海滨, 张建军, 2007. 伊犁苹果绵蚜的发生与防治. 新疆农垦科技 (1): 29.
王西存, 周洪旭, 郭建英, 万方浩, 于毅, 2011. 苹果绵蚜在不同寄主果园中的种群数量动态比较. 生物安全学报, 20(3): 220-226.
王兴亚, 蒋春廷, 许国庆, 2011. 外来入侵种——苹果绵蚜在中国的适生区预测. 应用昆虫学报, 48(2): 379-391.
吴海军, 李友莲, 丁三寅, 李惠萍, 2007. 入侵生物苹果绵蚜在中国的风险性分析. 山西农业大学学报(自然科学版), 27(4): 368-371.
于江南, 陈卫民, 徐毅, 柳晓玲, 2008. 伊犁河谷苹果绵蚜生物学特性及防治. 新疆农业科学, 45(2): 298-301.
张福芹, 2002. 苹果绵蚜的综合防治技术. 河北林业科技 (4): 27-28.
赵增锋, 2012. 苹果病虫害种类、地域分布及主要病虫害发生趋势研究. 博士学位论文. 保定: 河北农业大学.周洪旭, 郭建英, 万方浩, 于毅, 2010. 日光蜂对苹果绵蚜的自然控制作用及其保护和利用. 植物保护学报, 37(2): 153-158.ATEYYAT M A, ALANTARY T M, 2009. Susceptibility of nine apple cultivars to woolly apple aphid,Eriosomalanigerum(Homoptera: Aphididae) in Jordan.InternationalJournalofPestManagement, 55(1): 79-84.
ATEYYAT M A, ALANTARY T M, 2010. Population trends of woolly apple aphid,Eriosomalanigerumand its parasitoid,Aphelinusmalion two apple cultivars in Jordan.JordanJournalofAgriculturalSciences, 6(3): 333-341.
CORNUET J M, LUIKART G, 1996. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data.Genetics, 144: 2001-2014.EVANNO G, REGNAUT S, GOUDET J, 2005. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study.MolecularEcology, 14: 2611-2620.FIGUEROA C C, SIMON J C, LE GALLIC J F, PRUNIER-LETERME N, BRIONES L M, DEDRYVER C A, NIEMEYER H M, 2005. Genetic structure and clonal diversity of an introduced pest in Chile, the cereal aphidSitobionavenae.Heredity, 95(1): 24-33.
FRANKS S J, PRATT P D, TSUTSUI N D, 2010. The genetic consequences of a demographic bottleneck in an introduced biological control insect.ConservationGenetics, 12: 201-211.HAVILL N P, MONTGOMERY M E, YU G, SHIYAKE S, CACCONE A, 2006. Mitochondrial DNA from hemlock woolly adelgid (Hemiptera: Adelgidae) suggests cryptic speciation and pinpoints the source of the introduction to eastern North America.AnnalsoftheEntomologicalSocietyofAmerica, 99: 195-203.
KELM M, LENORT M, LICZNAR M, 2004. Occurrence of woolly aphidEriosomalanigerumHausm. in different systems of apple orchards.ProgressinPlantProtection, 44(2): 809-811.LAVANDERO B, FIGUEROA C C, RAMIREZ C C, CALIGARI P D S, FUENTES-CONTRERAS E, 2009. Isolation and characterization of microsatellite loci from the woolly apple aphidEriosomalanigerum(Hemiptera: Aphididae: Eriosomatinae).MolecularEcologyResources, 9(1): 302-304.NICHOLAS A H, SPOONER-HART R N, VICKERS R A, 2005. Abundance and natural control of the woolly aphidEriosomalanigerumin an Australian apple orchard IPM program.BioControl, 50(2): 271-291.
PRITCHARD J K, STEPHENS M, DONNELLY P, 2000. Inference of population structure using multilocus genotype data.Genetics, 155: 945-959.
SUNNUCKS P, HALES D, 1996. Numerous transposed sequences of mitochondrial cytochrome oxidase I-II in aphids of the genusSitobion(Hemiptera: Aphididae).MolecularBiologyandEvolution, 13: 510-524.
TIMM A E, PRINGLE K L, WARNICH L, 2005. Genetic diversity of woolly apple aphidEriosomalanigerum(Hemiptera: Aphididae) populations in the Western Cape, South Africa.BulletinofEntomologicalResearch, 95(3): 187-191.
TSUTSUI N D, SUAREZ A V, HOLWAY D A, CASE T J, 2000. Reduced genetic variation and the success of an invasive species.ProceedingsoftheNationalAcademyofSciences, 97(11): 5948-5953. WEIR B S, COCKERHAM C C, 1984. Estimating F-statistics for the analysis of population structure.Evolution, 38: 1358-1370.YEH F C, YANG R C, BOYLE T, YE Z H, MAO J X, 1999.POPGENE,theuser-friendlysharewareforpopulationgeneticanalysis. Edmonton, Canada: Molecular Biology and Biotechnology Centre, University of Alberta.
ZHOU H X, WANG X C, YU Y, 2013. Chemical characteristics of normal, woolly apple aphid-damaged, and mechanically damaged twigs of six apple cultivars, measured in autumn wood.JournalofEconomicEntomology, 106(2): 1011-1017.
ZHOU H X, ZHANG R M, GUO D, 2014a. Two putative bridgehead populations ofAphelinusmali(Hymenoptera: Aphelinidae) introduced in China as revealed by mitochondrial DNA marker.FloridaEntomologist, 97(2): 401-405.
ZHOU H X, ZHANG R M, GUO D, 2014b. Analysis of genetic diversity and structure of two clades ofAphelinusmali(Hymenoptera: Aphelinidae) in China.FloridaEntomologist, 97(2): 699-706.
ZHOU H X, ZHANG R M, TAN X M, TAO Y L, WAN F H, WU Q, CHU D, 2015. Invasion genetics of woolly apple aphid (Hemiptera: Aphididae) in China.JournalofEconomicEntomology, 108(3): 1040-1046.
(责任编辑:杨郁霞)
The genetic structure dynamics ofEriosomalanigerum(Hausmann) based on microsatellite markers
YANG Mingming1, TAN Xiumei1, WEI Chengmin1, ZHANG Yongxiao1, WAN Fanghao1,2, ZHOU Hongxu1*
1KeyLabofIntegratedCropPestManagementofShandongProvince,CollegeofAgronomyandPlantProtection,QingdaoAgriculturalUniversity,Qingdao,Shandong266109;2StateKeyLaboratoryforBiologyofPlantDiseasesandInsectPests,InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China
【Aim】Eriosomalanigerum, an important quarantine pest in the world, could cause serious harm to the apple tree. In recent years,E.lanigerumspread far in China and caused serious damage. Studying the molecular ecological changes during its invasion can provide basis for comprehensive control. 【Method】 We made six microsatellite markers to explore the genetic diversity dynamics ofE.lanigerumfrom six areas (Yantai, Weihai, Qingdao, Weifang, Liaocheng, Taian) in Shandong Province from 2012 to 2015. 【Result】 The genetic diversity ofE.lanigerumgradually reduced over time in all six areas. The genetic diversity in 2012 was significantly higher than in later years, and no significant differences were found from 2013 to 2015. Genetic diversity expressed as the value of gene alleles observed (Na) and expected heterozygosity (He) had a tendency to gradually reduce over time.E.lanigerumin all six areas went through the bottleneck effect, detected by analysis of IAM (infinite allele model), TPM (two-phased model of mutation) and SMM (stepwise mutation model), which mainly caused reduction of genetic diversity. The number of alleles and the genetic diversity index in microsatellite loci Erio20, Erio75 and Erio78 decreased over time. 【Conclusion】 The locus of Erio20, Erio75 and Erio78 are the main loci to cause the decrease of genetic diversity ofE.lanigerum.E.lanigerummay have evolved a "super clone" genotype to adapt to different environments in China and increase the diffusion range.
Eriosomalanigerum; invasive pest; microsatellite; genetic diversity; dynamics
2016-09-23 接受日期(Accepted): 2016-11-21
国家自然科学基金项目(31371994); 国家重点研发计划(2016YFC1201200); 国家重点基础研究发展计划(“973”计划) (2013CB127600); 山东省“泰山学者”建设工程专项经费资助
杨明明, 女, 硕士研究生。 研究方向: 农业昆虫与害虫防治。 E-mail: 996251660@qq.com
*通信作者(Author for correspondence), E-mail: hxzhou@qau.edu.cn
10. 3969/j.issn.2095-1787.2017.02.003
猜你喜欢
特产研究(2022年6期)2023-01-17 05:05:06
智慧健康(2021年17期)2021-07-30 14:38:32
种子(2021年3期)2021-04-12 01:42:22
四川动物(2017年4期)2017-07-31 23:54:19
现代检验医学杂志(2016年5期)2016-08-20 03:16:54
外语教学理论与实践(2016年1期)2016-06-11 05:51:48
法医学杂志(2015年4期)2016-01-06 12:36:40
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:37
河北遥感(2014年3期)2014-07-10 13:16:48
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:29
