
优质桔红心大白菜种质创新与新品种选育
2017-06-05 02:52张一卉李化银王凤德李景娟丁谦刘立锋孙令强徐少君孔涛王翠花何启伟高建伟��
山东农业科学 2017年5期
张一卉 李化银+王凤德 李景娟 丁谦 刘立锋+孙令强 徐少君+孔涛+王翠花 何启伟 高建伟 ��

摘要:本試验利用分子育种手段开展了优质桔红心大白菜(Brassica rapa L. ssp. pekinensis)种质创新与新品种选育研究。我们克隆了控制大白菜桔红心的关键基因BrCRTISO,分析了桔红心、白心大白菜之间BrCRTISO基因的结构差异。根据启动子和编码区序列的差异,筛选出了与桔红心紧密连锁的SNP和InDel分子标记。在传统有性杂交的基础上,利用上述SNP和InDel标记对后代分离群体进行辅助选择,筛选出了早熟、优质、抗病的桔红心优异纯系种质08428、08460、08468、08469自交不亲和系。利用上述4个纯系配置组合,选育出了早熟优质抗病的特色大白菜新品种“天正桔红65”、“天正桔红62”。连续多年(2010—2016)种植试验及抗病性鉴定的结果表明:这两个新品种抗病性强,适宜春秋种植,早熟(55~60天);叶球中小型,桔红色。其中“天正桔红65”β-胡萝卜素含量高达26.163 mg/gDW,是普通大白菜的11.15倍;可溶性固形物含量高,口感品质极佳。2014年,“天正桔红62”成为泰安市岱岳区地理标志性产品(商标注册号:13066669)。2015—2016年,两个品种通过了国家、省(市)农作物品种审定委员会的审定(鉴定)。本研究是利用分子育种手段快速培育优良大白菜新品种的成功范例,可为其它生物分子育种工作提供参考。
关键词:大白菜;桔红心;分子标记;种质创新;新品种选育
中图分类号:S634.103.2文献标识号:A文章编号:1001-4942(2017)05-0014-10
Germplasm Innovation and New Variety Breeding in Orange
Leafy Head Chinese Cabbage (Brassica rapa L. ssp. pekinensis)
Zhang Yihui1, Li Huayin1*, Wang Fengde1, Li Jingjuan1, Ding Qian1, Liu Lifeng1,
Sun Lingqiang2, Xu Shaojun3, Kong Tao4, Wang Cuihua1, He Qiwei1, Gao Jianwei1
(1. Institute of Vegetables and Flowers, Shandong Academy of Agricultural Sciences, Jinan 250100, China;
2.Qingdao Seed Management Station, Qingdao 266071, China; 3. Qingdao Hefeng Seeds Co., Ltd.,
Jiaozhou 266300, China;4. Daiyue District Agricultural Bureau, Taian 271021, China)
AbstractIn this research, the molecular breeding was used to innovate germplasm and select new varieties in orange leafy head Chinese cabbage (Brassica rapa L. ssp. pekinensis). We cloned the candidate key gene BrCRTISO, which coded carotenoid isomerase controlling synthesis of orange pigments in leafy head. Differences in structure of BrCRTISO gene between orange and white leafy head Chinese cabbage were also analyzed, in which, one SNP(C952-T952)marker and one InDel marker Bror-intron1 closely linked with orange leafy head were developed. New good pure self-incompatible lines of orange leafy head germplasm had been innovated by molecular marker assisted selection, of which, 08428, 08460, 08468 and 08469 were better than others. These four lines were applied to make cross combinations, from which, new orange leafy head varieties Tianzhengjuhong 65 and Tianzhengjuhong 62 were obtained. Planting and disease resistance tests for several years (2010-2016) showed that these two new varieties had strong disease resistance, and their growth periods were short (55~60 days), which were suitable for being cultivated in spring or autumn. The β-carotene content of Tianzhengjuhong 65 reached 26.163 mg/gDW, 11.15 times of common Chinese cabbage. In addition, the content of soluble solids was also higher than other Chinese cabbage varieties, which owned good taste. In 2014, Tianzhengjuhong 62 was approved to be Chinese National Protected Geographic Indication Product (No. 13066669 ) in Taian, China. In 2015, Tianzhengjuhong 65 was approved by both National and Beijing Crop Variety Approval Committee. Meanwhile, Tianzhengjuhong 62 was approved by Beijing Crop Variety Approval Committee. In 2016, Tianzhengjuhong 65 was approved by Shandong Crop Variety Approval Committee. This paper would be useful to other plant molecular breeding because of our successful breeding work.
KeywordsChinese cabbage; Orange leafy head; Molecular markers; Germplasm innovation; New variety breeding
大白菜(Brassica rapa L. ssp. pekinensis)起源于中国,是我国的特产蔬菜。韩国、日本等东亚国家种植的大白菜最早由我国传入。长期以来,大白菜一直是我国和东亚国家的骨干蔬菜,其重要地位和生产面积是其它蔬菜难以相比的。在我国北方,大白菜是秋季栽培和冬春供应的主要蔬菜,种植面积大、范围广[1]。山东省作为大白菜的主要起源地之一,栽培历史悠久,种质资源极为丰富,其中有誉满海内外的胶东大白菜,有特点鲜明的德州香把子,有深受济南本地人喜爱的唐王大白菜等[2]。随着我国农业产业结构的调整和保护地栽培蔬菜的发展,大白菜一季生产半年供应的格局已发生了显著变化。随着人们生活水平的不断提高,人们将更多地关注大白菜的质量,即形状、大小、色泽、营养、风味口感及无公害等指标。
桔红心大白菜因其色泽艳丽、富含胡萝卜素等多种维生素、营养价值高、口感好而倍受消费者青睐。日本桔红心大白菜育种起步较早,培育了许多优良品种。但是在我国,日本的桔红心大白菜品种种子昂贵,不抗病毒病,病重地区和年份常常绝产,难以获得丰产和稳产。因此,培育适宜山东等地种植的高抗病毒病、霜霉病、软腐病的桔红心大白菜新品种,实现其种子国产化显得尤为重要[3]。
针对上述问题,本课题组自2002年起开始收集、鉴定桔红心大白菜资源材料,研究桔红心性状遗传特性。2007—2010年间,我们通过拓宽育种资源,利用杂交组合选配测配,结合分子标记辅助选择技术,选育出适合春秋两季种植的桔红心大白菜新品种“天正桔红65”、“天正桔红62”。2014年,“天正桔红62”成为泰安市岱岳区地理标志性产品(商标注册号:13066669)。2015年,两品种通过了国家、北京市农作物品种审定委员会的审定(鉴定);2016年,“天正桔红65”又通过了山东省农作物品种审定委员会的审定。
1材料与方法
1.1试验材料
桔红心大白菜一代杂交种:北京桔红心、红抗1号、长研橘宝、申萌桔红心、神狮桔红心。桔红心大白菜纯系:桔08410、14-490、1480、14-102、14-245、14-253、14-257、14-277、14-426、14-662、14-669。白心(或者黄心)大白菜一代杂交种:寒秀、金典春王、凯春、日本夏阳。白心(或者黄心)大白菜纯系:新福08410、14-401、663、1466、1469、1505、1510、1720、叠锥抗2。杂交组合(14-401×14-490)及其F2代分离群体。
1.2基因克隆及生物信息学分析
1.2.1基因组DNA提取取大白菜叶片,液氮冷冻后快速研磨,采用快捷型植物基因组DNA提取系统(天根生化科技有限公司)提取基因组DNA,利用1%琼脂糖凝胶电泳进行DNA检测。
1.2.2总RNA提取和反转录取结球后期的球叶提取总RNA。采用TRNzol-A+试剂盒(天根生化科技有限公司)进行RNA提取,方法参考说明书。反转录按照TaKaRa PrimeScript RT reagent Kit with gDNA Eraser (大连宝生物)产品说明书操作。
1.2.3PCR扩增PCR反应体系(20 μL)为:2×高保真PCR Mix 10 μL,上游引物(10 mmol/L)0.5 μL,下游引物(10 mmol/L)0.5 μL,模板cDNA 1 μL,ddH2O 8 μL。PCR反应程序为:94℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 1 min,36个循环;72℃ 10 min。
1.2.4SDS-PAGE电泳检测将PCR产物与上样缓冲液混匀,然后将混合物95℃预变性10 min。取4 μL点样,利用8%变性聚丙烯酰氨凝胶进行电泳检测,在Sequi-Gen GT核酸电泳系统(Bio-Rad,USA)中进行电泳分离,电极缓冲液为1×TBE,在25℃、55 W恒功率下电泳1 h,通过银染法显影、拍照并记录结果。
1.2.5生物信息学分析将克隆的基因序列在GenBank中进行BLAST检索,查找大白菜目标基因在其它物种中的同源序列。利用DNAMAN 6.0软件对来自不同物种的基因编码的氨基酸序列进行比对并构建系统进化树。
1.3β-胡萝卜素等营养成分的测定
利用高效液相色谱法测定大白菜β-胡萝卜素含量[5-7]。称取5.0 g左右干净的成熟期大白菜内叶,剪碎,置于三角瓶中,用石油醚∶丙酮(体积比为80∶20)混合液振摇提取,移出上清液,重复提取直至提取液无色,合并提取液,转入圆底烧瓶中,于旋转蒸发仪上蒸发至干,用二氯甲烷溶液提取残渣,过滤后用凝胶色谱仪净化。净化后的溶液用于HPLC分析。使用吉尔森高效液相色谱仪,色柱:C18柱,250 mm×4.6 mm,5 μm,流动相为甲醇∶乙腈(体积比为90∶10),流速为1.0 mL/min;检测波长为450 μm,进样量20 μL。利用PAD在波长200~800 nm范围内扫描,分析物质的特征吸收峰。
可溶性固形物含量的测定参照标准《NY/T 2637—2014 水果和蔬菜可溶性固形物含量的测定 折射仪法(2015-1-1实施)》。
1.4分子标记辅助选择及常规杂交育种
分子标记辅助选择育种及常规杂交育种参照钱前等(2012)[4]和柯桂兰(2010)[1]。
2结果与分析
2.1控制大白菜桔红心形成的关键候选基因的选择策略
与普通大白菜相比,桔红心大白菜的叶球内叶呈现桔红色。切开后剖面经阳光照射,球叶的桔红色快速增强。以往研究表明,桔红心大白菜内叶含有类似番茄红素的复合物等多种类胡萝卜素[5-7]。类胡萝卜素在植物体内通过类异戊二烯途径合成,呈现黄色、橙红色和红色。在拟南芥[6]、番茄[7]等模式植物中,人们对类胡萝卜素生物合成途径及其相关酶(或基因)的功能研究得很清楚[6-8]。植物中,许多桔红色性状的出现都与类胡萝卜素异构酶有关。类胡萝卜素异构酶CRTISO 能催化前番茄红素形成全反式番茄红素[9,10]。Park 等(2002)[6]在拟南芥中分离了CRTISO基因,通过分析其功能,发现类胡萝卜素异构酶缺失突变体主要積累前番茄红素和前链孢红素(7, 9, 9′–三顺式–链孢红素)。Isaacson 等(2002)[7]利用图位克隆技术获得了番茄桔红色基因tangerine,该基因编码一个类胡萝卜素异构酶CRTISO,tangerine突变体缺失CRTISO 导致番茄积累前番茄红素和β-胡萝卜素,果实呈现桔红色。因此,在筛选控制大白菜桔红心形成的关键候选基因时,我们首先选择了编码类胡萝卜素异构酶的基因BrCRTISO作为候选基因。
另外,与白心大白菜相比,桔红心大白菜叶球被剖开的瞬间呈现黄色。我们在考虑筛选控制大白菜桔红心形成的关键候选基因时,也应该考虑控制黄心形成的关键基因。Galpaz等(2006)[8]在番茄中克隆了控制白花形成的基因CrtR-b,它是由于编码β-胡萝卜素羟化酶的基因发生突变引起的。在大白菜中,存在3个CrtR-b成员,分别为BrCrtR-b1、BrCrtR-b2、BrCrtR-b3。综合考虑类胡萝卜素代谢途径,在筛选大白菜桔红心关键候选基因时,我们又选择了BrCrtR-b1、BrCrtR-b2、BrCrtR-b3基因作为候选基因。
2.2控制大白菜桔红心形成的关键基因及分子标记的挖掘
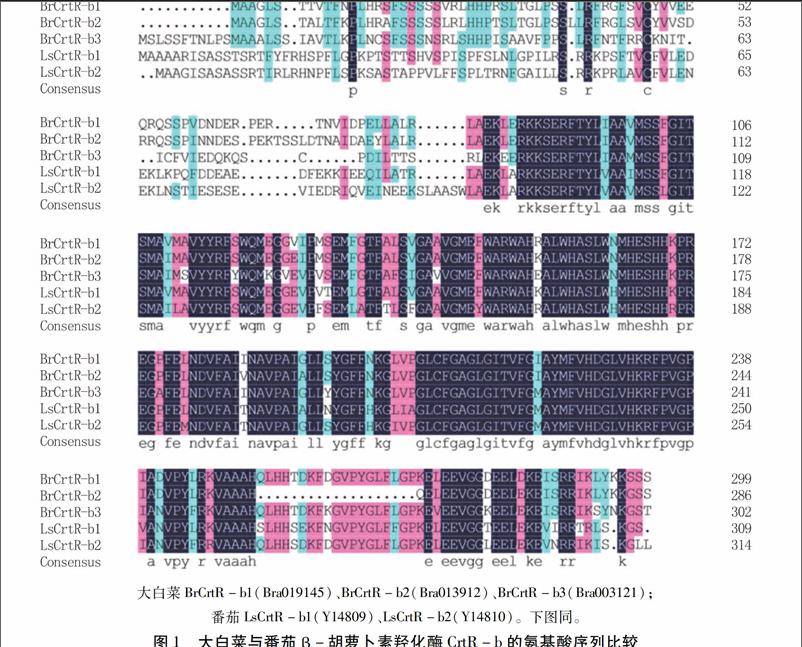
2.2.1大白菜BrCrtR-b1、BrCrtR-b2、BrCrtR-b3不是控制叶球桔红心形成的关键基因2007年,我们利用番茄LsCrtR-b1(Accession No. Y14809)和LsCrtR-b2(Accession No. Y14810)基因序列及其编码的氨基酸序列在GenBank (https://www.ncbi.nlm.nih.gov/) 中搜寻大白菜对应的EST、基因组序列,发现大白菜中存在3个CrtR-b成员(BrCrtR-b1、BrCrtR-b2、BrCrtR-b3)。根据上述信息,我们设计了扩增BrCrtR-b1、BrCrtR-b2、BrCrtR-b3基因cDNA序列的引物。利用桔红心大白菜纯系亲本“桔08410”和白心大白菜纯系“新福08410”结球后期的心叶作为供试材料,扩增出了BrCrtR-b1、BrCrtR-b2、BrCrtR-b3基因的cDNA序列。而后,又进一步扩增这3个基因的基因组序列。结果显示,BrCrtR-b1、BrCrtR-b2、BrCrtR-b3基因的cDNA、基因组序列在桔红心大白菜纯系亲本“桔08410”和白心大白菜纯系“新福08410”之间无差异。
我们获得的BrCrtR-b1、BrCrtR-b2、BrCrtR-b3基因的cDNA、基因组序列与2011年公布的大白菜全基因组序列[11]结果相同。我们采用Wang等(2011)[11]发布的BrCrtR-b1、BrCrtR-b2、BrCrtR-b3基因及其蛋白信息,对大白菜BrCrtR-b与番茄LsCrtR-b的氨基酸序列进行了比对分析(图1、2)。结果表明,在大白菜和番茄之间,CrtR-b氨基酸序列的保守性较强,一致性为75.11%。
2.2.2大白菜BrCRTISO是控制叶球桔红心形成的关键基因2007年,我们利用拟南芥AtCRTISO(Accession No. AAF63149)基因序列及其编码的氨基酸序列在GenBank(https://www.ncbi.nlm.nih.gov/)中搜寻,获得了对应的大白菜ESTs(Accession No. EX023490、EX043675、EX040997、EX056774)等序列信息。根据这些序列信息,我们拼接出了大白菜BrCRTISO的全长
cDNA序列,而后克隆了BrCRTISO基因全长cDNA。利用DNAMAN软件对大白菜(采用编号Bra031539)[11]、拟南芥、番茄、水稻和矮牵牛的CRTISO进行了比较分析(图3、4)。结果表明,BrCRTISO与AtCRTISO同源性高,氨基酸序列的一致性高达88.63%;BrCRTISO与OsCRTISO同源性低,氨基酸序列的一致性为70.65%。
利用桔红心大白菜纯系亲本“桔08410”和白心大白菜纯系“新福08410”结球后期的心叶作为
供试材料,扩增出了BrCRISO基因的基因组序列及编码序列。结果显示,BrCRISO基因的基因组序列及编码序列在“桔08410”和“新福08410”之间有明显差异。进一步的群体验证结果表明,这种差异与叶球桔红心性状共分离。因此,我们推测大白菜BrCRTISO是控制桔红心形成的关键基因。
2.2.3与大白菜叶球桔红心紧密连锁的SNP和InDel分子标记通过我们以及其他不同课题组对于白心和桔红心大白菜BrCRTISO基因序列的分析,发现两种大白菜之间存在较大的序列差异,包括大量的SNPs和InDels。在大量的SNPs中,有7个SNPs与桔红心性状完全连锁,但其中仅有一个SNP(C952-T952)造成了氨基酸的突变(L318-F318)(图5)。进一步利用白心和桔红心的杂交F2代群体、自然群体以及部分栽培品种进行检測,发现SNP952与桔红心性状完全连锁[12]。因此,我们将SNP952作为分子标记应用于桔红心大白菜种质资源的筛选和新品种的培育中。
除了SNP952,位于第一个内含子上的一个InDel标记(Bror-intron1)也被我们广泛用于桔红心新品种的培育(图6)。该标记位于BrCRTISO基因的第一个内含子上,其中桔红心与白心相比存在25个碱基的插入。利用该标记可以快速、准确地鉴定不同大白菜种质资源是否带有桔红心性状基因,并可区分其是纯合体还是杂合体[12]。
2.3SNP和InDel分子标记辅助选择创制桔红心亲本纯系
引进的桔红心大白菜“红抗1号”杂交种抗霜霉病、软腐病,综合性状优良,但不抗病毒病。2007年,我们利用叠抱、高抗病毒病的白心大白菜纯系“叠锥抗2”作为病毒抗源供体回交两次,而后利用分子标记[桔红心标记:Bror-intron1、SNP(C952-T952);病毒病抗性标记BRRV:BrID10694、101309;耐热性标记:BRRH]辅助选择,进行抗性转育,再经多代自交分离、筛选,获得了新的桔红心育种纯系材料10份(08462、08463、08468、08469、08471、08472、08475、08477、08480、08481)。这10份纯系材料自交不亲和,田间表现高抗病毒病,结球紧实,株形和配合力好。其结球类型分别是:扣抱(叶色浅绿)、扣抱(叶色浓绿)、扣抱(叶色浓绿)、扣抱(叶色浅绿)、扣抱(叶色绿)、扣抱(叶色浅绿)、扣抱(叶色绿)、合抱(叶色浅绿)、扣抱(叶色浓绿)、扣抱(叶色浓绿)。
我们又引进了“北京桔红心”杂交一代,该品种高抗病毒病、霜霉病和软腐病。2007年,我们利用分子标记(软腐病抗性标记:BRS1;霜霉病抗性标记:BRDM2)辅助选择技术,对北京桔红心(杂交种F1)进行多代自交纯化,筛选出8个自交不亲和系(08427、08428、08431、08435、08436、08437、08451、08460)。这8个自交不亲和系,结球紧实,株型和经济性状优良,配合力好,且抗多种病害。其结球类型分别是:扣抱(叶色浓绿)、合抱(叶色浅绿)、合抱(叶色绿)、扣抱(叶色浅绿)、扣抱(叶色绿)、合抱(叶色浅绿)、扣抱(叶色浓绿)、扣抱(叶色浓绿)。
(a)为桔红心InDel标记Bror-intron1;14-401为白心大白菜纯系,14-490为桔红心大白菜纯系;
Bror-intron1-F/R为桔红心InDel标记Bror-intron1正/反向引物。
(b)用14-401×14-490及其F2代分离个体验证Bror-intron1标记。1—11为白心个体,12—19为桔红心个体。
(c)利用白心(或者黄心)、桔红心大白菜品种验证。1—11为白心(或者黄心)大白菜品种,12—23为桔红心大白菜品种[12]。
2.4桔红心大白菜新品种“天正桔红65”、“天正桔红62”的选育及其生物学特性
2010年,首次测配组合。测配结果表明,组合08428×08468(“天正桔红65”)、08469×08460(“天正桔红62”)表现优良。经2010、2011年春秋季田间品比试验、性状调查及抗病性鉴定,确定这两个优良组合为适宜春秋种植的桔红心特色品种。经高效液相色谱测定,“天正桔红65”β-胡萝卜素含量高达26.163 mg/gDW,是普通大白菜的11.15倍;可溶性固形物含量比北京新3号高,口感品质极佳;对病毒病、霜霉病及软腐病抗性强,结球紧实,采收期长。2014年,我们以“天正桔红62”注册了地理标志证明商标“泰安黄芽白菜”(商标注册号:13066669)。2015年,“天正桔红65”、“天正桔红62”通过了国家、北京市农作物品种审定委员会的审定(鉴定);2016年,“天正桔红65”又通过了山东省农作物品种审定委员会的审定。
2.4.1天正桔红65株高33 cm,合抱,生长期65天,单球重1.5~2.0 kg,净菜率71.1%。球叶颜色鲜艳,桔紅色深(图 7)。三年全国区域试验结果如下。2012年:秋中早熟品种,平均生长期为55~60天,品种纯度较高,叶球紧实;高抗病毒病、霜霉病、软腐病;净菜产量为70 080.0 kg/hm2,比对照增产13.8%,增产显著。2013年:秋中早熟品种,平均生长期为60天,品种纯度较高,叶球紧实;高抗病毒病、软腐病,抗霜霉病;净菜产量为60 396.0 kg/hm2,比对照增产8.6%,增产显著。2014年:秋中早熟品种,平均生长期为60天,品种纯度较高,叶球紧实;高抗病毒病、霜霉病、软腐病;净菜产量为65 934.0 kg/hm2,比对照增产9.3%,增产显著。
2.4.2天正桔红62株高42 cm,株型紧凑,扣抱,生长期55~60天,单球重2.5 kg左右,净菜率71.3%,橘红色深,口感品质极佳(图 7)。三年全国区域试验结果如下。2012年:秋中早熟品种,平均生长期为74.9天,品种纯度较高,叶球紧实;高抗病毒病、霜霉病及软腐病;净菜产量为64 605.0 kg/hm2,比对照增产4.9%。2013年:秋中早熟品种,平均生长期为73.9天,品种纯度较高,叶球紧实;高抗病毒病,抗霜霉病、软腐病;净菜产量为59 161.5 kg/hm2,比对照增产6.3%。2014年:秋中早熟品种,平均生长期为74.1天,品种纯度较高,叶球紧实;高抗病毒病、软腐病,抗霜霉病;净菜产量为66 682.5 kg/hm2,比对照增产10.5%。
2.4.3新品种天正桔红65、天正桔红62栽培技术要点天正桔红65、天正桔红62大白菜新品种属于早熟品种,与普通大白菜早熟品种栽培技术相同。春播,要求日气温不低于13℃。由于生长期短,秋季可根据上市时间分期播种,山东地区一般8月10号至9月2号均可播种,60 000~75 000株/hm2左右;若120 000株/hm2左右,可作为娃娃菜使用。其前茬应尽量避免十字花科作物,且有良好的水浇条件,地力肥沃,施足基肥,及时追肥,及时防治菜青虫、小菜蛾等各种害虫。
这两个桔红心大白菜新品种均为春、秋兼用品种,由于品质、商品性好,适合包装箱菜、超市销售,适宜在山东及全国其它相似生态地区种植。
3讨论与结论
桔红心大白菜因其叶球内叶颜色鲜艳呈现桔红色而得名。根据以往的文献,桔红心大白菜有两个来源:一是引种于日本[13];二是通过生物技术手段将大白菜与桔红色的芜菁杂交而得到的一个新的种质资源[5]。日本最早出现的桔红心大白菜也是日本白菜遗传育种学家在利用有性杂交将芜菁的耐抽薹性等性状转入大白菜的选育过程中获得的。因此,桔红心大白菜中控制桔红心形成的关键基因来自于桔红色的芜菁。目前,关于桔红心大白菜叶色成分及遗传育种方面的研究主要有如下进展:(1)选育并推广了一批桔红心大白菜新品种,丰富了白菜市场,初步满足了消费者对多彩蔬菜的需求[13-16];(2)在大白菜中,桔红心属隐性性状,由单个基因控制;(3)参照拟南芥、番茄等模式作物,逐步确定了桔红心大白菜叶球中类胡萝卜素含量高,桔红色是由于球内叶中产生了过多的前番茄红素等类胡萝卜素而形成的[5,17-19];(4)仅在2012—2015年的3年时间里,许多研究小组就有关桔红心连锁遗传图谱、利用图位克隆手段克隆桔红心基因的研究结果进行了各自独立的报道[17-24]。尽管这些研究手段及结果有交叉和类同之处,但均是独立完成的。控制大白菜桔红心形成的关键候选基因编码类胡萝卜素异构酶(CRTISO),基因编号为Bra031539,位于A09号染色体末端上。2017年,我们课题组报道了桔红心球叶发育的转录组表达谱分析结果,基于BrCRTISO基因,开发了一个新的InDel分子标记和一个新的SNP标记[12]。这两个标记已应用于育种实践,并获得了很好的育种效果。尽管以往研究取得了长足进展,对桔红心形成的分子机制及其应用了解的越来越清晰,但是有一个问题始终未解决:BrCRTISO是控制大白菜桔红心形成的关键候选基因,尚未有足够的证据彻底将“候选”二字去掉。目前,我国白菜基因组学研究处于国际领先地位,相信这个问题将会很快被解决。
大白菜属于十字花科芸薹属。十字花科包含模式植物拟南芥以及诸多具有遗传、形态多樣性的物种。近年来,我国科学家在十字花科植物,特别是芸薹属植物的基因组学研究方面做出了突出贡献,积累了丰富的基因组测序数据[25-29]。2000年,国际拟南芥基因组合作联盟完成了拟南芥全基因组测序与分析。随后拟南芥功能基因组学研究快速发展,为各国科学家研究植物基因功能以及农作物遗传改良提供大量便利的信息和研究方法。如何利用拟南芥的相关研究成果服务于大白菜研究,是一个很有意义的问题。比较基因组学的发展为我们提供了很好的思路和方法。两种具有较近共同祖先的植物,它们具有种属差异的基因组是由祖先基因组进化而来的,两种生物在进化的阶段上越接近,它们的基因组相关性就越高。如果生物之间存在很近的亲缘关系,那么它们的基因组就会表现出共线性,即基因序列部分或全部保守。这样就可以利用基因组编码序列、结构上的同源性,通过已知基因组的作图信息定位另外基因组中的基因,从而揭示基因的潜在功能以及基因组的内在结构[29]。桔红心大白菜叶球内叶黄色、桔红色是由于类胡萝卜素的积累造成的,其合成途经及分子调控网络与拟南芥很相似。大白菜桔红心候选基因BrCRTISO与拟南芥、番茄CRTISO有类似的结构和功能。根据拟南芥、番茄CRTISO的结构和功能信息,可以获得控制大白菜桔红心的基因。我们的研究工作证明了这一点。通过与拟南芥基因的共线性分析,挖掘和克隆与大白菜重要农艺性状相关的关键候选基因,是开展大白菜分子育种的快速高效的途径之一。
利用分子标记、形态标记进行多个优良基因的聚合育种,是创新种质、缩短育种进程的有效方法。我们在利用分子标记辅助创制优异桔红心大白菜种质的过程中,注意对早熟、叶球中小型等性状的选育。随着生活水平的提高及反季节蔬菜的快速发展,中小型叶球、早熟以及口味好、外观品质优良等性状越来越成为今后大白菜育种的目标性状。在育种实践中,我们综合考虑上述因素,最终将新品种选育由原来的5~10年缩短到3年半,选育的新品种抗病高产,同时好看好吃。
参考文献:
[1]柯桂兰. 中国大白菜育种学[M]. 北京:中国农业出版社,2010:16-34.
[2]徐家炳,张凤兰. 中国大白菜图鉴[M]. 北京:中国农业出版社,2016:1-32.
[3]丁谦,杜立忠,陈士亮,等. 橘红心大白菜遗传育种研究进展[J]. 山东农业科学,2014,46(12):129-132.
[4]钱前,郭龙彪,曾大力. 水稻分子育种技术指南[M]. 北京:科学出版社,2012:417-471.
[5]李佩荣,张淑江,章时蕃,等. 大白菜橘红心类胡萝卜素组分及其基因分析[J]. 园艺学报,2014,41(3):469-478.
[6]Park H,Kreunen S S,Cuttriss A J,et al. Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis,prolamellar body formation,and photomorphogenesis[J]. The Plant Cell Online,2002,14(2):321-332.
[7]Isaacson T,Ronen G,Zamir D,et al. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of β-carotene and xanthophylls in plants[J]. The Plant Cell Online,2002,14(2):333-342.
[8]Galpaz N,Ronen G,Khalfa Z,et al. A chromoplast-specific carotenoid biosynthesis pathway is revealed by cloning of the tomato white-flower locus[J]. Plant Cell,2006,18: 1947-1960.
[9]Breitenbach J,Vioque A,Sandmann G. Gene sll0033 from Synechocystis 6803 encodes a carotene isomerase involved in the biosynthesis of all-E lycopene [J]. Z. Naturforschung C,2001,56(9/10):915-917.
[10]Masamoto K,Wada H,Kaneko T,et al. Identification of a gene required for cis-to-trans carotene isomerization in carotenogenesis of the cyanobacterium Synechocystis sp. PCC 6803[J]. Plant Cell Physiol.,2001,42(12):1398-1402.
[11]Wang X,Wang H,Wang J,et al. The genome of the mesopolyploid crop species Brassica rapa[J]. Nature Genetics,2011,43:1035-1039.
[12]Li J J,Zhang Y H,Ding Q,et al. Transcriptome analysis of orange head Chinese cabbage (Brassica rapa L. ssp. pekinensis) and molecular marker development[J]. International Journal of Genomics,2017:6835810.
[13]徐家炳,陈广,张凤兰,等. 一种桔红心白菜的选育方法: 99103404[P].2002.
[14]杨晓云. 高端白菜桔红心大白菜M168[J]. 蔬菜,2009(12):13.
[15]苏学军,徐茂俊. 彩色大白菜新品种红抗1、2号的选育[J]. 中国瓜菜,2005(4):25-27.
[16]王翠花,何启伟,牟晋华. 橘红心大白菜系列新品种[J].长江蔬菜,2009(7):9.
[17]Seohee L, Sang-Choon L, Dong H B, et al. Association of molecular markers derived from the BrCRISTO1 gene with prolycopeneen riched orange-colored leaves in Brassica rapa[J]. Theoretical & Applied Genetics,2014,127(1):179-191.
[18]Su T B, Yu S C, Wang J,et al. Loss of function of the carotenoid isomerase gene BrCRTISO confers orange color to the inner leaves of Chinese cabbage (Brassica rapa L. ssp. pekinensis)[J]. Plant Mol. Biol. Rep.,2015,33:648.
[19]Li P R, Zhang S J, Zhang S F,et al. Carotenoid identification and molecular analysis of carotenoid isomerase-encoding BrCRTISO,the candidate gene for inner leaf orange coloration in Chinese cabbage[J]. Mol. Breeding,2015,35:72.
[20]Zhang J,Yuan H,Fei Z,et al. Molecular characterization and transcriptome analysis of orange head Chinese cabbage (Brassica rapa L. ssp. pekinensis)[J]. Planta,2015,241(6):1381-1394.
[21]Zhang J X,Li H X, Zhang M K,et al. Fine mapping and identification of candidate Br-or gene controlling orange head of Chinese cabbage (Brassica rapa L.ssp. pekinensis)[J]. Mol Breeding,2013,32(4):799-805.
[22]Zhang F L,Wang G, Wang M, et al. Identification of SCAR markers linked to or, a gene inducing beta-carotene accumulation in Chinese cabbage[J]. Euphytica,2008,164(2):463-471
[23]Zhang D S,Wang W H. Genetic relationship between Chinese cabbage with orange color in inner head and purple color in leaf[J]. China Vegetables,2011,18:25-29.
[24]Feng H,Li Y F,Liu Z Y,et al. Mapping of or, a gene conferring orange color on the inner leaf of the Chinese cabbage (Brassica rapa L. ssp. pekinensis)[J]. Molecular Breeding,2012,29(1):235-244.
[25]Yang J H,Liu D Y,Wang X W,et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection[J]. Nature Genetics,2016,48:1225-1232.
[26]Cheng F,Sun R F,Hou X L,et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea[J]. Nature Genetics,2016,48:1218-1224.
[27]Liu S, Liu Y,Yang X, et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes[J]. Nat. Commun.,2014,5(5):3930.
[28]Margaret R W,Cheng F,Piresc J C,et al. Origin, inheritance, and gene regulatory consequences of genome dominance in polyploids[J]. Plant Cell,2013,25:1541-1554.
[29]王曉波. 十字花科共线性分析及数据库平台的构建[D]. 北京:中国农业科学院,2015.
猜你喜欢
饮食与健康·下旬刊(2019年3期)2019-03-08
农村百事通(2018年23期)2018-02-21
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
天津农业科学(2016年12期)2017-01-11
阅读与作文(小学高年级版)(2016年3期)2016-03-08
天津农业科学(2015年5期)2015-05-30
